











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.60 supl.2 San José Abr. 2012
Small-scale estimation of relative abundance for the coastal spotted dolphins (Stenella attenuata) in Costa Rica: the effect of habitat and seasonality
*Dirección para correspondencia
Abstract
The coastal spotted dolphin (Stenella attenuata graffmani) is one of the most common species of dolphin in inshore Pacific waters of Costa Rica. We conducted surveys in protected waters of the Papagayo Gulf, Costa Rica, to determine relative abundance of dolphins in relation to environmental variables. We used Generalized Additive Models to investigate the influence of a particular set of environmental factors and determine inter-annual trends in relative abundance. School sizes ranged from 1 to 50 individuals ( mean 9.95, SD=10.28). The number of dolphins increased linearly with water depth and transparency, and non-linearly with the dissolved oxygen concentration. High variability in the relative abundance occurred during the dry season (January-April). A previous study on this population found that high number of groups are involved in foraging activities during the dry season. Seasonal changes in relative abundance probably are associated with food availability, a variable that we did not measure. Understanding local resident populations may have important implications for conservation and management strategies. Large-scale studies may overlook variables affecting the abundance of local resident populations that may be detected with studies on a smaller scale such as this one.
Key words: Pantropical Spotted Dolphin; relative abundance; Habitat; Conservation; Pacific Ocean; Central America; Costa Rica, Cetacea.
Resumen
El delfín manchado costero (Stenella attenuata graffmani) es una de las especies de delfines mas comunes de las aguas costeras del Pacifico de Costa Rica. En este estudio realizamos muestreos dentro de las aguas protegidas del Golfo de Papagayo para determinar su abundancia relativa en relación a características físico-químicas de su hábitat. Usamos modelos aditivos generalizados para investigar la influencia de un juego de variables ambientales y determinar tendencias inter-anuales en la abundancia relativa. El tamaño de los grupos varió de 1 a 50 individuos (promedio 9.95, SD=10.28). La cantidad de delfines aumentó linealmente con la profundidad y claridad del agua, y de forma no lineal con el oxígeno disuelto. Durante la época seca (enero-abril) encontramos la mayor variabilidad en la abundancia relativa. Un estudio anterior sugiere que una cantidad importante de delfines se alimenta aquí en la época seca. Ello sugiere que los cambios estacionales en la abundancia relativa de delfines manchados costeros podría estar asociada con la disponibilidad de alimento, una variable que no medimos pero que podría explicar la variación observada debido a la naturaleza productiva estacional del área. Los estudios de poblaciones locales y residentes pueden tener un mayor impacto en estrategias de conservación y manejo. Por lo general, los estudios a mayor escala geográfica pueden pasar por alto variables importantes que afectan la abundancia local de poblaciones residentes, las cuales pueden ser detectadas en estudios de menor escala como el presente.
Palabras clave: Delfín manchado pantropical, abundancia, hábitat, conservación, Océano Pacifico, América Central, Población.
The spotted dolphin Stenella attenuata is one of the most common dolphin species in the eastern tropical Pacific (ETP) and at same time the most affected by the tuna industry (Dizon et al. 1994, Hall 1998). For management purposes, the species has been classified into four stocks based on morphological and geographical differences: Northerwestern, Southern offshore, Coastal, and Hawaiian (e.g., Perrin 1975, Douglas et al.1984, Schnell et al.1986, Perrin et al.1987, Dizon et al. 1994, Escorza-Treviño et al. 2005). The Northernwestern offshore stock is the most affected by the tuna fishery because of its association with the yellow-fin tuna (Thunnus albacares), and more efforts have been put in estimating the abundance and trends in abundance of the offshore than the Coastal or other stocks (e.g., Perrin et al.1982, Polacheck 1987, DeMaster et al. 1992, Wade & Gerrodette 1993, Fiedler & Reilly 1994, Wade 1995, Gerrodette & Palacios 1996, Archer et al. 2004, Gerrodette & Forcada 2005, Wade et al. 2007, Cramer et al. 2008, Archer et al. 2010).
The Coastal stock (S. attenuata graffmani), ranges within a 200 km band of coastal waters from southern México to Ecuador (Dizon et al. 1994). Genetic studies indicate the presence of several resident populations (Escorza-Treviño et al. 2005) along its distribution. Some of the latest abundance estimations include Gerrodette and Forcada (2006) using simulation approaches. They reanalyzed data collected over 21 years and found that coastal spotted dolphin abundances ranged between 96, 738 to 228, 038 dolphins, with a CV of 35%. The population trends for the offshore stock indicate a possible decline of the population over the past 21 years (Wade et al. 2007). However, estimations for the coastal stock have not been possible due to limitations in survey effort in coastal waters (Gerrodette & Forcada 2006). Abundance estimations of coastal spotted dolphins for the Exclusive Economical Zone of Costa Rica are limited to an old report in which relative abundance was estimated to be around 16, 600 dolphins, approximately 4.3-9.5% of the population size for all small cetaceans combined (Gerrodette & Palacios 1996).
Cetacean abundance is generally estimated with reference to a large regional-scale (e.g. Eastern Tropical Pacific); however for the coastal spotted dolphins, which have small distribution range, high residency times in regionally limited areas, and high annual mortality in fisheries operations, smaller scale studies are likely to provide more useful information for local conservation and management. Previous studies indicate that fluctuations in abundance of the Pantropical spotted dolphin in the Eastern Tropical Pacific are related to sea surface water temperature and thermocline depth throughout its distribution (Au & Perryman 1985, Reilly 1990, Reilly & Fielder 1994, Fieldler & Reilly 1994, Reilly et al. 2002). The role of local environmental variation in the population dynamics of dolphin resident populations is thus considered important in temperate waters, but very little has been documented in tropical waters (e.g., Reilly 1990, Garcia & Dawson 2003). Within the tropical surface water province in the Pacific Ocean, the Central American waters are considered the most variable, in which coastal upwellings are generated seasonally (Wyrki 1964, 1967, 1974, Au & Perryman 1985, Lizano 2008, Amador et al. 2006, Levin et al. 2006) and dolphin abundance is likely to respond to this environmental variation.
Generalized Additive Models (GAMs; Hastie & Tibshirani 1990) are useful to study abundance and distribution of highly mobile animals in relation with the environment at different scales, and in a more flexible way than linear analyses. The GAMs help determining whether apparent spatial and temporal trends represent real abundance changes or just natural fluctuations in distribution due to habitat variation (Forney 2000).
At the Pagayo Gulf, Costa Rica, coastal spotted dolphins appear to be resident year round (May-Collado 2001, 2009, May-Collado & Morales-Ramírez 2005, May-Collado et al. 2005, Martínez-Fernández et al. 2011). The gulf is characterized by two clearly marked seasons, dry and rainy, of which the dry season is characterized by strong northeast trade winds that generate the richest coastal upwelling of the country. Using GAMs, we modeled changes in relative abundance over time to evaluate how the local environment and seasonality can determine variations in resident populations of dolphins.
Methods
Study site: Murciélago Archipelago (N10°51’/W085°54’) is a set of old continental islands located in the northern part of the Papagayo Gulf, Costa Rica (Fig. 1). The area is affected by annual north-south migration of the inter-Tropical Convergent Zone (iTCZ); during the dry season (December-April) the iTCZ moves southward, the northeast trade winds are intensify generating superficial currents and the richest coastal upwelling of the country. During the second half of the year (rainy season, May to November) the iTCZ moves northward, the southeast trade winds bring rains, reducing salinity and intensifying the equatorial countercurrent near the coast of Central America (Wyrtki 1964, 1967, 1974, Amador et al. 2006, Lavin et al. 2006, Alfaro & Lizano 2001).
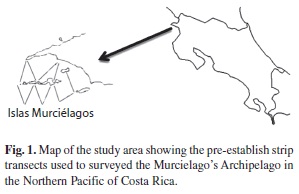
Sampling Methods: A total of 13 strip transects (1000 m x1000 m) were surveyed during the morning (6 a.m. to 12 p.m) and afternoon (1 p.m. to 6 p.m.) from January 1999 to November 2000, using small outboard engine boat. Because of the small range of the area sampled and the constant sighting cues of dolphin groups, we assumed total detection groups up to a distance of approximately 500 m at each side of the transect lines. This distance was the maximum distance at which dolphins were reliably detected from our small boat. We established this maximum distance using a range finder at different distances from a buoy. For every group detected we measured: sea surface temperature (Celsius) using a normal thermometer, concentration of dissolved oxygen (mg O2/L) was obtained using a dissolve oxygen meter (YSi 52), salinity was measured with a refractometer (ATAGO S/Mill-E) and water transparency (m) using a Secchi disk. in addition, thermocline depth was estimated in two stations that were monthly surveyed using a Niskin bottle we sampled the water column every 10 meters from the surface up to 50 m and measure dissolved oxygen, salinity, and temperature for each sample. Station Termo was located between the Archipelago and the mainland (N10º51.976’ - W085º54.283’, 40m deep) and station Catalina was located behind the main island, San Jose, facing offshore waters (N10º50.703’ - W085º55.016’, 50m deep). The thermocline was defined as a rapid change in the water temperature along the water column (Philander 1996, Thurman 1996). The thermocline occurs particularly in tropical and subtropical waters where it plays a significant role on the distribution of many marine organisms (Quesada-Alpizar and Morales-Ramirez 2004).
We also measured cloud cover using a scale of 0-8, sea state and wind speed based by discretely categorizing both as (1) zero with ocean surface flat and little wind, (2) light sea state when ocean surface had a few white caps, (3) moderate sea state, when ocean surface had considerably occurrence of white caps and, (4) strong sea state when wave action generated high waves, and many white caps. The data analyzed in this study was collected when sea state was zero and light. Finally, water depth was estimated from digital maps of the area with contour lines using ArcGiS tools.
Analysis
We used generalized additive models (Hastie & Tibshirani 1990) to evaluate changes in relative abundance corresponding to the total length of the transects surveyed as a function of environmental variables in a descriptive analysis, and second, we analyzed seasonal changes in relative abundance.
In the descriptive analysis, the estimated number of dolphins
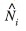 of each encountered school was the response variable to a set of environmental predictors
of each encountered school was the response variable to a set of environmental predictors 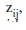 , where the j sub-script corresponds to the jth survey. Since the response described dolphin counts we assumed a Poisson distribution error in the model, and, because of the sparseness of our data we allowed for over-dispersion by specifying an error structure with variance proportional to the mean. We used a logarithmic function to link response and predictors. Our model had the general structure:
, where the j sub-script corresponds to the jth survey. Since the response described dolphin counts we assumed a Poisson distribution error in the model, and, because of the sparseness of our data we allowed for over-dispersion by specifying an error structure with variance proportional to the mean. We used a logarithmic function to link response and predictors. Our model had the general structure: 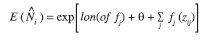
where “off” was an offset variable corresponding to the total length (sum) of the transects of survey j; θ is the intercept of the model to be estimated while fitting the data; and the fj are smoothed functions of the environmental covariates, which we chose to be smoothing splines. Smoothness, measured in degrees of freedom, was selected as a compromise between bias and variance of the fitted model, and described the linearity between
and . To select environmental covariates with high explanatory power of the response we used forward stepwise model fitting, starting with a general model including all possible covariates as linear terms and, at each step, one linear covariate was either replaced by a smoothed function with increasing degrees of freedom, from two to six, or was dropped from the model according to what option provided best fit of the model to the data. At each step, the best options were selected by the model with lowest Akaike information Criteria (AiC, Akaike 1973), a statistic that measures the best compromise between number of parameters, i.e. covariates or degrees of freedom of covariates, and an adequate description of the data.
To investigate seasonality, we modified a trend analysis based on GAMs developed by Fewster et al. (2000). in contrast to Fewster et al. (2000) analysis, we did not sample different sites, and we measured within season variability by replicated surveys of the same group of transects each month. Replicates within one month were considered independent, since were conducted at different times of the day. in our model, each survey was assumed to follow an independent Poisson distribution with mean number of dolphins μit in replicate i and month t. Within the GAM framework, month becomes an additive non-linear predictor:
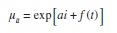
where αi is the replicate effect and f(t) was the month effect, estimated with a smoothing spline . With this model, the trend in the logarithm of the expected count over time is the same for each replicate; even absolute counts differed between replicates. An index of abundance over time is then constructed with the ratio of the estimates of
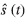 over
over 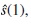 the estimated effect of the first month:
the estimated effect of the first month: 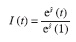
To provide a reasonable account of short term changes in dolphin counts, we selected smoothing splines with 7 degrees of freedom which revealed fine scale changes over time without decreasing unnecessarily the smoothing the output trend curve. To estimate the variance of this index we used a non-parametric bootstrap approach proposed by Fewster et al. (2000), in which replicated surveys in each month were treated as resampling units.
We estimated second derivatives of the smoothed index I(t) to identify major change points over time. At these points the second derivative is different from 0. if the second derivative was positive there was a change upwards, and if it was negative the change was downwards, i.e. a significant increase or decrease in abundance. The significant changes in the second derivatives were assessed with the approximate confidence intervals of the second derivatives, estimated from the bootstrap replicates of the indices.
Results
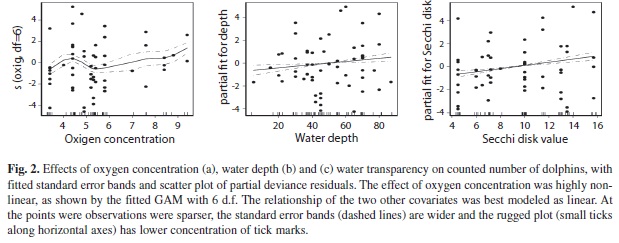
A total of 7,524 km of survey effort was completed in the two years of study and a high proportion of the surveys (62.5%) were completed with good sighting conditions (see methods). A total of 637 dolphins were seen in 64 groups. Group size ranged from 1 to 50 dolphins, two to five individuals being that most frequently observed. Mean group size was 9.95 (SD=10.28). The best-adjusted model retained three environmental variables: concentration of dissolved oxygen, water transparency and depth, based on the smallest AiC (Table 1, Fig. 2). The number of coastal spotted dolphins increased non-linearly with increasing superficial dissolved oxygen concentration and linearly with the water transparency and depth (Fig. 2). Despite of the variability in the data there is a trend in that coastal spotted dolphins seem to aggregate in relatively larger schools in clearer and deeper waters around the islands (Fig. 2).
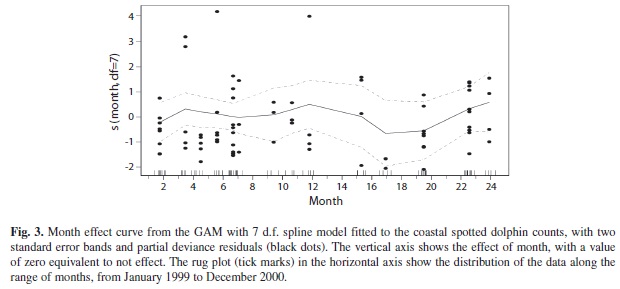
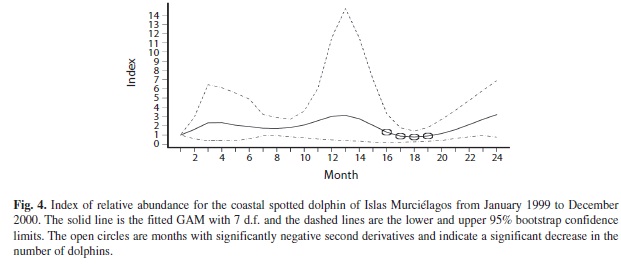
The trend analysis confirmed the effect of the month in the dolphin’s counts, showing a gradual decrease in the counts from the dry season to mid-rains (Fig. 3). The wider stan dard error bands show large within-season variability, as estimated by the 95% bootstrap confidence intervals (Fig. 4). The largest variability is observed during the dry season in both years and the lowest during the months of rain. Overall, relative abundance of spotted dolphins varied annually, increasing in numbers at the end of the rainy season and reaching its peak in the dry season, during the months of upwelling. Afterwards, during the early and mid-rainy season there was a dramatic decrease in dolphin abundance. These results indicate that coastal spotted dolphins relative abundance varies annually in response to the environmental seasonality of the habitat.
Furthermore, differences in the months in which abundance reached its highest and lowest values suggest that dolphin inter-annual variation may be occurring as well. During 2000 the largest variability in abundance was in January, followed by a decrease in the number of dolphins during the early and midrainy season (May to September). This decrease is revealed by significant upturns of the trend line (Fig. 2) from April to July of 2000, which could be detected given the low variability in dolphin counts during these months. in July 2000 the relative abundance of the dolphins increased again and continued to increase throughout the last months of rains, and during the dry season. During the previous year, the higher number of dolphins was observed in March, and a significant decrease occurred from July to September. The abundance of dolphins started to increase in October.
Discussion
Generalized Additive Models are an effective way to detect environmental variables that best predict changes in the relative abundance of the coastal spotted dolphin in the waters surrounding the Murciélago islands, Costa Rica. These models may allow for a better detection of seasonal trends in abundance of small study areas. Month effects were not significant probably because each month represented a small time window to detect changes in abundance. The trend analysis, however, clearly indicated a seasonal pattern both in relative number of dolphins and its variability.
Regional scale seasonal variation in tropical waters plays an important role in the relative abundance of coastal spotted dolphins as suggested by our models; relative abundance of dolphins increased linearly with water depth and water transparency, and non-linearly with the concentration of dissolved oxygen. These results did not indicate direct environmental forcing of dolphin abundance. instead these environmental factors appear to be associated with the onset of the dry season where the highest relative abundance occurred, from December to April.
The Pacific waters of Costa Rica are part of the tropical waters of the “Central American Bight” (near coastal waters from Guatemala to Ecuador). These waters are considered the most variable within the Tropical Surface Water Province due to the influence of the inter-Tropical Convergence Zone (iTCZ) which annual migration generates dramatic changes in the oceanography of the area (Au & Perryman 1985, Rasmusson & Akin 1993). Moreover, these waters constitute the most important area of overlap for spotted, spinner, striped and common dolphins (Au & Perryman 1985). Our studied area is probably the most affected by the iTCZ in Costa Rica when this feature is located in its most southward position (dry season) and the northeast trade winds are intensified generating the richest upwelling of the country. An approximate five-fold increase in the biomass and zooplankton abundance occurs during the dry season, compared to the rainy season, due to a seasonal upwelling (Bednarski & Morales-Ramirez 2004). Thus, we propose that this well marked seasonal variation in the environment may be influencing prey availability and therefore affects the presence of dolphins in the area.
in agreement with this suggestion, MayCollado & Morales-Ramírez (2005) reports that seasonal changes in behavioral patterns and abundance of dolphins in the Gulf of Papagayo are likely to be associated with that increase in zooplankton biomass. They reported at high number of groups involved in foraging behaviors, and a high investment in foraging activities during the dry season. Other predators seem to respond as well to seasonal changes, for example the normally pelagic mahi-mahi (Coryphaena hippurus), which feeds on prey similar to that of dolphins, approaches the shore during the dry season, to feed (Allen & Robertson 1998). Very little is known about the feeding habits of coastal spotted dolphins. in the Eastern Tropical Pacific the oceanic ecotype is consider a generalist feeder, which distribution seems to be largely influenced by prey availability (Perrin et al. 1973, Fieldler et al. 1998). These changes in tropical areas might be particularly more intense in local populations, perhaps not as dramatic as in local temperate dolphin populations, but sufficient to have an impact in the abundance and distribution of the dolphins and their prey. Furthermore, more recent studies comparing northern and southern populations of spotted dolphins of the Pacific coast of Costa Rica have found similar results, primarily the relationship between water transparency and dolphin abundance (Martínez-Fernández et al. 2011).
in conclusion, Generalized Additive Models helped us in identifying what factors might be driving small-scale dolphin abundance and what are the implications of seasonality in dolphin abundance for conservation and management strategies of local resident populations. Large-scale studies may overlook variables affecting the abundance of local resident populations that may be detected with studies on smaller scale such as this one. Genetic studies on the coastal spotted dolphins suggest that these dolphins are highly resident and are likely to have limited distribution ranges (Escorza-Treviño et al. 2005) subject to seasonal variation. Therefore, designing conservation strategies relies on the knowledge of their occurrence and seasonal use of important areas.
Acknowledgments
Funding and logistic support was provided by The Nature Conservancy, Emily Shane Award 1998, Society of Marine Mammals, and Vicerrectoria de investigación and Centro de investigación en Ciencias del Mar y Limnología (CiMAR) of the Universidad de Costa Rica, and finally to the Parque Nacional Santa Rosa, Area de Conservación Guanacaste. For assistance on the field we thank F. Lara (father and son), Samba, M. Lara, Guicha, M. Quesada, and many students that volunteer along the way. Thanks to Alvaro Morales, Roger Blanco, Frank Joyce, Doña Carmen for all their support along the study. Thanks to ingi Agnarsson and Tim Gerrodette for important comments to the manuscript.
References
Akaike, H. 1973. information and an extension of the maximum likelihood principle, Pp. 267-281. In: B.N. Petran & F. Csáki (eds). international Symposium on information Theory. Akadémiai Kiadi Press, Budapest, Hungary. [ Links ]
Archer, F. i., Gerrodette, T., Chivers, S., & A. Jackson. 2004. Annual estimates of the unobserved incidental kill of pantropical spotted dolphin (Stenella attenuata attenuata) calves in the tuna-seine fishery of the eastern tropical Pacific. Fish. Bull. 102: 233-244. [ Links ]
Archer, F. i., Redfern, J. V., Gerrodette, T., Chivers, S. J., & W. F. Perrin. 2010. Estimation of relative exposure of dolphins to fishery activity. Mar. Ecol. Prog. Ser. 410: 245-255. [ Links ]
Alfaro, E.J. & O.G. Lizano. 2001. Some relationships between upwelling zones of the Central American Pacific and tropical Atlantic and Pacific oceans. Rev. Biol. Trop. 49: 187-193. [ Links ]
Allen, G.R. & D.R. Robertson. 1998. Peces del Pacífico Oriental Tropical. CONABiO Press, Distrito Federal, Mexico. [ Links ]
Amador, J.A., Alfaro, E. J. & O.G. Lizano. 2006. Atmospheric forcing of the eastern tropical Pacific: a review. Progr. Oceanog. 69: 101-142. [ Links ]
Archer, F.i., J.V. Redfern, T. Gerrodette & W.F. Perrin. 2010. Estimation of relative exposure of dolphins to fishery activity. Mar. Ecol. Prog. Ser. 410: 245-255. [ Links ]
Au, D.W.K. & W.L. Perryman. 1986. Dolphin habitats in the eastern tropical Pacific. Fish. Bull. 83: 623-643. [ Links ]
Bednarski, M. & A. Morales-Ramirez. 2004. Composition, abundance, and distribution of macrozooplankton in Culebra Bay, Gulf of Papagayo, Pacific coast of Costa Rica and its value as bioindicator of pollution. Rev. Biol. Trop. 52: 105-119. [ Links ]
Cramer, K.L., W.L. Perryman & T.Gerrodette. 2008. Declines in reproductive output in two dolphin populations depleted by the yellowfin tuna purse seine fishery. Mar. Ecol. Prog. Ser. 369: 273-285. [ Links ]
De Master, D.P., E.F. Edwards, P. Wade & J.E. Sisson. 1992. Status of dolphin stocks in the eastern tropical Pacific, Pp.1038-1050. In: D.R. MaCullough & R.H. Barrett (eds). Wildlife 2001: Populations. Proceedings of an international conference on population dynamics and management of vertebrates (exclusive of primates and fish) (July 29-31, 1991, Oakland, California). Elsevier, New York, USA. [ Links ]
Dizon, A.E., W.F. Perrin & P.A. Akin. 1994. Stocks of dolphins (Stenella spp. and Delphinus delphis) in the eastern tropical Pacific: a phylogeographic classification. U.S. Department of Commerce. NOAA Technical Report NMFS 119: 20p. [ Links ]
Douglas, M.E. G.D. Schnell & D.J. Hough. 1984. Differentiation between inshore and offshore spotted dolphins in the eastern tropical Pacific Ocean. J. Mammal. 65: 375-387. [ Links ]
Escorza-Trevino S., F.i. Archer, M. Rosales, A. Lang & A.E. Dizon. 2005. Genetic differentiation and intraspecific structure of eastern tropical Pacific spotted dolphins, Stenella attenuata, revealed by DNA analyses. Conserv. Genet. 6: 587-600. [ Links ]
Fewster, R.M., S.T. Buckland, G.M. Siriwardena, S.R. Baillie & J.D. Wilson. 2000. Analysis of population trends for farmland birds using generalized additive models. Ecology 81: 1970-1984. [ Links ]
Fiedler, P.C. & S.B. Reilly. 1994. interannual variability of dolphin habitats in the eastern tropical Pacific. ii: effects on abundances estimated from tuna vessel sightings, 1975-1990. Fish. Bull. 92: 451-463. [ Links ]
Fiedler, P. C., Barlow, J. & T. Gerrodette. 1998. Dolphin prey abundance determined from acoustic backscatter data in eastern Pacific surveys. Fish. Bull. US 96: 237-247. [ Links ]
Forney, K.A. 2000. Environmental models of cetacean abundance: reducing uncertainity in population trends. Conserv. Biol. 14: 1271-1286. [ Links ]
Garcia, C. & S. M. Dawson. 2003. Distribution of pantropical spotted dolphins in Pacific coastal waters of Panama. LAJAM. 21: 1676-7497. [ Links ]
Gerrodette, T. & D.M. Palacios. 1996. Estimates of cetacean abundance in EEZ waters of the eastern tropical Pacific. Southwest Fisheries Science Center Administrative Report LJ-96-10. 28 p. [ Links ]
Gerrodette T. & J. Forcada. 2005. Non-recovery of two spotted and spinner dolphin populations in the eastern tropical Pacific Ocean. Mar. Ecol. Prog. Ser. 291: 1-21. [ Links ]
Gerrodette, T. & J. Forcada. 2006. Estimates of abundance of northeastern offshore spotted, coastal spotted, and eastern spinner dolphins in the eastern tropical Pacific Ocean. Southwest Fisheries Science Center Administrative Report LJ-02-06, La Jolla, California 41p. [ Links ]
Hall, M.A. 1998. An ecological view of the tuna-dolphin problem:impacts and trade offs. Rev. Fish. Biol. Fish. 8: 1-34. [ Links ]
Hastie, T.J. & R.J. Tibishirani. 1990. Monographs on Statistics and Applied Probability: Generalized Additive Models. Chapman and Hall Press, Florida, USA. [ Links ]
Levin, M.F., Fiedler, P.C. & J.A. Amador. 2006. A review of eastern tropical Pacific oceanography: summary. Progr. Oceanog. 69:391-398. [ Links ]
Lizano, O.G. 2008. Water dynamics around Cocos island, Costa Rica. Rev. Biol. Trop. 56: 31-48. [ Links ]
Martínez-Fernández, J.D., A. Montero-Cordero & L.J. May-Collado. 2011. Cetaceos de las aguas costeras de Pacifico norte de Costa Rica. Rev. Biol. Trop. 59: 283-290. [ Links ]
May-Collado, L.J. 2009. Marine Mammals, p. 479-496. In: i.S. Wehrmann & J. Cortes (eds). Marine Biodiversity of Costa Rica, Central America. Springer, The Netherlands. [ Links ]
May-Collado, L.J. & A. Morales-Ramírez. 2005. Presencia y patrones de comportamiento del delfín manchado costero, Stenella attenuata graffmani (Cetacea: Delphinidae) en el Golfo de Papagayo, Costa Rica. Rev. Biol. Trop. 53: 265-276. [ Links ]
May-Collado, L.J., T. Gerrodette, J. Calambokidis, K. Rassmusen & i. Sereg. 2005. Distribution of cetaceans sightings in the EEZ of Costa Rica. Rev. Biol. Trop. 53: 249-263. [ Links ]
Schnell, G.D., M.E. Douglas & D.J. Hough. 1986. Geographic patterns of variation in offshore spotted dolphins (Stenella attenuata) of the eastern tropical Pacific ocean. Mar. Mamm. Sci. 2: 186-213. [ Links ]
Oviedo, L. 2007. Dolphin sympatric ecology in a tropical fjord: habitat partitioning by bathymetry and topography as a strategy to coexist. J. Mar. Biol. Assoc. U.K. 87: 1327-1335. [ Links ]
Philander, G. 1996. El Niño and La Niña, p. 72-87. In R.G. Pirie (ed). Oceanography: contemporary readings in ocean science. Oxford Univ. New York, N.Y. [ Links ]
Perrin, W.F., R.R. Warner, C.H. Fiscus & D.B. Holts. 1973. Stomach contents of porpoise Stenella spp. and yellowfin tuna, Thunnus albacares, in mixed-species aggregations. Fish. Bull. 71: 1077-1092. [ Links ]
Perrin, W.F., T.D. Smith & G.T. Sakagawa. 1982. Status of populations of spotted dolphin, Stenella attenuata, and spinner dolphin, S. longirostris in the eastern tropical Pacific. Mammals in the Seas, FAO. Fisheries Series No.5, Vol.iV, ISBN 92-5-100514-1. [ Links ]
Perrin, W.F., M.D. Scott, J.G. Walker & V.L. Cass. 1985. Review of geographical stocks of tropical dolphins (Stenella spp. and Delphinus delphis) in the eastern Pacific. U.S. Department of Commerce. NOAA Technical Report NMFS 28: 28p. [ Links ]
Perrin, W.F., E.D. Mitchell, J.G. Mead, D.K. Caldwell, M.C. Caldwell, P.J.H. Van Bree & W.H. Dawbin. 1987. Revision of the spotted dolphins Stenella spp. Mar. Mamm. Sci. 3: 99-170. [ Links ]
Polacheck, T. 1987. Relative abundance, distribution and inter.specific relationship of cetacean schools in the eastern tropical Pacific. Mar. Mamm. Sci. 3: 54-77. [ Links ]
Quesada-Alpizar, M. A. & A. Morales-Ramirez. 2004. Comportamiento de las masas de agua en el GolfoDulce, Costa Rica durante El Nino (1997-1998). Rev. Biol. Trop. 52(2): 95-103. [ Links ]
Rasmusson, E. M. & P. A. Akin. 1993. A global view of large-scale precipitation variability. J Clim. 6: 1495-1522. [ Links ]
Reilly, S.B. 1990. Seasonal changes in distribution and habitat differences among dolphins in the eastern tropical Pacific. Mar. Ecol. Prog. Ser. 66: 1-11. [ Links ]
Reilly, S.B. & P.C. Fiedler. 1994. interannual variability of dolphin habitats in the eastern tropical Pacific i: Research vessel surveys 1986-1990. Fish. Bull. 92: 451-463. [ Links ]
Thurman, H.V. 1996. Essentials of Oceanography. Prentice Hall, New Jersey, New Jersey. 400 p. [ Links ]
Wade, P.R. 1995. Revised estimates of incidental kill of dolphins (Delphinidae) by the purse-seine tuna fishery in the eastern tropical Pacific, 1959-1972. Fish. Bull. 93: 345-353. [ Links ]
Wade, P.R. & T. Gerrodette. 1993.Estimates of Cetacean abundance and distribution in the eastern tropical Pacific. Rep. int. Whal. Commn. 43: 477-483. [ Links ]
Wade, P. R., Watters, G. M., Gerrodette, T. & S. B. Reilly. 2007. Depletion of spotted and spinner dolphins in the eastern tropical Pacific: modeling hypotheses for their lack of recovery. Mar. Ecol. Prog. Ser. 343: 1-14. [ Links ]
Wyrtki, K. 1964. Upwelling in the Costa Rica Dome. U.S. Fishery and Wildlife Service. Fish. Bull. 63: 355-372. [ Links ]
Wyrtki, K. 1967. Circulation and water masses in the eastern equatorial Pacific Ocean. int. J. Oceanol. Limnol. 1: 117-147. [ Links ]
Wyrtki, K. 1974. Equatorial currents in the Pacific 1950 to 1970 and their relations to the trade winds. J. Phys. Oceanogr. 4: 372-380. [ Links ]
*Correspondencia a:
Laura J. May-Collado.University of Puerto Rico, Department of Biology, San Juan, Puerto Rico, 00931¸lmaycollado@gmail.com. George Mason University, Department of Environmental Science & Policy, MSN 5F2, 4400 University Drive, Fairfax, Virginia, USA 22030
Jaume Forcada. NERC, British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 0ET, United Kingdom; jfor@bas.ac.uk
1. University of Puerto Rico, Department of Biology, San Juan, Puerto Rico, 00931¸lmaycollado@gmail.com
2. George Mason University, Department of Environmental Science & Policy, MSN 5F2, 4400 University Drive, Fairfax, Virginia, USA 22030
3. NERC, British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 0ET, United Kingdom; jfor@bas.ac.uk
Received 22-IV-2011. Corrected 01-XII-2011. Accepted 15-II-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}