










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.1 San José Mar. 2013
Bioacústica del canto de advertencia de Ceratophrys cranwelli (Anura: Ceratophrydae)
Bioacoustic of the advertisement call of Ceratophrys cranwelli (Anura: Ceratophrydae)
Bioacoustic of the advertisement call of Ceratophrys cranwelli (Anura: Ceratophrydae)
*Dirrección para correspondencia:
Abstract
The advertisement call plays an important role in the life history of anuran amphibians, mainly during the breeding season. Call features represent an important character to discriminate species, and sound emissions are very effective to assure or reinforce genetic incompatibility, especially in the case of sibling species. Since frogs are ectotherms, acoustic properties of their calls will vary with temperature. In this study, we described the advertisement call of C. cranwelli, quantifying the temperature effect on its components. The acoustic emissions were recorded during 2007 using a DAT record Sony™ TCD-100 with stereo microphone ECM-MS907 Sony™ and tape TDK™ DAT-RGX 60. As males emit their calls floating in temporary ponds, water temperatures were registered after recording the advertisement calls with a digital thermometer TES 1 300±0.1°C. Altogether, 54 calls from 18 males were analyzed. The temporal variables of each advertisement call were measured using oscillograms and sonograms and the analyses of dominant frequency were performed using a spectrogram. Multiple correlation analysis was used to identify the temperature-dependent acoustic variables and the temperature effect on these variables was quantified using linear regression models. The advertisement call of C. cranwelli consists of a single pulse group. Call duration, Pulse duration and Pulse interval decreased with the temperature, whereas the Pulse rate increased with temperature. The temperature-dependent variables were standardized at 25°C according to the linear regression model obtained. The acoustic variables that were correlated with the temperature are the variables which emissions depend on laryngeal muscles and the temperature constraints the contractile properties of muscles. Our results indicated that temperature explains an important fraction of the variability in some acoustic variables (79% in the Pulse rate), and demonstrated the importance of considering the effect of temperature in acoustic components. The results suggest that acoustic variables show geographic variation to compare data with previous works.
Key words: Ceratophrys cranwelli, bioacoustics, temperature effects, ethology.
Resumen
El canto de advertencia en anuros es especie-específico, pero como los anfibios son organismos ectotermos, las variables acústicas pueden estar afectadas por la temperatura. Se describe el canto de advertencia de Ceratophrys cranwelli cuantificando el efecto de la temperatura sobre sus componentes. Los cantos se grabaron in situ y se registró la temperatura del sitio de canto. Un total de 54 cantos de 18 individuos fueron digitalizados y analizados. Las variables temporales de cada canto de advertencia fueron medidas a partir de los oscilogramas y sonogramas, mientras que el análisis de frecuencia se realizó a través del espectrograma. Mediante análisis de correlación múltiple se identificaron las variables acústicas temperatura-dependientes y el efecto de la temperatura fue cuantificado mediante los modelos de regresión lineal obtenidos. C. cranwelli posee un canto de advertencia simple pulsado. Las variables Duración del canto, Duración del pulso e Intervalo entre pulso decrecen con la temperatura, en tanto que la Tasa de pulsos está directamente correlacionada con la temperatura. Las variables temperatura-dependientes fueron estandarizadas a 25ºC según el modelo de regresión lineal obtenido para ser comparadas con datos publicados. Se demuestra la importante variabilidad que produce la temperatura en algunas propiedades del canto de C. cranwelli.
Palabras clave: Ceratophrys cranwelli, bioacústica, efecto de temperatura, etología.
Las señales sonoras constituyen un importante mecanismo de comunicación en los anfibios anuros, tanto a nivel intra como interespecífico (Duellman & Trueb 1986), cumpliendo una función estructuradora en las comunidades (Barrio 1964). El canto de advertencia actúa en la orientación de ambos sexos hacia los sitios de reproducción, en la formación de agregaciones reproductivas, en la localización y discriminación por parte de las hembras de los machos coespecíficos. Por lo tanto, estas emisiones sonoras son especie-específicas (Gerhardt & Davis 1988) y son muy importantes en sus historias de vida (Schneider & Sinsch 1992, Schneider et al. 1993, Márquez & Bosch 1995, Schneider & Sinsch 1999, Martino & Sinsch 2002). Debido a la condición ectotérmica de los anuros, la actividad acústica y las variables relacionadas con el canto están potencialmente influenciadas por la temperatura (Duellman & Trueb 1986). Se ha demostrado la correlación de la temperatura con variables temporales del canto de advertencia en diferentes especies de anfibios anuros (Blair 1958, Zweifel 1959, Gerhardt 1978, Gayou 1984, Schneider & Sinsch 1992, Schneider et al. 1993, Márquez & Bosch 1995, Navas 1996, Schneider & Sinsch 1999, Esteban et al. 2002, Martino & Sinsch 2002, Heyer & Reid 2003, Sinsch & Juraske 2006).
Barrio (1980) distingue las poblaciones diploides del género Ceratophrys como C. cranwelli, especie de amplia distribución en el Chaco argentino y el Gran Chaco paraguayo, mientras que las poblaciones octoploides de C. ornata (Bell 1843) se distribuyen en la región pampeana de Argentina. Barrio (1980) aporta datos bioacústicos de ambas especies de Ceratophrys y no obtuvo diferencias entre los cantos de advertencia de estas dos entidades, pero en el análisis no se consideró el efecto de la temperatura sobre sus componentes.
En el presente trabajo se describe el canto de advertencia de C. cranwelli cuantificando el efecto de la temperatura sobre las diferentes variables acústicas.
Materiales y métodos
Área de estudio: El trabajo de campo se realizó al Noreste de la localidad de Alejandro Roca (33º22’ S - 63º40’ W, altitud 210m.s.n.m.), departamento Juárez Celman, Provincia de Córdoba, Argentina. El predio corresponde a una Reserva “Recreativa” Natural y tiene dependencia municipal. Cuenta con 42 hectáreas, 16 de ellas están representadas por un cuerpo de agua permanente y otras ocho son inundables durante las estaciones de lluvias. Debido a la escasa pendiente y a las depresiones presentes en el lugar se produce la acumulación de agua en la época con la consecuente formación de charcas temporarias. El clima de la región es templado-sub-húmedo, con un régimen de precipitaciones medios anuales de 900mm, concentrados en el periodo primavera-verano (Valetti 2003).
Descripción de la especie: Es un anuro de la familia Ceratophryidae de tamaño mediano (70-130mm). Se lo conoce vulgarmente con el nombre de “escuerzo grande” y por su aspecto y tamaño es temido por los lugareños. Su cabeza es grande y chata de contorno casi circular. Sus ojos son prominentes y dorsales con pupilas lenticulares. Una característica distintiva respecto de C. ornata son sus crestas triangulares en el borde superior de sus párpados. Es de color marrón claro con manchas alargadas oscuras dispuestas simétricamente en el dorso, y el vientre es blanco amarillento. Sus extremidades poseen bandas oscuras transversales. En los machos se observa el saco vocal de color negro (Barrio 1980, Cei 1980, Gallardo 1987). Di Tada (1999) expresa que esta especie se reproduce en cuerpos de agua temporarios y que posee ciclos de reproducción trianuales. Su distribución geográfica se relaciona con el Chaco de Argentina, Bolivia, Paraguay y sudeste de Brasil. En el extremo sudeste de su distribución se encuentra en simpatría con la especie críptica C. ornata (Cei 1980, Di Tada 1999).
Trabajo de campo: El día 15 de Diciembre de 2007, luego de una intensa lluvia de 90mm, los machos de C. cranwelli comenzaron su actividad acústica a las 21:30h y permanecieron activos hasta las 05:00h del día posterior en una laguna temporaria de aproximadamente 900m2. Los individuos vocalizaban semi-sumergidos en el interior de la laguna de agua clara de 5 a 30cm de profundidad. El registro de las series de cantos pertenecientes a C. cranwelli se realizó desde las 22:00hr hasta las 03:00hr mediante un grabador Sony™ TCD-100 con micrófono estéreo ECM-MS907 Sony™ y cintas de registro TDK™ DA-RXG 60. Debido a que los machos vocalizaban a una distancia mayor a dos metros entre ellos, los registros acústicos se realizaron de a uno por vez, y como el lugar de canto era en el interior de la laguna, la temperatura del agua en el sitio de canto de cada individuo fue registrada inmediatamente después de la grabación acústica mediante un termómetro digital TES 1 300±0.1°C.
Trabajo de laboratorio: Los registros acústicos se procesaron mediante el uso del programa Canary 1.2 (Charif et al. 1995). Los cantos se digitalizaron a 44 100Hz y 16bits y fueron medidos mediante el uso de oscilogramas, sonogramas, y espectrogramas según Valetti et al. (2009). Los parámetros acústicos medidos fueron: (1) Duración del canto [ms], (2) Intervalo entre cantos [ms], (3) Número de pulsos por canto, (4) Duración del pulso [ms], (5) Intervalo entre pulsos [ms], (6) Tasa de pulsos [pulsos/s], (7) Frecuencia dominante [Hz]. Se analizaron tres cantos de 18 individuos (54 cantos en total) y para los análisis posteriores se calculó la media aritmética de cada variable acústica por individuo.
Mediante un análisis de correlación múltiple se identificaron las variables acústicas que dependen de la temperatura, de aquí en adelante temperatura-dependientes (p<0.05) y posteriormente se cuantificó el efecto de la temperatura sobre estas variables mediante análisis de regresión lineal. Las variables temperatura-dependientes fueron estandarizadas a 25ºC utilizando los modelos de regresión lineal obtenidos (Heyer & Reid, 2003) para poder ser comparadas con las del trabajo de Barrio (1980). Todos los análisis estadísticos fueron realizados con el programa Statgraphics Plus 5.0. Los registros acústicos digitalizados se depositaron en la sonoteca particular del Dr. Adolfo Martino en la cátedra de Ecología General, Departamento de Ciencias Naturales, Universidad Nacional de Río Cuarto, Argentina.
Resultados
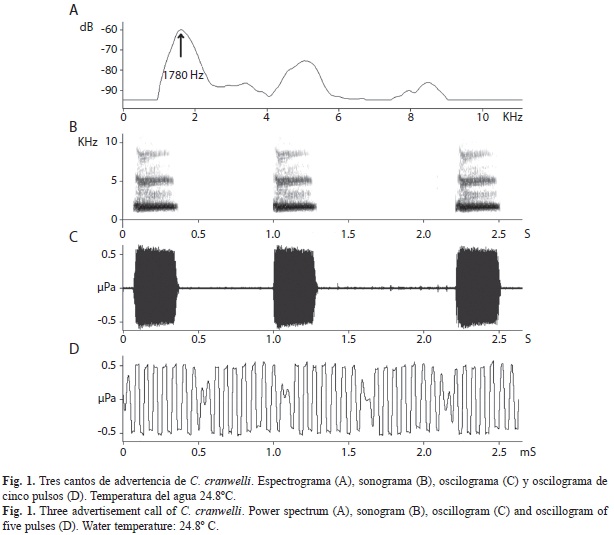
El canto de advertencia de C. cranwelli está constituidos por un número variable de pulsos (Fig. 1). La intensidad del canto varía, es elevada en los pulsos del centro y baja en los pulsos de los extremos del canto. De esta manera, al observarse en el oscilograma (Fig. 1), el canto muestra una forma de “huso”. La Duración de los pulsos y los Intervalos entre pulsos no varían a lo largo del canto. El análisis del sonograma muestra que la Frecuencia fundamental corresponde a la Frecuencia dominante. Esta frecuencia tiene un valor promedio de 1 715Hz, y es seguida por un número variable de armónicos de mayor frecuencia y menor intensidad.
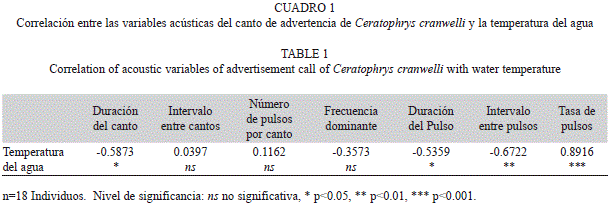
El análisis de correlación múltiple muestra que cuatro variables son temperatura-dependientes (Cuadro 1). La Duración del canto, la Duración del pulso y el Intervalo entre pulsos disminuyen al aumentar la variable Temperatura, mientras que la Tasa de pulsos aumenta con el incremento de la Temperatura (Cuadro 2). En la figura 2 se pueden observar las regresiones lineales de las variables temperatura-dependientes. La temperatura explica la variabilidad en las componentes acústicas temperatura-dependientes desde el 28% en la Duración del pulso hasta el 79% en la variable Tasa de pulsos (Cuadro 2).
En el cuadro 3 se muestran los valores medios y desviaciones estándar de las variables acústicas del canto de advertencia de C. cranwelli. Las variables temperatura-dependientes fueron previamente estandarizadas a 25ºC de temperatura según el modelo de regresión lineal obtenido.
La Duración del canto de advertencia es de 317ms a 25ºC disminuyendo 14.2ms por cada grado centígrado de temperatura que se incrementa (Cuadro 2). El Número de pulsos por cantos es de 61 y es independiente de la temperatura. La disminución tanto de la Duración del pulso como el Intervalo entre pulsos, respecto al incremento de la Temperatura, producen una disminución de la Duración del canto, sin afectar de manera significativa el Número de pulsos por canto.
Discusión
Las propiedades de los cantos de advertencia en los anfibios anuros están controladas principalmente por las contracciones musculares del tronco y de la laringe, induciendo el pasaje de aire desde los pulmones hacia los sacos vocales y atravesando el aparato laríngeo que es el lugar donde se produce el sonido (Duellman & Trueb 1986). En este sentido, en estudios previos se ha demostrado que la temperatura condiciona las propiedades contráctiles de los músculos del tronco y de la laringe involucrados en la producción del canto (Martin 1972, Manz 1975, Pough et al. 1992). Por lo tanto, como estos organismos son ectotermos, las propiedades acústicas de sus cantos son afectadas por la temperatura en la cual se producen dichas emisiones. Las variables acústicas que se correlacionaron con la temperatura del agua son aquellas variables cuyas emisiones dependen de los músculos del tronco y músculos laríngeos (Schmidt 1965, Martin & Gans 1972, Schneider 1977, Schneider 1988, Pough et al. 1992). En los hílidos, los cuales tienen cantos constituidos de una serie de pulsos repetidos, los músculos laríngeos abren y cierran la laringe en sincronía con los músculos del tronco para producir los pulsos regularmente espaciados que resultan importantes en el reconocimiento específico (Gerhardt 1991, Girgenrath & Marsh 1997). En general, la temperatura podría estar encubriendo diferencias o similitudes en propiedades acústicas entre especies cuando sus cantos de advertencias están siendo comparados a diferentes magnitudes de la misma.
La familia Ceratophryidae posee tres géneros, Chacophrys, Lepidobatrachus y Ceratophrys (Pyron & Wiens 2011). El canto de advertencia de C. cranwelli responde a la estructura temporal de canto simple (“single periodic pulse train”, Littlejohn 2001), al igual que los cantos de todas las especies de la familia Ceratophryidae descritos (Barrio 1963, 1968, 1980, Lescano 2011). En el trabajo de Barrio (1980) los valores de las variables acústicas están expresados para una temperatura de 25ºC en la localidad de Bell Ville. La Duración del canto de los individuos estudiados en dicha localidad tuvo un valor promedio de 350ms con cantos desde 320 a 400m. En este trabajo la Duración media del canto de advertencia fue de 317ms, con valores entre 272 y 348ms. La cantidad de pulsos por canto varió entre 53-70 con una media de 61, en tanto que para la localidad de Bell Ville este número fue mayor (70 pulsos promedio con valores entre 63 y 80). La Tasa de pulsos por segundos fue menor en la población estudiada comparada con la de Bell Ville (191.8, con valores entre 171-200, y 198, con valores entre 190-210, respectivamente). La Frecuencia dominante también resulta algo diferente entre las dos poblaciones. La media fue de 1 715Hz, con valores en un ámbito de 1 593-2 076, en tanto que Barrio (1980) reporta valores en un ámbito de 1 250-1 500 Hz.
Si bien los cantos de advertencia en los anuros son especie-específicos (Gerhardt & Davis 1988), determinadas propiedades del canto pueden variar con la temperatura, como se ha demostrado en este trabajo, y con la presencia de diferentes condiciones ecológicas en los ambientes (Gerhardt 1991). La variación geográfica en las propiedades acústicas de los anuros puede ser debida a distinta presiones selectivas (Castellano et al. 2003), esto sumado a los procesos estocásticos como la deriva génica (Smith & Hunter 2005). La variación geográfica del canto ha sido demostrada en numerosas especies de anuros. Particularmente en el área central de Argentina, Bionda et al. (2006) y Baraquet et al. (2007) encontraron variaciones geográficas en el canto de advertencia de Physalaemus biligonigerus y Hypsiboas pulchellus, respectivamente. Barrio (1980) encontró diferencias acústicas entre dos poblaciones distantes en aproximadamente 400km (Sumampa, Provincia de Santiago del Estero y Bell Ville, Provincia de Córdoba). Dado que la localidad del presente trabajo se encuentra a 125 y 440km respecto de las poblaciones Bell Ville y Sumampa, las diferencias respecto al trabajo de Barrio (1980) podrían deberse a variaciones geográficas.
Los resultados demuestran la importancia de la temperatura en la variabilidad de ciertas propiedades del canto en C. cranwelli, siendo la Tasa de pulsos una de las variables más importantes en la discriminación de los machos conespecíficos por parte de las hembras cuando están en simpatría con otras especies relacionadas (Sinsch 1990, Martino & Sinsch 2002, Holloway et al. 2006). Por lo tanto, resulta importante realizar un análisis bioacústico entre el par de especies C. cranwelli/C. ornata en un amplio rango de temperaturas y distintas localidades para evaluar si existen diferencias en los cantos de advertencia de estas dos especies.
Agradecimientos
La secretaría de Medio ambiente y recursos naturales de la municipalidad de Alejandro Roca, Córdoba, permitió realizar los trabajos de campo en la reserva natural municipal “Juan Carlos Fano”. Nuestros estudios fueron autorizados por la Secretaría Ambiente de la provincia de Córdoba, Argentina. La Secretaría de Ciencia y Técnica (SECyT-UNRC) subsidió en parte esta investigación (PPI 18/ C350). Al valioso aporte de los cuatro revisores anónimos que permitieron mejorar la versión final del manuscrito.
Referencias
Baraquet, M., N. Salas & I. Di Tada. 2007. Variación geográfica en el canto de advertencia de Hypsiboas pulchellus (Anura, Hylidae) en Argentina. Rev. Esp. Herp. 21: 107-118. [ Links ]
Barrio, A. 1963. Consideraciones sobre comportamiento y “grito agresivo” propio de algunas especies de Ceratophrynidae (Anura). Physis 24: 143-148. [ Links ]
Barrio, A. 1964. Caracteres eto-ecológicos diferenciales entre Odontophrynus americanus (Dumeril et Bibron) y O. occidentales (Berg) (Anura, Leptodactylidae). Physis 24: 385-390. [ Links ]
Barrio, A. 1968. Revisión del género Lepidobatrachus Budgett (Anura, Ceratophrynidae). Physis 28: 95-166. [ Links ]
Barrio, A. 1980. Una nueva especie de Ceratophrys (Anura: Ceratophryidae) del Dominio Chaqueño. Physis 39: 21-30. [ Links ]
Bell, T. 1843. Reptiles. In C. Darwin. Zoology of the voyage of H.M.S. Beagle under the command of Capt. Fitzroy R.N. during the years 1832 to 1836. Smith, Elder & Co., Londres, Reino Unido. [ Links ]
Bionda, C., N. Salas & I. Di Tada. 2006. Variación bioacústica en poblaciones de Physalaemus biligonigerus (Anura: Leptodactylidae) en Córdoba, Argentina. Rev. Esp. Herp. 20: 95-104. [ Links ]
Blair, W.F. 1958. Mating call in the speciation of anuran amphibian. Am. Nat. 92: 27-51. [ Links ]
Castellano, S., C. Giacoma & M. Ryan. 2003. Call degradation in diploid and tetraploid green toads. Biol. J. Linn. Soc. 78: 11-26. [ Links ]
Cei, J.M. 1980. Amphibians of Argentina. Monit. Zool. Ital. Monogr. Nº 2, Torino. [ Links ]
Charif, R.A., S. Mitchell & C.W. Clarck. 1995. Canary 1.2 user’s manual. Cornell Laboratory of Ornithology, Ithaca, Nueva York, EE.UU. [ Links ]
Di Tada, E.I. 1999. Patrones de distribución de los Anfibios Anuros de la provincia de Córdoba. Tesis doctoral en Ciencias Biológicas, Universidad Nacional de Córdoba, Córdoba, Argentina. [ Links ]
Duellman, W.E. & L. Trueb. 1986. Biology of Amphibians. McGraw-Hill Book, Nueva York, EE.UU. [ Links ]
Esteban, M., M.J. Sánchez-Herráiz, L.J. Barbadillo, J. Castanet & R. Márquez. 2002. Effects of age, size and temperature on the advertisement calls of two Spanish populations of Pelodytes punctatus. Amphibia Reptilia 23: 249-258. [ Links ]
Gallardo, J.M. 1987. Anfibios Argentinos. Guía para su identificación. Biblioteca Mosaico, Lib. Agropecuaria, Buenos Aires, Argentina. [ Links ]
Gayou, D.C. 1984. Effects of temperature on the mating call of Hyla versicolor. Copeia 3: 733-738. [ Links ]
Gerhardt, H.C. & M.S. Davis. 1988. Variation in the coding of species identity in the advertisement calls of Litoria verreauxi (Anura: Hylidae). Evolution 42: 556-565. [ Links ]
Gerhardt, H.C. 1978. Temperature coupling in the vocal communication system of the gray tree frog, Hyla versicolor. Science 199: 992-994. [ Links ]
Gerhardt, H.C. 1991 Female mate choice in treefrogs: static and dynamic acoustic criteria. Anim. Behav. 42: 615-635. [ Links ]
Girgenrath, M. & R.L. Marsh. 1997. In vivo performance of trunk muscles in tree frogs during calling. J. Exp. Biol. 200: 3101-3108. [ Links ]
Heyer, W.R. & Y.R. Reid. 2003. Does advertisement call variation coincide with genetic variation in the genetically diverse frog taxon currently known as Leptodactylus fuscus (Amphibia: Leptodactylidae)? An. Acad. Bras. Ciências 75: 39-54. [ Links ]
Holloway, A.K., D.C. Cannatella, H.C. Gerhardt & D.M. Hillis. 2006. Polyploids with different origins and ancestors form a single sexual polyploid species. Am. Nat. 167: E88-E101. [ Links ]
Lescano, J. 2011. Description of the advertisement and distress call of Chacophrys pierottii and comments on the advertisement call of Lepidobatrachus llanensis (Anura: Ceratophryidae). J. Nat. Hist. 45: 2929-2938. [ Links ]
Littlejohn, M.J. 2001. Patterns of differentiation in temporal properties of acoustic signals of anurans, p.102-120. In M.J. Ryan (ed.). Anuran communication. Smithsonian Institution, Washington, EE.UU. [ Links ]
Manz, R. 1975. Die Fusionsfrequenzen der Kehlkopfmuskeln und eines Beinmuskels in Abhaengigkeit von der Temperatur bei europaeischen Froschlurchen (Anura). Zool. Jb. Physiol. 79: 221-245. [ Links ]
Márquez, R. & J. Bosch. 1995. Advertisement calls of the midwife toads Alytes (Amphibia, Anura, Discoglossidae) in continental Spain. J. Zool. Systemat. Evol. Res. 33: 185-192. [ Links ]
Martin, W.F. 1972. Evolution of vocalizations in the genus Bufo, p. 279-309. In W.F. Blair (ed.). Evolution in the genus Bufo. University of Texas, Austin, EE.UU. [ Links ]
Martin, W.F. & C. Gans. 1972. Muscular control of vocal tract during release signalling in the toad Bufo valliceps. J. Morphol. 137: 1-27. [ Links ]
Martino, A.L. & U. Sinsch. 2002. Speciation by polyploidy in Odontophrynus americanus. J. Zool. Lond. 257: 67-81. [ Links ]
Navas, C.A. 1996. The effect of temperature on the voval activityof tropical anurans: A comparison of high and low-elevation species. J. Herpetol. 30: 488-495. [ Links ]
Pough, F., W. Magnusson, M.J. Ryan, K. Wells & T. Taigen. 1992. Behavioral energetics, p. 395-436. In M.E. Feder & W.W. Burggren (eds.). Environmental Physiology of Amphibians, Chicago University, Chicago, Illinois, EE.UU. [ Links ]
Pyron, R.A. & J.J. Wiens. 2011. A large-scale phylogeny of Amphibia including over 2,800 species, and a revised classification of extant frogs, salamanders, and caecilians. Mol. Phylogenet. Evol. 61: 543-583. [ Links ]
Schmidt, R.S. 1965. Larynx control and call production in frogs. Copeia 1965: 143-147. [ Links ]
Schneider, H. 1977. Acoustic behaviour and physiology of vocalization in the European tree frog, Hyla arborea (L.). In D.H. Taylor & S.I. Guttman (eds.). The reproductive biology of amphibians, Nueva York, Plenum, EE.UU. [ Links ]
Schneider, H. 1988. Peripheral and central mechanisms of vocalization, p. 537-558. In B. Fritzsch, M.J. Ryan, W. Wilczynsky, T.E. Hetherington & W. Walkowiak (eds.). The evolution of the Amphibian Auditory System, Wiley, Nueva York, EE.UU. [ Links ]
Schneider, H. & U. Sinsch. 1992. Mating call variation in lake frogs referred to as Rana ridibunda Pallas, 1771: taxonomic implications, Zeitschrift für Zoologische Systematik und Evolutionsforschung 30: 297-315. [ Links ]
Schneider, H. & U. Sinsch. 1999. Taxonomic reassessment of Middle Eastern lake frogs: Bioacoustic variation among populations consideres as Rana ridibunda, R. bedriagae or R. levantina. J. Zool. Systemat. Evol. Res. 57: 57-65. [ Links ]
Schneider, H, U. Sinsch & S. Sofianidout. 1993. The water frogs of Greecebioacoustic evidence for a new species. Zeitschrift für Zoologische Systematik und Evolutionsforschung 31: 47-63. [ Links ]
Sinsch, U. 1990. Froschlurche (Anura) der zentralperuanischen Anden: Artdiagnose, Taxonomie, Habitate, Verhaltensökologie. Salamandra 26: 177-214. [ Links ]
Sinsch, U. & N. Juraske. 2006. Advertisement calls of hemiphractine marsupial frogs: I. Gastrotheca marsupiata group. Herpetologia Bonnensis II. Proceedings of the 13th Congress of the Societas Europaea Herpetologica 145-148. [ Links ]
Smith, M.J. & D. Hunter. 2005. Temporal and geographic variation in the advertisement call of the booroolong frog (Litoria booroolongensis: Anura: Hylidae). Ethology 111: 1103-1115. [ Links ]
Valetti, J.A. 2003. Monitoreo Acústico de una Comunidad de Anfibios en una Reserva Natural (Alejandro Roca, Córdoba, Argentina). Tesis de grado de Licenciatura en Ciencias Biológicas. Universidad Nacional de Río Cuarto, Río Cuarto, Argentina. [ Links ]
Valetti, J.A., N.E. Salas & A.L. Martino. 2009. A new polyploid species of Pleurodema (Anura: Leiuperidae) from Sierra de Comechingones, Córdoba, Argentina and redescription of Pleurodema kriegi (Müller, 1926). Zootaxa 2073: 1-21. [ Links ]
Zweifel, R.G. 1959. Effect of temperature on call of the frog Bombina variegata. Copeia 1959: 322-327. [ Links ]
*Correspondencia:
Julián Alonso Valetti. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8-km 601, (X5804BYA) Río Cuarto, Argentina; jvaletti@exa.unrc.edu.ar.
Nancy Edith Salas . Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8-km 601, (X5804BYA) Río Cuarto, Argentina; nsalas@exa.unrc.edu.ar.
Adolfo Ludovico Martino. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8-km 601, (X5804BYA) Río Cuarto, Argentina; amartino@exa.unrc.edu.ar.
1 Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8-km 601, (X5804BYA) Río Cuarto, Argentina; jvaletti@exa.unrc.edu.ar, nsalas@exa.unrc.edu.ar, amartino@exa.unrc.edu.ar
Recibido 10-I-2012. Corregido 18-VII-2012. Aceptado 20-VIII-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}