










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.1 San José Mar. 2013
Dinámica poblacional y reproductiva de Turbinella angulata y Busycon perversum (Mesogasteropoda: Turbinellidae y Melongenidae) en el Banco de Campeche, México
Turbinella angulata and Busycon perversum (Mesogasteropoda: Turbinellidae and Melongenidae) population and reproductive dynamics in the Campeche Bank, Mexico
Turbinella angulata and Busycon perversum (Mesogasteropoda: Turbinellidae and Melongenidae) population and reproductive dynamics in the Campeche Bank, Mexico
*Dirección para correspondencia:
Abstract
Turbinella angulata and Busycon perversum are two marine gastropods that support the 90% of conch fisheries in the Gulf of Mexico, nevertheless there is no information available to support a management strategy for both species. The aim of this work was to obtain preliminary fishery information to promote management actions for both species in the Campeche Bank. The population of conchs was sampled in 12 transects (150x1.5m) by SCUBA diving from April 2008 to March 2010. All conchs were collected and siphonal length (SL) was measured. These samples were complemented with data from the commercial catches, analyzing a total of 2 832 individuals of T. angulata. Siphonal length varied between 63 and 282mm. Sex ratio was of 0.98:1 female:male. The von Bertalanffy growth equation parameters were: L∞=346mm; K=0.30 and t0=-0.24. Spawning extended from January to May with a long resting stage from June to September for over 80% of the adult population. The first gonadic maturity (L50) was at 210mm (SL) for females and 200mm for males. For B. perversum a total of 1 655 individuals were analyzed. Siphonal length varied between 78 and 291mm with significant differences between male and female (p<0.05). Sex ratio was of 1:0.25 female:male. The growth parameters obtained were: L∞=360mm; K=0.35 and t0=-0.19. The spawning period was from August to December with resting stage from January to April. The L50 was 215mm (LS) for females and 190mm for males. Both species presented one recruitment pulse. These results will be useful to design management schemes based on population dynamics and reproductive attributes.
Key words: growth, recruitment, reproductive cycle, first gonadic maturity, Campeche.
Resumen
Turbinella angulata y Busycon perversum aportan la mayor captura de caracol en el Golfo de México (90%).Se realizaron muestreos mediante transectos lineales (150x1.5m) de abril 2008 a marzo 2010 y en las capturas comerciales del Banco de Campeche (18°-20º N - 90°- 91º O). 2 832 individuos de T. angulata fueron medidos, las tallas variaron entre 63-282mm (LS), la proporción hembras:machos fue 0.98:1. Los valores de crecimiento fueron: L∞=346mm, K=0.30 y t0=-0.24. El periodo reproductivo fue de enero a mayo con reposo de junio a septiembre (80%). La talla de primera madurez (L50) fue de 210mm (LS) en hembras y 200mm en machos. De B. perversum,1 655 ejemplares fueron analizados con tallas entre 78-292mm, presentando diferencias significativas entre longitudes de machos y hembras (p<0.05). La proporción hembras:machos fue 1:0.25. Los valores de crecimiento fueron L∞=360 mm, K=0.35 y t=-0.19. El periodo reproductivo fue de agosto a diciembre con reposo de enero a abril. La L50 fue de 215mm (LS) en hembras y 190mm en machos. Se observó un pulso de reclutamiento anual en ambas especies relacionado con el incremento en la temperatura marina. Los resultados ayudarán a definir un esquema de manejo basado en los atributos poblacionales y reproductivos de las especies.
Palabras clave: crecimiento, reclutamiento, ciclo reproductivo, talla de primera madurez, Campeche.
La producción mundial de moluscos es de alrededor de siete millones de toneladas. En la última década, se han producido importantes cambios en sus pesquerías, mientras que las capturas de cefalópodos incrementaron más de un millón de toneladas, las capturas de moluscos de aguas continentales descendieron notablemente, debido a su extrema vulnerabilidad, a la degradación del hábitat y a la sobreexplotación (FAO 2010).
La producción de moluscos en México en 2010 fue de 177 737 toneladas, con valor total de 130 000 dólares americanos, lo cual corresponde al 11% del volumen y al 7.7% del valor de la producción pesquera nacional. Las pesquerías de moluscos de mayor importancia en el Golfo de México son la de Octopus maya y Octopus vulgaris, Crassostrea virginica, Turbinella angulata y Busycon perversum. La última incrementó considerablemente su volumen de captura de 1980 a 1992, pasando de 2 000 a 9 000 toneladas. En este periodo, el 94% de la captura correspondió al estado de Campeche y el 6% restante a los estados de Quintana Roo, Tabasco y Veracruz (CONAPESCA-SAGARPA 2010).
En la península de Yucatán, la captura de caracoles mutiespecífica, compuesta de
siete especies comerciales: Strombus gigas, Strombus costatus, Strombus pugilis, Turbinella angulata, Busycon perversum, Pleuroploca gigantea, Melongena melongena y Melongena corona bispinosa (De la Torre 1984, Baqueiro-Cárdenas et al. 2000, Pérez-Pérez & Aldana-Aranda 2000). En los últimos 10 años, las especies predominantes en la captura han sido T. angulata (50%) y B. perversum (46%), el
4% restante lo aportan S. costatus y P. gigantea (CONAPESCA-SAGARPA2010).
T. angulata, se distribuye desde Florida hasta la península de Yucatán y el Caribe. B. perversum, es una especie particular de la bahía de Campeche. Ambas especies presentan sexos separados, sin dimorfismo sexual exterior, a excepción del pene en los machos. Su fecundación es interna y el desarrollo embrionario es directo. La puesta de huevos en ambas especies tiene forma de cordón, con cápsulas ovígeras córneas en forma de moneda (Bandel 1976).
A pesar de la importancia económica y social que tiene el recurso caracol en el Golfo de México, no existen estudios que sirvan como base para un esquema de manejo que propicie la sustentabilidad de la pesquería. De las especies que componen la pesquería, las del género Strombus han sido las más estudiadas. En S. gigas, Appeldoorn (1988), Jesús Navarrete & Oliva-Rivera (1997), Jesús Navarrete et al. (2000) y Jesús Navarrete (2001), estimaron los parámetros de crecimiento, mortalidad y reclutamiento. D’Asaro (1965), Davis et al. (1987), Appeldoorn (1988), Corral & Ogawa (1987), Stoner et al. (1992) y Weill & Laughlin (1994), determinaron el periodo reproductivo de S. gigas en el Caribe y Golfo de México. Pérez-Pérez & Aldana-Aranda (2003), asociaron además la actividad reproductiva a los diferentes tipos de hábitat. Por otro lado, también se ha determinado el periodo reproductivo de S. pugilis, M. melongena y F. tulipa, observando diferentes estrategias gametogénicas como respuesta al ambiente: gametogénesis corta durante un periodo de tiempo o continúa durante la mayor parte del año (Reed 1995, Zetina-Zárate et al. 2000, Aldana-Aranda et al. 2003, Baqueiro-Cárdenas et al. 2005, Tapia-Arjona & Aldana-Aranda 2007).
Así, el objetivo del presente trabajo fue determinar los patrones reproductivos y poblacionales de las especies de caracol, dominantes en el Golfo de México: T. angulata y B. perversum, mismos que servirán como base para proponer recomendaciones de manejo.
Materiales y métodos
Área de estudio: El Banco de Campechese se localiza entre 18°20’ N - 90°91’ W, en la costa oeste de la península de Yucatán, es de origen kárstico, constituido por rocas calcáreas, con algunas playas formadas por sedimentos finos carbonatados de origen terrígeno y materia orgánica, producto del arrastre costero (Lankford 1974). En el área proliferan extensos bancos de Thalassia sp. y Syringodium sp. El clima es cálido-húmedo, con lluvias en verano y precipitación media anual de 1 132mm. La temperatura media anual es superior a 23°C, las mareas son de tipo mixto, con rangos máximos de 1.4m (García 1981).
Recolecta de muestras: La recolecta del material biológico se realizó de abril 2008 a marzo 2010, en Tenabo, Campeche y Champotón, sitios de mayor captura (Fig. 1). Se realizaron transectos lineales de 150x1.5m, recolectándose los organismos mediante buceo autónomo. Por otra parte, se realizaron muestreos en embarcaciones comerciales en los principales centros de descarga de cada localidad. A cada organismo se le tomó la longitud sifonal (LS), peso total (PT) y se determinó el sexo de forma macroscópica, el cual fue corroborado posteriormente con base en el análisis microscópico de las gónadas.
Análisis poblacional: Para establecer la estructura poblacional, se utilizaron los datos de longitud sifonal. Se estableció la proporción de sexos (hembra:macho). La relación peso-longitud se determinó usando la ecuación de Ricker (1975). Los parámetros de crecimiento del modelo de von Bertalanffy (1938) se estimaron con el programa ELEFAN 1.0 (Gayanilo et al.1989), utilizándose para tal fin las distribuciones de frecuencia mensual a intervalos de 10mm. La longevidad máxima
(Tmax), se calculó mediante el método de Taylor (1960). A partir de los parámetros de crecimiento, se estimó el patrón mensual de reclutamiento de la pesquería, usando el programa FISAT.
Análisis reproductivo: Para determinar el ciclo reproductivo de T. angulata y B. perversum, se recolectaron mensualmente 20 organismos adultos de cada especie (n=960). Para el proceso histológico fueron fijadas en Bouin e incluidas en Paraplast® secciones transversales de la parte media de la gónada-glándula digestiva (en ambas especies, la gónada está fusionada con la glándula digestiva). Se realizaron cortes histológicos de seis micras, teñidas posteriormente con Tricromo de Masson y Azul Alcian (Gabe 1968). Los cortes se analizaron a 10x y
40x en un microscopio óptico Primo StarZeiss. La presencia de células sexuales masculinas y femeninas así como la ausencia de estas, definieron a los individuos como machos, hembras o sexualmente indiferenciados.
A cada organismo se le asignó un estadio de desarrollo gonadal de acuerdo a las descripciones hechas por Lucas (1965) y Aldana-Aranda et al. (2003), las cuales consideran cinco etapas de acuerdo a lo siguiente:
• (Estadio I) Reposo: no hay diferenciación sexual, células germinales indiferenciadas.
• (Estadio II) Gametogénesis: presenta división celular activa, ovocitos y espermatocitos primarios y secundarios.
• (Estadio III) Madurez: con dominancia de gametos maduros, ovocitos con núcleo
y nucleolo bien definido, los espermatozoides presentan las colas dirigidas hacia el lumen.
• (Estadio IV) Evacuación o desove: los túbulos se encuentran parcialmente vacíos
con gametos remanentes.
(Estadio V) Post evacuación o post desove: los túbulos son parcial o totalmente vacíos con ovocitos y espermatozoides en reabsorción y presencia de fagocitos.
El período reproductivo en ambas especies, fue determinado por el desarrollode las células sexuales en la gónada. Para asignar las diferentes etapas de madurez sexual, en las hembras, se consideró la cantidad de tejido conjuntivo, de túbulos ovígeros, ovogonias y ovocitos. En los machos se consideró la cantidad de tejido conjuntivo, conductos espermáticos, espermatogonias, espermatocitos, espermátidas y espermatozoides (Aldana-Aranda et al. 2003). El ciclo reproductivo se estableció mediante las frecuencias mensuales de madurez gonádica.
Para determinar la talla de primera madurez para ambos sexos, se calculó el porcentaje de organismos maduros en intervalos de 10mm. Se ajustó una ojiva logística a los datos para determinar la talla a la que el 50% (L50) de los organismos se encontraron sexualmente maduros (Sparre &Venema 1997).
Para las dos especies (T. angulata y B. perversum), los análisis estadísticos incluyeron una comparación de la longitud sifonal y el peso total (LS y PT) entre sexos, mediante una prueba Z, considerándose una significancia estadística de p<0.05 (Zar 1996).
Se aplicó una prueba de χ2 para evaluar las desviaciones estadísticas significativas de una proporción sexual balanceada 1:1, entre hembras y machos de una misma especie, en las diferentes áreas de pesca, con un nivel de significancia de p<0.05 (Zar 1996). También se realizó un análisis de correlación entre el porcentaje de reclutamiento y la temperatura marina, usando el coeficiente de correlación de Spearman (Zar 1996).
Resultados
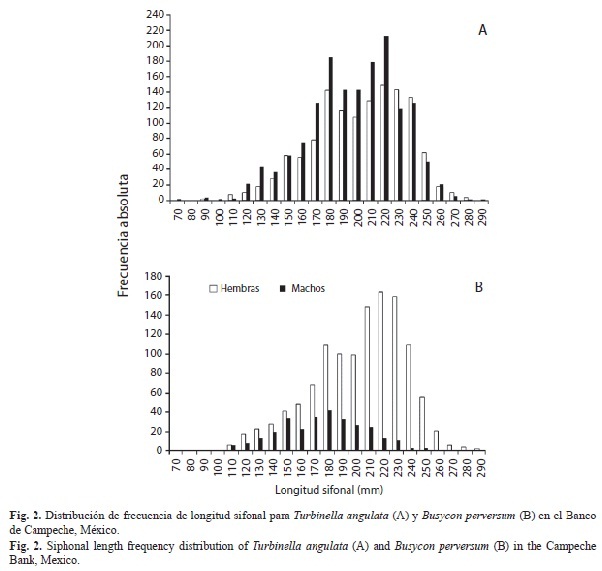
Estructura poblacional: Durante el periodo de estudio se analizaron 2 832 organismos de T. angulata (1 272 hembras, 1 560 machos). Se observaron tallas de longitud sifonal entre los 63-282mm y de peso total de 40-2 380g (Fig. 2A). No se encontró diferencia significativa entre las tallas de hembras y machos (χ2=7.43, p>0.05). En B. perversum, se analizaron un total de 1 655 organismos (1 326 hembras y 329 machos). Las hembras de sta especie fueron más grandes y pesadas, presentaron un intervalo de talla de 100-292mm y de peso total de 50-1 554g, mientras que los machos fueron menores (78-240mm, 28-1 130g) (Fig. 2B). Se observaron diferencias significativas entre el tamaño de los organismos y el sexo (χ2=25.91, p<0.05). El mayor porcentaje de machos se observó en las tallas menores (170-190mm LS), las hembras presentaron tallas de 210-250mm LS.
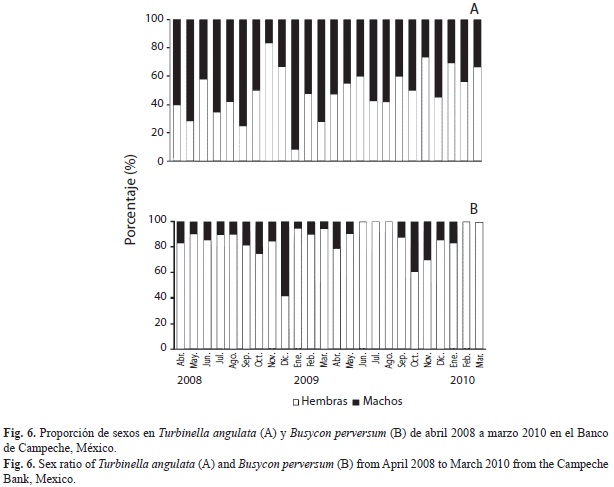
Proporción de sexo: La proporción hembra macho (H:M) en T. angulata, fue de 0.98:1, observándose la predominancia de machos durante el periodo de estudio, con excepción de noviembre 2008, noviembre 2009, enero y marzo 2010, en los que predominaron las hembras (Fig. 3A), no se observaron diferencias significativas entre los sitios de captura (χ2=3.24, p>0.05). Mientras que B. perversum presentó una relación H:M de 1:0.25, con dominancia de hembras durante el periodo de estudio (Fig. 3B). La proporción de hembras fue mayor que la de machos, se observó diferencia significativa en todas las zonas de pesca, 1:0.26 para Tenabo (χ2=25.33, p<0.05), 1:0.33 para Campeche (χ2=40.61, p<0.05) y 1:0.39 para Champotón (χ2=32.51, p<0.05).
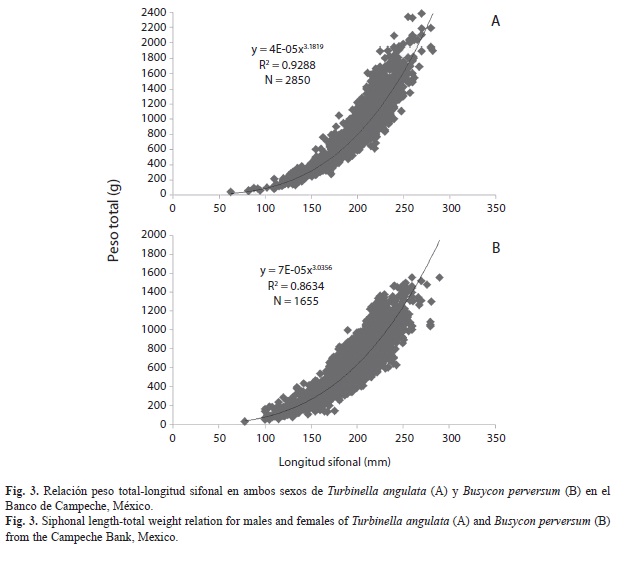
Relación peso-longitud: Las ecuaciones longitud-peso para T. angulata fueron:
PT=0.00005(LS)3.19, r2=0.92 en hembras; PT=0.00002(LS)3.27, r2=0.92 en machos y PT=0.00004(LS)3.18, r2=0.93 en ambos sexos (Fig. 4A). Para B. perversum las relaciones fueron: PT=0.00006 (LS)3.05, r2=0.86 en hembras; PT=0.00006(LS)3.06, r2=0.86 en machos y PT=0.00007(LS)3.03, r2=0.86 en ambos sexos (Fig. 4B). El valor de b observado en todos los casos no fue diferente de tres, confirmando que el crecimiento de ambas especies es isométrico.
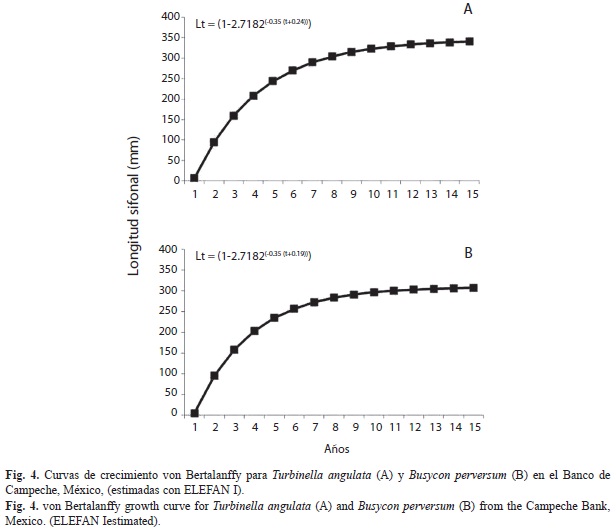
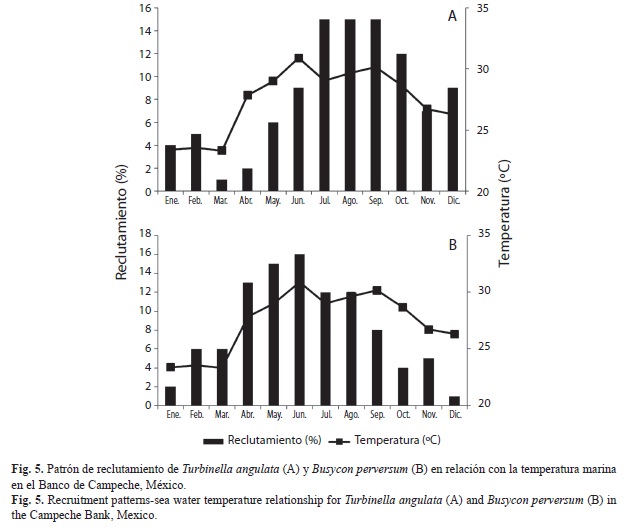
Crecimiento y reclutamiento: Los valores de crecimiento estimados para T. angulata fueron: L infinita (∞) de 346mm, tasa de crecimiento (K anual) de 0.30 y el valor de t0 fue de -0.24 (Fig. 5A). Los resultados obtenidos indican que la longevidad es de 12 años, la mayor parte de la población se encontró entre las edades de tres a seis años (60%). El crecimiento individual muestra un crecimiento muy leve a partir de los 320mm de LS, correspondiente a 10 años de edad.
Para B. perversum los valores de crecimiento estimados fueron: L∞=360mm, K anual de 0.35 y t0=-0.19 (Fig. 5B). La edad que se determino, varió entre dos y 15 años, y las edades mejor representadas se encontraron entre tres y siete años (56%). El crecimiento individual en esta población mostró un aumento muy leve a partir de 330mm de LS, el cual corresponde a 11 años de edad.
El reclutamiento de T. angulata, se observó a lo largo del año, con un periodo de mayor intensidad de mayo a diciembre (88%). En B. perversum, el patrón de ingreso de juveniles a la pesquería fue constante, con mayor intensidad de abril a septiembre (76%). En ambos casos, el mayor porcentaje de reclutamiento se relacionó con el incremento en temperatura del agua de mar. El coeficiente de correlación entre el porcentaje de reclutamiento y la temperatura marina fue de r2=0.70 para la primera especie (Fig. 6A) y r2=0.84 en la segunda.
Ciclo reproductivo: La figura 7A, muestra el ciclo reproductivo de T. angulata para ambos sexos. La gametogénesis se observó en tres periodos, de abril a junio 2008, de octubre 2008 a marzo 2009 con máximo en diciembre (67%) y de noviembre a marzo 2010 (40-54%). El período de madurez se observó de enero a mayo en los dos años con valores de 10-20%, la evacuación de gametos se presentó en tres pulsos: abril-mayo 2008 (50%), enero-mayo 2009 (75%), octubre-enero 2010 (35%) y la post emisión en dos pulsos: abril-junio (35%) y enero-septiembre 2009 (25%). Los periodos de reposo fueron prolongados de junio a septiembre (80%) en ambos años. En esta especie, la gametogénesis y madurez se relacionaron con las menores temperaturas en invierno (22-24°C); contrariamente, el reposo se observó cuando se presentaron temperaturas de 27 a 31°C, en primavera-verano. El coeficiente de correlación entre el porcentaje de organismos en reproducción (gametogénesis, madurez y desove) y la temperatura marina fue negativa, con valor de r2=-085.
El ciclo reproductivo de B. perversum se muestra en la figura 7B. En este se observó que el periodo de gametogénesis se registró en primavera, con presencia de organismos maduros a partir de abril. El pico de madurez fue de agosto (85%) a noviembre 2008 (60%) y de agosto (75%) a diciembre 2009 (60%). La evacuación de gametos se registró de septiembre a noviembre 2008 (20%) y de octubre a noviembre 2009 (20%). La mayor presencia de organismos en la post evacuación de gametos se registró en octubre (25%), febrero (20%) y de diciembre 2009 a marzo 2010 (30%), lo cual sugiere que la evacuación de gametos ocurre en otoño e invierno. Los caracoles (ambos sexos) permanecieron sexualmente inactivos de enero (50%) a abril 2009 (85%) y enero (30%) a marzo 2010 (83%). En esta especie los periodos de madurez y desove coincidieron con las mayores temperaturas y la etapa de reposo con el periodo de menor temperatura del agua. El coeficiente de correlación entre el porcentaje de organismos en reproducción (gametogénesis, madurez y desove) y la temperatura marina fue positiva con valor de r2=0.69.
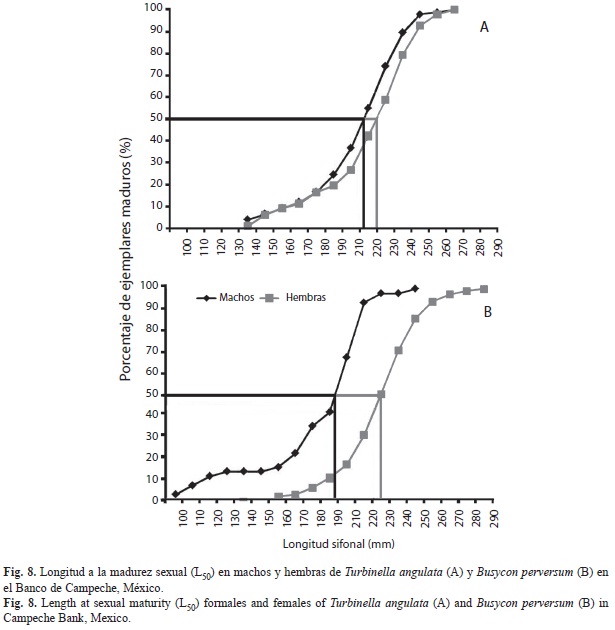
Talla de primera madurez: La figura 8A muestra la relación entre el porcentaje de T. angulata maduros y la longitud sifonal. La hembra madura más pequeña midió 102mm y la hembra madura más grande 260mm, estimándose L50 en 210mm para las hembras. El macho maduro más pequeño midió 132mm y el más grande 260mm, estimándose la L50 en 200mm para los machos. De acuerdo a las estimaciones realizadas, ambos sexos alcanzan la madurez sexual a los cuatro años de edad.
En la figura 8B, se muestra la relación entre el porcentaje de ejemplares maduros de ambos sexos y la longitud sifonal de B. perversum. La hembra madura más pequeña midió 160mm y la más grande, 280mm, por lo que la L50 se estimó en 215mm. El macho maduro más pequeño midió 98mm y el más grande 232mm, estimándose el valor de L50 en 190mm, cabe señalar que la estimación se realizó con 46 organismos. De acuerdo a estos valores, las hembras alcanzarían la talla de madurez sexual a la edad de cuatro años y medio, mientras que los machos a los cuatro años de edad.
Discusión
Los resultados permiten describir que la dinámica poblacional es particular para cada especie. T. angulata, constituye una población equilibrada sin diferencias significativas en las tallas, ni en la proporción de sexos. Contrariamente, B. perversum presentó diferencias significativas en las tallas y proporción de sexos, las hembras fueron mayores que los machos y predominaron en todos los sitios de pesca (80%). Estudios poblacionales de moluscos bivalvos realizados por Ansell (1961) y Wolowicz (1984), determinaron que la razón de sexos no varían con la edad, sin embargo, Derbali et al. (2009), describen un patrón similar al del presente estudio en una población de Cerastoderma glaucum, con presencia de machos entre los individuos de menor talla y predominio de hembras entre los de mayor talla. Así, el incremento de la proporción de hembras con la talla podría explicarse por un crecimiento diferencial entre sexos, donde los machos tienen un rol importante en la reproducción, pero viven poco y tienen pocas reservas, lo que causa que las hembras vivan entre cinco y 10 veces más, tiempo en el crecen y acumulan reservas para la reproducción (Derbali et al. 2009).
T. angulata y B. perversum presentan parámetros de crecimiento similares caracterizado por un crecimiento acelerado durante los primeros cuatro años (entre 80 y 40mm por año), mismo que va disminuyendo conforme avanza hacia la etapa adulta, entre el quinto y décimo año, los incrementos anuales variaron entre 30 y 10mm, pudiendo alcanzar longitudes máximas de 360 y 350mm de LS, a los 15 y 12 años de edad respectivamente. Baqueiro-Cárdenas & Aldana-Aranda (2003), en un estudio realizado con 13 especies de bivalvos y gasterópodos, observaron que a nivel multiespecífico, las especies que habitan en condiciones ambientales semejantes presentan características similares en tasa de crecimiento K y longitud máxima L∞ en función de la biología de cada especie.
Para ambas especies, no existen estudios de crecimiento que permitan realizar una comparación directa, aunque existen evaluaciones de crecimiento de B. carica, especie del mismo género, carnívora, morfológicamente similar a B. perversum y que habita en la misma región (Baqueiro-Cárdenas et al. 2000). Los valores reportados para B. carica son variables; Kraeuter et al. (1989) estima un crecimiento de 14.4mm por año durante los primeros 10 años y de 6.5mm por año entre los 12 y 14 años. Eversole et al. (2008), reporta una tasa media de crecimiento menor en B. carica desde <0.001 hasta 0.022mm/día, tomando el último valor, los ejemplares pueden alcanzar de 36 a 80mm en 10 años.
El reclutamiento de T. angulata y B. perversum presentó el mismo patrón: un ingreso constante de reclutas a través del año, con pulso anual de mayor magnitud. Sin embargo, se observaron variaciones en la temporalidad del reclutamiento entre ambas especies que coexisten en una misma localidad. Baqueiro-Cárdenas & Aldana-Aranda (2003) describen estas variaciones como factores de atenuación de competencia. El reclutamiento de ambas especies se correlacionó con la temperatura del agua. La temperatura tiene un papel fundamental en la dinámica de las poblaciones de moluscos bentónicos; de acuerdo con García-Cubas (1982), Kendall et al. (1987), Sanders (1988), Miller & Lawrence (1993), Delgado et al. (2004), Ramírez & Olabarría (2005), Aufderheide et al. (2006), Lima et al. (2006) y
Mieszkowska et al. (2007), las variaciones anómalas de temperatura pueden provocar cambios en los patrones de reclutamiento, en la estructura, distribución y abundancia poblacional.
De acuerdo a las características reproductivas, en ambas especies se observó un desarrollo gonádico simultáneo para hembras y machos, además ambas presentaron el patrón característico de los ciclos reproductivos de gasterópodos marinos, el cual inicia con la proliferación y desarrollo de gametos, seguido por una fase maduración sexual hasta la evacuación de gametos y culmina con una temporada de reposo (Aldana-Aranda et al. 2003, Baqueiro-Cárdenas et al. 2005).
Estudios realizados con gasterópodos de la región Golfo de México y Mar Caribe, se
menciona que los procesos de maduración y emisión de gametos pueden presentarse en uno o dos pulsos y también pueden ser constantes a través del año (Reed 1995, Zetina-Zárate et al. 2000, Aldana-Aranda et al. 2003, Baqueiro-Cárdenas et al. 2005). En T. angulata y B. perversum, los procesos de maduración y desove/evacuación fueron observados en un pulso reproductivo anual, sin embargo, la temporalidad de la etapa reproductiva fue diferente para cada especie, mientras que en T. angulata la gametogénesis y madurez se presentaron en primavera-verano; la población de B. perversum se encontraba en reposo. Así mismo, el periodo de maduración de B. perversum se presentó en invierno cuando la población de T. angulata se encontraba sexualmente inactiva. Un patrón reproductivo similar fue descrito por Aldana-Aranda et al. (2003), en los gasterópodos Melongena melongena y Fasciolaria tulipa, ambas especies con similar estrategia de vida, hábitos alimenticios y desarrollo larval directo, presentaron una clara alternancia en el pico de evacuación de gametos, mientras M. melongena lo tuvo de enero a abril, F. tulipa lo presentó en octubre y diciembre. Ambos estudios demuestran un control ambiental en el comportamiento del ciclo reproductivo y de estrategias adaptativas para optimizar la competencia entre especies. Lubet (1991) menciona que los factores que inducen la gametogénesis y definen la temporalidad e intensidad del ciclo reproductivo son la temperatura y disponibilidad de alimento.
La talla de primera madurez sexual ha sido determinada por primera vez para T. angulata y B. perversum en este trabajo. En la primera especie, la talla a la madurez sexual fue similar en machos y hembras, sin embargo en B. perversum la talla de hembras es superior en 30mm a la de los machos. En gasterópodos del Golfo de México, sólo existe una estimación de la talla de primera madurez de B. contrarium en 220mm (De la Torre 1984), este valor es similar al estimado en hembras de B. perversum (215mm). Los resultados obtenidos en este trabajo ayudarán a definir un esquema de manejo que propicie el aprovechamiento sustentable del recurso caracol en el Golfo de México.
Agradecimientos
Los autores expresan su agradecimiento a Thierry Brulé y Teresa Colás Marrufo (laboratorio de Ictiología del CINVESTAV-IPN Mérida) por la asesoría recibida y equipamiento. Al Centro Regional de Investigaciones Pesqueras de Lerma, Campeche especialmente a Isaac Rojas por las facilidades otorgadas para la realización de este estudio. A Manuel Medina y a José Francisco Chávez Villegas por el apoyo técnico. Al laboratorio de Biología Marina de la Universidad de las Antillas francesas (Olivier Gros) por su infraestructura y asesoría. Al Consejo Nacional de Ciencia y Tecnología por el apoyo con la beca para Doctorado en Ciencias No. 22342. Proyectos: Fondos Mixtos (CAMP-2008-01-96462) y ECOS-ANUIES (Caracterización reproductiva, molecular, ecológica y de interacción Apicomplexa-Strombidae en la región Caribe-Golfo de México).
Referencias
Aldana-Aranda, D., E. Baqueiro, I. Martínez-Morales, A. Zetina-Zárate & T. Brulé. 2003. A review of the reproductive patterns of gastropod mollusks from Mexico. Bull. Mar. Sci. 73: 629-641. [ Links ]
Ansell, A.D. 1961. Reproduction, growth and mortality of Venus striatula (da Costa) in Kames Bay, Millport. J. Mar. Biol. Assoc. U.K. 41: 191-215. [ Links ]
Appeldoorn, R.S. 1988. Age determination, growth, mortality and age of first reproduction in adult queen conch, Strombus gigas L., off Puerto Rico. Fish. Res. 6: 363-378. [ Links ]
Aufderheide, J., R. Warbritton, N. Pounds, S. File-Emperador, C. Staples, N. Caspers & V. Forbes. 2006. Effects of husbandry parameters on the life-history traits of the apple snail, Marisa cornuarietis: effects of temperature, photoperiod and population density. Invert. Biol. 125: 9-20. [ Links ]
Bandel, K. 1976. Egg masses of 27 Caribbean Opistobranchs from Santa Marta, Columbia. Stud. Neot. Fauna Environ. 11: 87-118. [ Links ]
Baqueiro-Cárdenas, E. & D. Aldana-Aranda. 2003. Patrones en la biología poblacional de moluscos de importancia comercial en México. Rev. Biol. Trop. 51: 97-107. [ Links ]
Baqueiro-Cárdenas, E., D. Aldana-Aranda & I. Martínez-Morales. 2005. Gonad development and reproductive pattern of fighting conch Strombus pugilis (Linee, 1758) (Gastropoda, Prosobranchia) from Campeche, Mexico. J. Shellfish Res. 24: 1127-1133. [ Links ]
Baqueiro-Cárdenas, E., D. Murillo & M. Medina. 2000. Fisheries Biology Topics of Conch Resources from the Northern Coast of Campeche, Mexico. Proc. Gulf. Carib. Fish. Inst. 51: 16-59. [ Links ]
CONAPESCA-SAGARPA. 2010. Carta Nacional Pesquera. Diario Oficial de la Federación. Mazatlán, Sinaloa, México (Consultado: 2 septiembre 2011, http://conapesca.sagarpa.gob.mx/wb/cona/actualizacion_de_la_carta_nacional_pesquera_2010). [ Links ]
Corral, J.L. & J. Ogawa. 1987. Cultivo masivo de larvas de caracol Strombus gigas en estanques de concreto. Proc. Gulf. Carib. Fish. Inst. 38: 345-351. [ Links ]
D’Asaro, C.N. 1965. Organogenesis, development and metamorphosis in the queen conch Strombus gigas, with notes on breeding habits. Bull. Mar. Sci. 15: 359-416. [ Links ]
Davis, M., C. Hesse & G. Hodgkins. 1987. Commercial hatchery produced queen conch, Strombus gigas, seed for the research and grow-out market. Proc. Gulf. Carib. Fish. Inst. 38: 326-335. [ Links ]
de Jesús Navarrete, A. & J.J. Oliva-Rivera. 1997. Densidad, crecimiento y reclutamiento del caracol rosado Strombus gigas en Punta Gavilán, Quintana Roo, México. Rev. Biol. Trop. 42: 797-801. [ Links ]
de Jesús Navarrete, A. 2001. Crecimiento del caracol Strombus gigas (Gastropoda: Strombidae) en cuatro ambientes de Quintana Roo, México. Rev. Biol. Trop. 49: 85-91. [ Links ]
de Jesús Navarrete, A., M. Domínguez-Viveros, A. Medina-Quej & J.J. Oliva-Rivera. 2000. Crecimiento, mortalidad y reclutamiento del caracol Strombus gigas en Punta Gavilán, Q. Roo, México. Ciencia Pesquera 14: 1-4. [ Links ]
De la Torre, R. 1984. Diagnóstico de la pesquería de caracoles en el estado de Quintana Roo. SEPESCA, México. [ Links ]
Delgado, G.A., C.T. Bartels, R.A. Glazer, N.J. Brown-Peterson & K.J. McCarthy. 2004. Translocation as a strategy to rehabilitate the queen conch (Strombus gigas) population in the Florida Keys. Fish. Bull. 102: 278-288. [ Links ]
Derbali, A., O. Jarboui & M. Ghorbel. 2009. Reproductive biology of the cockle Cerastoderma glaucum (Mollusca: Bivalvia) from the north coast of Sfax (Gulf of Gabes, Tunisia). Ciencias Marinas 35: 141-152. [ Links ]
Eversole, A.G., W.D. Anderson & J. Jeffery-Isely. 2008. Age and growth of Knobbed Whelk Busyconcarica (Gmelin, 1791) in South Carolina subtidal waters. J. Shellfish Res. 27: 423-426. [ Links ]
FAO, 2010. El estado mundial de la pesca y la acuicultura 2010. FAO, Roma, Italia. [ Links ]
Gabe, M. 1968. Techniques histologiques. Masson, Paris, France. [ Links ]
García, E. 1981. Modificaciones al sistema de clasificación climática de Köppen. Instituto de Geografía, UNAM, México D.F., México. [ Links ]
García-Cubas, A. 1982. Mollusks of a tropical lagoon system in the southern Gulf of Mexico (Laguna de Terminos, Campeche). An. Inst. Cienc. Mar Limnol. Univ. Nac. Auton. Mex. 294: 167-173. [ Links ]
Gayanilo, F.C., M. Soriano & D. Pauly. 1989. A draft guide to the complete ELEFAN. ICLARM Contribution 435: 65. [ Links ]
Kendall, M., P. Williamson & P. Garwood. 1987. Annual variations in recruitment and populations structure of Monodon talineata and Gibula umbilicalis populations at Aberaeron, Mid-Wales. Est. Coast. Shelf. Sci. 24: 499-511. [ Links ]
Kraeuter, J.N., M. Castagna & R. Bisker. 1989. Growth rate estimates for Busycon carica (Gmelin, 1791) in Virginia. J. Shellfish Res. 8: 219-225. [ Links ]
Lankford, R.R. 1974. Coastal lagoons of Mexico, their origin and classification, p. 182-215. In L.E. Cronnin (ed.). Estuarine processes. Circulation, sediments and transfer of material, in the Estuary. Academic, Nueva York, EEUU. [ Links ]
Lima, F.P., N. Queiroz, P.A. Ribeiro, S.J. Hawkins & A.M. Santos. 2006. Recent changes in the distribution of marine gastropod Patella rustica Linnaeus 1758, and their relationship to unusual climatic events. J. Biogeogr. 33: 812-822. [ Links ]
Lubet, P. 1991. Reproduction des mollusques, p. 167-203. In G. Barnabé (ed.). Bases Biologiques de l’Aquaculture. Tec et Doc, Lavoisier, Francia. [ Links ]
Lucas, A. 1965. Recherche sur la sexualité des mollusques bivalves. Tesis de Doctorado, Université de Rennes, France. [ Links ]
Mieszkowska, N., S.J. Hawkins, M.T. Burrows & M.A. Kendall. 2007. Long-term changes in the geographic distribution and population structure of Osilinus lineatus (Gastropoda: Trochidae) in Britain and Ireland. J. Mar. Biol. Assoc. U.K. 87: 537-545. [ Links ]
Miller, A.C. & S.E. Lawrence. 1993. Long-term trends in black abalone Haliotis cracherodii Leach, 1814, population along the Palos Verdes peninsula, California. J. Shellfish Res. 12: 195-200. [ Links ]
Pérez-Pérez, M. & D. Aldana-Aranda. 2000. Distribución, abundancia y morfometría de Strombus costatus, Turbinella angulata, Busycon contrarium y Pleuroploca gigantea (Mesogasteropoda: Strombidae, Turbinellidae, Neptuneidae y Fasciolaridae) en Yucatán, México. Rev. Biol. Trop. 48: 145-152. [ Links ]
Pérez-Pérez, M. & D. Aldana-Aranda. 2003. Actividad reproductiva de Strombus gigas (Mesogasteropoda: Strombidae) en diferentes hábitats del Arrecife Alacranes,
Yucatán. Rev. Biol. Trop. 51: 119-126. [ Links ]
Ramirez, E. & C. Olabarria. 2005. Aspects of the distribution, population structure and reproduction of the gastropod Tibia delicatula (Nevill, 1881) inhabiting the oxygen minimum zone of the Oman and Pakistan continental margins. J. Sea Res. 54: 299-306. [ Links ]
Reed, S.E. 1995. Reproductive seasonality, periodicity and associated behavior in a colony of Strombus pugilis (Mollusca: Gasteropoda) in Puerto Rico. Am. Malacol. Bull. 11: 117-121. [ Links ]
Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish populations. J. Fish. Res. Board Can. 191: 1-382. [ Links ]
Sanders, I.M. 1988. Estimation of growth and population size of the fighting conch Strombus pugilis with a comparison of the Fabens and ELEFAN methods for estimating von Bertalanffy growth parameters. J. Shellfish Res. 7: 174. [ Links ]
Sparre, P. & S.C. Venema. 1997. Introducción a la evaluación de recursos pesqueros tropicales. FAO Documento Técnico de Pesca 306, Roma, Italia. [ Links ]
Stoner, A.W., V.J. Sandt & I.F. Boidron-Metairon. 1992. Seasonality in reproductive activity and larval abundance of queen conch Strombus gigas. Fish. Bull. 90: 161-170. [ Links ]
Tapia-Arjona, M.A. & D. Aldana-Aranda. 2007. Ciclo Reproductor del Gasterópodo Marino Melongena corona bispinosa (Neogastropoda: Melongenidae) en la Laguna de Chelem, Yucatán, México. Proc. Gulf. Carib. Fish. Inst. 59: 273-278. [ Links ]
Taylor, C.C. 1960. Temperature, Growth and Mortality-The Pacific Cockle. J. du Conseil 26: 117-124. [ Links ]
Von Bertalanffy, L. 1938. A quantitative theory of organic growth (inquiries on growth laws. II). Human Biol. 10: 181-213. [ Links ]
Weil, E. & R.A. Laughlin. 1994. Laboratory culture of Strombus gigas L. in the Dos Mosquises Marine Station, Los Roques National Park, Venezuela: Final results, p. 275-294. In R.S. Appeldoorn & B. Rodríguez (eds.). Strombus gigas Queen Conch Biology, Fisheries and Mariculture. Fund. Cient. Los Roques, Caracas, Venezuela.
Wolowicz, M. 1984. Cardium glaucum (Poiret, 1789) population from Gdansk Bay (Baltic Sea). Pol. Arch. Hydrobiol. 31: 33-44. [ Links ]
Zar, J.H. 1996. Biostatistical analysis. Prentice-Hall, Nueva Jersey, EEUU. [ Links ]
Zetina-Zárate, A.I., D. Aldana-Aranda, T. Brulé & E. Baqueiro-Cárdenas. 2000. Ciclo reproductor e índices de condición usados en Melongena corona (Mollusca: Gastropoda). Rev. Biol. Trop. 48: 77-86. [ Links ]
*Correspondencia a:
Josefina Santos Valencia. Laboratorio de Biología y Cultivo de Moluscos. Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Mérida, Carretera Antigua a Progreso, Km. 6, A.P. 73 Cordemex, C.P. 97310, Mérida, Yucatán, México; jsantos@mda.cinvestav.mx, Centro Regional de Investigaciones Pesqueras del Instituto Nacional de Pesca, Carretera Campeche-Lerma km. 5 S/N A.P. 140, C.P. 24500, San Francisco de Campeche, Campeche, México.
Martha Enríquez Díaz. Laboratorio de Biología y Cultivo de Moluscos. Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Mérida, Carretera Antigua a Progreso, Km. 6, A.P. 73 Cordemex, C.P. 97310, Mérida, Yucatán, México; menriquez@mda.cinvestav.mx
Dalila Aldana Aranda. Laboratorio de Biología y Cultivo de Moluscos. Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Mérida, Carretera Antigua a Progreso, Km. 6, A.P. 73 Cordemex, C.P. 97310, Mérida, Yucatán, México; daldana@mda.cinvestav.mx.
1. Laboratorio de Biología y Cultivo de Moluscos. Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Mérida, Carretera Antigua a Progreso, Km. 6, A.P. 73 Cordemex, C.P. 97310, Mérida, Yucatán, México; jsantos@mda.cinvestav.mx, menriquez@mda.cinvestav.mx, daldana@mda.cinvestav.mx
2. Centro Regional de Investigaciones Pesqueras del Instituto Nacional de Pesca, Carretera Campeche-Lerma km. 5 S/N A.P. 140, C.P. 24500, San Francisco de Campeche, Campeche, México.
Recibido 01-XII-2011. Corregido 04-VI-2012. Aceptado 05-VII-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}