










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 suppl.1 San José Mar. 2012
Coral recruitment to two vessel grounding sites off southeast Florida, USA
*Dirección para correspondencia
Abstract
Over the last two decades, more than 10 major vessel groundings have occurred on coral reefs offshore southeast Florida. Lack of any published information on coral settlement, post-settlement survival, and juvenile coral growth in the southeast Florida region inhibits efforts to determine if coral populations will be able to effectively re-establish themselves. The goal of this study was to examine these processes to obtain background data needed to determine the potential for natural recovery. Over a three year period annual coral recruitment, juvenile growth, and mortality rates were measured in 20 permanent quadrats at each of two ship grounding and two control sites. The density of new recruits was generally low, ranging from 0.2±0.1 (SE) to 7.1±1.0 recruits m-2. Although the density of coral recruits was generally higher at the grounding sites, mortality rates were high at all sites during the study period. Growth rates of individual colonies were highly variable, and many of the colonies shrank in size due to partial mortality. Results indicate that corals are able to recruit to the damaged reefs but that slow growth rates and high mortality rates may keep these areas in a perpetual cycle of settlement and mortality with little or extremely slow growth to larger size classes, thus inhibiting recovery.
Key words: recruitment, reef recovery, vessel groundings, growth rates, post settlement survival.
Resumen
En las dos últimas décadas, más de 10 grandes encallamientos de embarcaciones se han producido en los arrecifes de coral mar afuera en el sureste de Florida. La falta de información publicada sobre el asentamiento de corales y sobrevivencia post-asentamiento y de los corales juveniles que crecen en la región, limita los esfuerzos para determinar si las poblaciones de coral serán capaces de restablecerse por ellas mismas. El objetivo de este estudio fue examinar estos procesos para obtener la información necesaria para determinar el potencial de recuperación natural. Se midió el reclutamiento anual de coral joven, el crecimiento y las tasas de mortalidad por un período de tres años, mediante 20 cuadrantes permanentes en cada uno de los dos encallamientos de barcos y dos sitios de control. La densidad de nuevos reclutamientos fue generalmente baja, de 0.2±0.1 a 7.1±1.0 reclutamientos m-2. Aunque la densidad del reclutamiento fue generalmente más alta en los sitios de encallamiento también hubo mayor mortalidad de corales juveniles en esos sitios durante el período de estudio. Las tasas de crecimiento de las colonias individuales fueron altamente variables, y muchas de las colonias se redujeron en tallas debido a mortalidad parcial. Los resultados indican que los corales presentan una disposición a reclutarse en arrecifes dañados, pero las lentas tasas de crecimiento y la alta mortalidad pueden mantener esas áreas en un perpetuo ciclo de asentamiento y mortalidad con poco o extremadamente lento crecimiento hacia las clases de tallas largas, por lo tanto inhibiendo la recuperación.
Palabras clave: reclutamiento, recuperación de coral, encallamiento de barcos, tasas de crecimiento, sobrevivencia post-asentamiento
The southeast Florida reef tract consists of four shore-parallel ridges with a diverse coral reef community similar to that found in the Western Atlantic and Caribbean (Moyer et al. 2003, Banks et al. 2008). Though similar in fauna, community structure of this high latitude (26° N) region differs from that found in other areas of the Caribbean and Florida Keys (Moyer et al. 2003). Montastraea cavernosa Linnaeus, 1767 replaces the Montastraea annularis (Ellis & Solander, 1786) species complex as the dominant species. Additionally, average coral colony size is typically small, and cover is generally low (Moyer et al. 2003).
Coral reefs in southeast Florida are located near a highly developed and heavily populated coastline. As a result, they are subjected to a number of anthropogenic stressors, one of which is physical impact from vessel groundings and anchor damage. Over the past 17 years, at least ten large ship groundings and six anchor damage events associated with the Port Everglades anchorage area have occurred near Ft. Lauderdale, Florida. Impacts include flattening of reef topography, removal of living benthic resources, and creation of large rubble fields. Restoration of these sites has been generally limited to reattachment of scleractinian corals. Removal of rubble was initiated at grounding sites occurring in 2004 and later in an effort to reduce further damage due to high mobility of debris that could hinder recovery of benthic communities. In general, there has been little attempt to restore topography at the damaged sites with the exception of some localized efforts initiated in 2004 that have accompanied rubble stabilization (e.g., cementing larger sized rubble created by the grounding of the M/V Eastwind) (Marine Resources Inc. 2005). However, efforts have been relatively small scale as compared to the larger scale efforts to stabilize reef framework and restore reef topography further south in the Florida Keys that have included use of artificial structures such as prefabricated cement modules and placement of limestone boulders (Jaap et al. 2006).
Recovery of damaged sites is generally defined as the return of both community structure and function to a state similar to that which existed pre-impact (Edwards & Gomez 2007). Although there are thousands of organisms which contribute to this structure and function, scleractinian corals are often used as indicators of recovery since they are the main builders of habitat. One of the first steps of recovery after damage has occurred is the recruitment of new colonies to the area. Re-establishment of coral populations is highly dependent on settlement of coral planulae, post-settlement survival, and juvenile coral growth. However, coral recruitment in the Caribbean and western Atlantic is typically low (Rogers et al. 1984, Hughes & Jackson 1985, Smith 1997, Miller et al. 2000). Furthermore, slow growth rates of generally less than 1 cm yr-1 for many Caribbean species (Bak & Engel 1979, Rogers et al. 1984, van Moorsel 1988, Edmunds 2000) can influence recovery times. Currently no information on coral recruitment rates or juvenile survival and growth rates has been published for southeast Florida. Therefore, the goal of this study was to examine these processes on a small number of sites off Ft. Lauderdale, Florida to obtain background data needed to determine the potential for natural recovery from damage due to vessel groundings that have occurred in this area.
Methods
Two ship grounding sites were chosen for study: the Eastwind (26°7’2”N, 80°5’32”W) and Federal Pescadores (26°6’44”N, 80°5’30”W). The M/V Eastwind grounded in March 2004 and resulted in approximately 11 000 m2 of damage on the inner reef just north of Port Everglades (Hudson Marine Management Services 2004a). The M/V Federal Pescadores created approximately 23 400 m2 of damage 0.4 km south of the Eastwind grounding site when it ran aground in October 2004 (Hudson Marine Management Services 2004b). Both groundings destroyed the existing relief, creating large rubble fields from which large rubble was subsequently removed or stabilized during restoration activities (Marine Resources Inc. 2005), but smaller sized debris remained on site.
Two control sites were chosen for comparison to the grounding sites. Because most of the large vessel groundings and anchor damage events off Ft. Lauderdale have occurred just west of the Port Everglades anchorage, this area was avoided when control sites were selected due to the potential for previous impact. Using GIS, a grid consisting of 15x15 m plots was overlaid on the inner reef just north of the anchorage area, and two plots were randomly picked for the control sites (control sites one: 26°9’37”N, 80°5’18”W and two: 26°10’4”N, 80°5’15”W).
Twenty permanently marked 1 m2 quadrats at each of the four sites were surveyed annually between May 2006 and June 2009. Quadrats were marked at the corners using a stainless steel pin and galvanized nails. A 1 m2 pvc frame subdivided into 100 squares 10x10 cm in size was placed over the pins to accurately locate corals within the quadrat. All juvenile corals (≤5 cm in diameter) in each quadrat were identified, measured, and mapped. Juvenile corals appearing in the quadrats that were not present in the previous years and were not a product of colony fission were considered coral recruits. Any corals that appeared in the quadrats but were absent in subsequent years were assumed to be dead. The change in maximum diameter of all corals that appeared in multiple years was calculated and used for measures of coral growth.
Differences in density of recruits among sites and among years were tested using a two-way ANOVA on ranks after data transformations failed to achieve normality due to the presence of multiple zero values. Differences in average annual mortality among sites were tested using ANOVA. Post hoc pair-wise comparisons were performed with Tukey tests when significant differences were found.
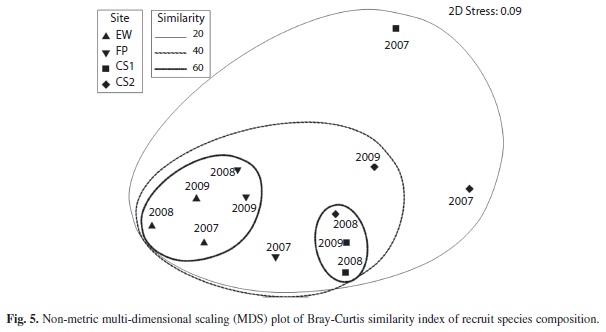
To further examine similarity in species composition of recruits among sites, a Bray-Curtis similarity index was determined. Species abundance data were pooled for all quadrats within a site during a survey year, and the data were square-root transformed (PRIMER v6). The results were also used to construct a non-metric, multi-dimensional scaling (MDS) plot.
Results
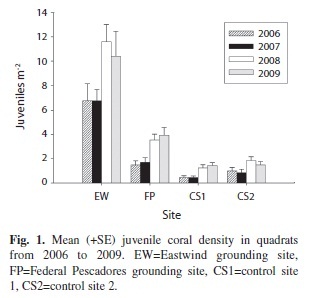
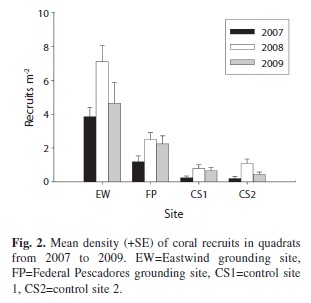
Mean juvenile coral density ranged from a low of 0.5±0.1 (SE) juveniles m-2 at control site 1 to a high of 11.6±1.4 juveniles m-2 at the Eastwind grounding site (Fig. 1). The density of recruits ranged from 0.2±0.1 recruits m-2 at control site 2 to 7.1±1.0 recruits m-2 at the Eastwind grounding site (Fig. 2). The results of a two-way ANOVA on ranks indicated that there was a significant difference in density of new recruits both among sites (p<0.001) and years (p<0.001). A Tukey pair-wise comparison test indicated that the two grounding sites each
had a significantly higher density of recruits than either control site (p<0.001) and that density of recruits was significantly higher at the Eastwind grounding site than at the Federal Pescadores grounding site (p<0.001). There was no significant difference in recruit density between the two control sites. Recruit density was significantly higher in 2008 than in 2007 (Tukey, p<0.001) or 2009 (p<0.01).
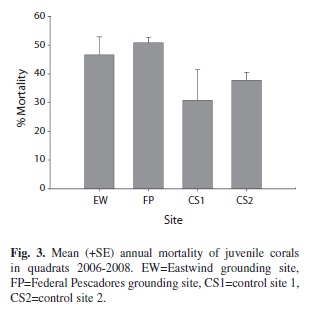
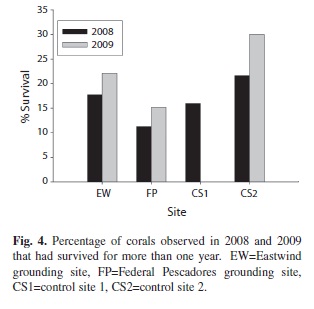
Annual mortality rates of juvenile corals ranged from 11% to 58%. Mean annual mortality rates were higher at the grounding sites than the control sites during the study period, but the difference was not significant (Fig. 3). Survival of juvenile corals for longer than one year ranged from 0 to 30% of the total corals
observed at each site in 2008 and 2009 (Fig. 4).
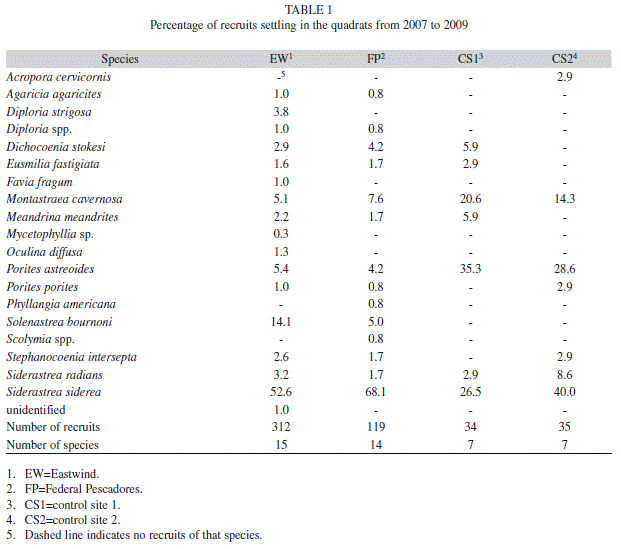
A total of 18 species recruited to the study sites, but the maximum number of species that recruited to any one site was 15 (Table 1). Twice as many species recruited to the grounding sites compared to the control sites (Table 1). Four common species, Siderastrea siderea (Ellis & Solander 1786), Siderastrea radians (Pallas 1766), Porites astreoides Lamarck, 1816, and Montastraea cavernosa, recruited to all the sites. Over half of the recruits at the grounding sites were S. siderea. Porites astreoides and S. siderea comprised a majority of the total recruits (64% to 67% combined) at the control sites though M. cavernosa also contributed to a relatively large portion (14% to 21%) compared to the grounding sites (5% to 8%).
A non-metric multi-dimensional scaling (MDS) plot was used to visualize the Bray-Curtis similarity of sites based on species abundance of recruits (Fig. 5). Two main clusters with 60% similarity were formed, one consisting of all the grounding sites (minus the 2007 Federal Pescadores) and the other consisting of 2008 and 2009 control site1 and 2008 control site 2. The 2007 Federal Pescadores data and 2009 control site 2 clustered with the above mentioned data at 40% similarity, and the 2007 data for both control sites clustered with the rest of the sites at 20% similarity.
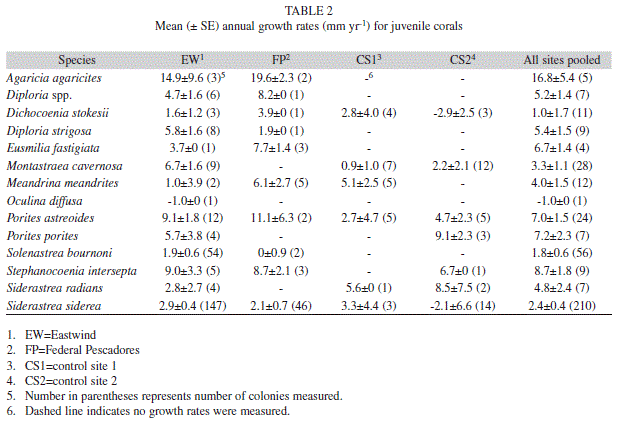
Growth rates were highly variable both within and between species and often between sites (Table 2). Most species grew less than 1 cm yr-1 except for Agaricia agaricites (Linnaeus 1758) which was only observed at the grounding sites. Many colonies (23%) suffered partial mortality, resulting in smaller colony size in subsequent years, and 6% of the colonies did not grow between survey periods. For some species, the number of corals measure has been observed at higher latitudes (Hughes et al. 2000, St. Gelais 2010), which may contribute to lower recruitment rates. Differential recruitment among sites in the current study is probably not impacted by larval supply since recruitment was significantly higher at the grounding sites compared to the controls. Higher recruitment to the grounding sites may be the result of the greater availability of less-colonized substrate. The presence of well-established communities at the control sites may have resulted in lower settlement through the preemption of space and lower survival through competitive interactions (Birkeland 1977, Bak & Engel 1979, Maida et al. 2001, Kuffner et al. 2006, Vermeij 2006, Arnold et al. 2010).
As is the case in other studies (Rylaarsdam 1983, Babcock 1985, van Moorsel 1988, Vermeij 2006), growth rates of individual colonies in this study were highly variable. Nevertheless, mean growth rates were within the range reported in the literature though on the lower end for most species (Bak & Engel 1979, van Moorsel 1988, Edmunds 2000). The vast majority of colonies grew less than 1 cm yr-1 . However, many of the colonies (23%) experienced negative growth and shrank in size due to partial mortality. Average colony size in the region tends to be small (< 50 cm, Moyer et al. 2003) in comparison to other areas, and partial mortality and slow growth rates observed in the current study likely contribute to these patterns.
Although mean annual mortality rates were not significantly different among sites, total mortality of juvenile corals was about 50% at the grounding sites and 40% at the control sites during the study period, indicating a slightly higher mortality of juvenile corals at the grounding sites over the three year study period. Additionally, the percentage of corals that survived more than one year was generally low (< 30%), indicating low longer-term (> 1 year) survival across both grounding and control sites.
Low survival of corals was likely mediated by different processes at the grounding and control sites due to the differences in habitat. Although no physical measurements were taken, qualitatively it appeared that sand and rubble movement occurred at the grounding sites at an increased rate in comparison to undisturbed sites. Some of the quadrats that were initially established on hard substrate at the grounding sites were noted to be covered with sand and rubble during subsequent surveys. The movement and accumulation of sand and rubble was likely facilitated by the flattening and destruction of the relief by the grounded ships. The presence and movement of sand and rubble likely inhibited recruit survival at the grounding sites through both burial and scour (Bak & Engel 1979).
Low survival at the control sites may have been affected by competitive interactions with more established benthic organisms and algae (Birkeland 1977, Bak & Engel 1979, Maida et al. 2001, Kuffner et al. 2006, Vermeij 2006). Though no quantitative data of percent cover were taken within the quadrats, visually the grounding sites appeared more barren in comparison to the control sites (Fig. 6). In contrast, Thanner et al. (2006) found that community structure on artificial reefs in southeast Florida reached between 45% to 58% similarity to natural reefs in benthic species composition 4 to 5 years after deployment and that scleractinian corals on the artificial reefs had similar cover, though higher abundance of juvenile corals, compared to the natural reef after the same period. The impoverished benthic community at the grounding sites 5 years post-grounding provides a sharp contrast to the moderately high similarity in benthic community structure between the natural reef and artificial reef structures 5 years post deployment that Thanner et al. (2006) reported and indicates that topographic complexity may be important for the recovery of the grounding sites.
Community structure of newly settled recruits was distinct at the grounding sites. More species recruited to the grounding sites (14-15 species) compared to the reference sites (7 species), but settlement was heavily dominated (53-68%) by S. siderea. Siderastrea spp. are able to live in conditions generally considered inimical to coral survival such as fluctuating salinity, low temperatures, and high sedimentation (Macintyre & Pilkey 1969, Muthiga & Szmant 1987, Lirman & Manzello 2007). Their dominance at the grounding sites might be a result of their tolerance to the high sedimentation observed due to the destruction of topography from the ship groundings. Montastraea cavernosa, the dominant species in the region (Moyer et al. 2003), comprised a much smaller proportion of the recruits at the grounding sites (<8%) compared to the reference sites (14-20%).
The concern is that even though corals are recruiting to the grounding sites, they are not surviving and growing to larger size classes. This phenomenon has been observed in other studies where corals have recruited to damaged sites but have failed to achieve larger size classes even on a decadal time scale (Gittings et al. 1990, Rogers 1984, Cook et al. 1994, Rogers & Garrison 2001). Corals that have low recruitment and growth rates, as many of the broadcast spawning, reef building corals do, may take even longer to recover as models have determined that convergence to pre-disturbance levels are highly dependent on recruitment rates (Hughes & Tanner 2000, Lirman & Miller 2003). Given the pattern of low recruitment and slow growth of the broadcast spawning corals observed in the present study, natural recovery is likely to be slow. Additionally, the low survival of all corals beyond a year observed at the study sites will contribute to slow recovery.
Recovery is generally defined as the return of community structure and function of the injured site to conditions similar to those that existed pre-disturbance (Edwards & Gomez 2007). However, injured sites have been known to recover to alternate community states distinct from those present pre-disturbance (Hatcher 1984). Aronson and Swanson (1997) found that a decade after the injury occurred, the Wellwood grounding site in the Florida Keys was more similar to a hard bottom community than to its original spur and groove structure. The grounding sites in the current study were visually distinct from control sites. They were noticeably flat and devoid of larger coral colonies and other benthic invertebrates such as sponges and gorgonians common at the control sites. Though return of benthic community structure, if possible, is expected to take on the order of decades, return of topography is expected to take much longer and may never occur. Thus, it is possible that the grounding sites may stabilize to some alternate state that is dissimilar from what was previously present before impact.
In conclusion, ship groundings on coral reefs cause not only serious damage to the reef builders, but also result in a loss of habitat for other organisms, consequently leading to barren areas. They often flatten the topography, leaving a source of sand and rubble that may move over the area and inhibit benthic organism settlement, growth, and survival. This study has shown that corals are able to recruit to damaged areas but that slow growth rates and high mortality rates may keep these areas in a perpetual cycle of settlement and mortality with little or extremely slow growth to larger size classes, thus inhibiting recovery. While the exact processes underlying differences in recruitment, growth, and mortality, both among sites in this study and in comparison to other areas of Florida, are unknown, we believe that the loss of topography at the grounding sites will negatively impact coral recovery. Therefore, methods of restoration compensating for increased sediment mobility and flattened relief are recommended to enhance natural recovery potential of grounding sites.
Acknowledgments
We thank Nova Southeastern University graduate students for their assistance with field work and Brian Walker for GIS assistance. The Hillsboro Inlet District and Florida Department of Environmental Protection provided funding for this project.
References
Arnold, S.N., R.S. Steneck & P.J. Mumby. 2010. Running the gauntlet: inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 414: 91-105. [ Links ]
Aronson, R.B. & D.W. Swanson. 1997. Video surveys of coral reefs: uniand multivariate applications. Proc. 8th Int. Coral Reef Symp., Panama 2: 1441-1446. [ Links ]
Babcock, R.C. 1985. Growth and mortality in juvenile corals (Goniastrea, Platygyra, and Acropora): the first year. Proc. 5th Int. Coral Reef Congr., Tahiti 4: 355-360. [ Links ]
Bak, R.P.M. & M.S. Engel. 1979. Distribution, abundance and survival of juvenile hermatypic corals (Scleractinia) and the importance of early life history strategies in the parent coral community. Mar. Biol. 54: 341-352. [ Links ]
Banks, K.W., B.M. Riegl, V.P. Richards, B.K. Walker, K.P. Helmle, L.K.B. Jordan, J. Phipps, M.S. Shivji, R.E. Spieler & R.E. Dodge. 2008. The reef tract of continental southeast Florida (Miami-Dade, Broward and Palm Beach Counties, USA), p. 175-220. In B.M. Riegl and R.E. Dodge (eds). Coral Reefs of the USA. Springer, London, England. [ Links ]
Birkeland, C. 1977. The importance of rate of biomass accumulation in early successional stages of benthic communities to the survival of coral recruits. Proc. 3rd Int. Coral Reef Symp., Miami 1: 15-21. [ Links ]
Cook, C.B., R.E. Dodge, & S.R. Smith. 1994. Fifty years of impacts on coral reefs in Bermuda. p. 160-166. In R.N. Ginsburg (compiler). Proc. Colloquium on Global Aspects of Coral Reefs: Health, Hazards, and History. University of Miami, Miami, Florida, USA. [ Links ]
Edmunds, P.J. 2000. Patterns in the distribution of juvenile corals and coral reef community structure in St. John, US Virgin Islands. Mar. Ecol. Prog. Ser. 202: 113-124. [ Links ]
Edwards, A.J. & E.D. Gomez. 2007. Reef restoration concepts and guidelines: making sensible management choices in the face of uncertainty. Coral Reef Targeted Research & Capacity Building for Management Programme, St. Lucia, Australia. [ Links ]
Gittings, S.R., T.J. Bright & B.S. Holland. 1990. Five years of coral recovery following a freighter grounding in the Florida Keys. p. 89-105. In W.A. Jaap (ed). Diving for Science: Proceedings of the American Academy of Underwater Sciences 10th Annual Scientific Diving Symposium. American Academy of Underwater Sciences, Costa Mesa, California, USA. [ Links ]
Harriott, V.J. 1999. Coral recruitment at a high latitude Pacific site: a comparison with Atlantic reefs. Bull. Mar. Sci. 65: 881-891. [ Links ]
Harriott, V.J. & S.A. Banks. 1995. Recruitment of scleractinian corals in the Solitary Islands Marine Reserve, a high latitude coral-dominated community in eastern Australia. Mar. Ecol. Prog. Ser. 123: 155-161. [ Links ]
Hatcher, B.G. 1984. A maritime accident provides evidence for alternate stable states in benthic communities on coral reefs. Coral Reefs 3: 199-204. [ Links ]
Hudson Marine Management Services. 2004a. M/V Eastwind Grounding Site Ft. Lauderdale, Florida Injury Assessment. Thomas Miller Inc., Miami, Florida, USA. [ Links ]
Hudson Marine Management Services. 2004b. M/V Federal Pescadores Site Ft. Lauderdale, Florida Injury Assessment Report. Vandeventer Black LLP, Norfolk, Virginia, USA. [ Links ]
Hughes, T.P. & J.B.C. Jackson. 1985. Population dynamics and life histories of foliaceous corals. Ecol. Monogr. 55: 141-166. [ Links ]
Hughes, T. P. & J. Tanner. 2000. Recruitment failure, life histories, and long-term decline of Caribbean corals. Ecology 81: 2250-2263. [ Links ]
Hughes, T.P, A.H. Baird, E.A. Dinsdale, N.A. Moltschaniwsky, M.S. Pratchet, J.E. Tanner & B.L. Willis. 2000. Supply-side ecology works both ways: the link between benthic adults, fecundity, and larval recruits. Ecology 81: 2241-2249. [ Links ]
Hughes, T.P., A.H. Baird, E.A. Dinsdale, V.J. Harriott, N.A. Moltschaniwsky, Pratchet, J.E. Tanner & B.L. Willis. 2002. Detecting regional variation using meta-analysis and large-scale sampling: latitudinal patterns in recruitment. Ecology 83: 436-451. [ Links ]
Jaap, W.C., J.H. Hudson R.E. Dodge, D. Gilliam & R. Shaul. 2006. Coral reef restoration with case studies from Florida. p. 478-514. In I.M. Cote & J.D. Reynolds (eds). Coral Reef Conservation. Cambridge University, Cambridge, England. [ Links ]
Kuffner, I.B., L.J. Walters, M.A. Becerro, V.J. Paul, R. Ritson-Williams & K.S. Beach. 2006. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 323: 107-117. [ Links ]
Lirman, D. & M.W. Miller. 2003. Modeling and monitoring tools to assess recovery status and convergence rates between restored and undisturbed coral reef habitats. Restor. Ecol. 11: 448-456. [ Links ]
Lirman, D. & D. Manzello. 2007. Patterns of resistance and resilience of the stress-tolerant coral Siderastrea radians (Pallas) to sub-optimal salinity and sediment burial. J. Exp. Mar. Biol. Ecol. 369: 72-77. [ Links ]
Macintyre, I.G. & O.H. Pilkey. 1969. Tropical reef corals: tolerance of low temperatures on the North Carolina continental shelf. Science 166: 374-375. [ Links ]
Maida, M., P.W. Sammarco & J.C. Coll. 2001. Effects of soft corals on scleractinian coral recruitment. 2: Allelopathy, spat survivorship and reef community structure. Mar. Ecol. 22: 397-414. [ Links ]
Marine Resources Inc. 2005. Proposed Restoration Plan for Post-Grounding Activities: M/V Eastwind Grounding Site Offshore Broward County, Florida. Houck, Hamilton, & Anderson, P.A., Miami, Florida, USA. [ Links ]
Miller, M.W., E. Weil & A.M. Szmant. 2000. Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA. Coral Reefs 19: 115-123. [ Links ]
Moyer, R.P., B. Riegl, K. Banks & R.E. Dodge. 2003. Spatial patterns and ecology of benthic communities on a high-latitude South Florida (Broward County, USA) reef system. Coral Reefs 22: 447-464. [ Links ]
Muthiga, N.A. & A.M. Szmant. 1987. The effects of salinity stress on the rates of aerobic respiration and photosynthesis in the hermatypic coral Siderastrea siderea. Biol. Bull. 173: 539-551. [ Links ]
Rogers, C.S, H.C. Fitz III, M. Gilnack, J. Beets & J. Hardin. 1984. Scleractinian coral recruitment patterns at Salt River Submarine Canyon, St. Croix, U.S. Virgin Islands. Coral Reefs 3: 69-76. [ Links ]
Rogers, C.S. & V. Garrison. 2001. Ten years after the crime: lasting effects of damage from a cruise ship anchor on a coral reef in St. John, U.S. Virgin Islands. Bull. Mar. Sci. 69: 793-803. [ Links ]
Rylaarsdam, K.W. 1983. Life histories and abundance patterns of colonial corals on Jamaican reefs. Mar. Ecol. Prog. Ser. 13: 249-260. [ Links ]
Smith, S.R. 1997. Patterns of coral settlement, recruitment and juvenile mortality with depth at Conch Reef, Florida. Proc. 8th Int. Coral Reef Symp., Panama 2: 1197-1202. [ Links ]
St. Gelais, A.T. 2010. Reproductive ecology of Siderastrea siderea: histological analysis of gametogenesis, spawning, and latitudinal fecundity variation. Master’s Thesis, Nova Southeastern University, Dania Beach, Florida, USA. [ Links ]
Thanner, S.E., T.L. McIntosh & S.M. Blair. 2006. Development of benthic and fish assemblages on artificial reef materials compared to adjacent natural reef assemblages in Miami-Dade County, Florida. Bull. Mar. Sci. 78: 57-70. [ Links ]
Van Moorsel, G.W.N.M. 1988. Early maximum growth of stony corals (Scleractinia) after settlement on artificial substrata on a Caribbean reef. Mar. Ecol. Prog. Ser. 50: 127-135. [ Links ]
Vermeij, M.J.A. 2006. Early life-history dynamics of Caribbean coral species on artificial substratum: the importance of competition, growth and variation in life-history strategy. Coral Reefs 25: 59-71. [ Links ]
*Correspondencia:
Alison L. Moulding: National Coral Reef Institute, Nova Southeastern University, Oceanographic Center, 8000 North Ocean Dr., Dania Beach, FL 33161 USA; moulding@nova.edu
Vladimir N. Kosmynin: Florida Department of Environmental Protection, Bureau of Beaches and Coastal Systems, 3900 Commonwealth Boulevard, Mail Station 300, Tallahassee, Florida 32399 USA; Vladimir.Kosmynin@dep.state.fl.us
David S. Gilliam: National Coral Reef Institute, Nova Southeastern University, Oceanographic Center, 8000 North Ocean Dr., Dania Beach, FL 33161 USA; gilliam@nova.edu
1. National Coral Reef Institute, Nova Southeastern University, Oceanographic Center, 8000 North Ocean Dr., Dania Beach, FL 33161 USA; moulding@nova.edu, gilliam@nova.edu
2. Florida Department of Environmental Protection, Bureau of Beaches and Coastal Systems, 3900 Commonwealth Boulevard, Mail Station 300, Tallahassee, Florida 32399 USA; Vladimir.Kosmynin@dep.state.fl.us
Received 12-VII-2011. Corrected 22-XI-2011. Accepted 20-XII-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}