










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.4 San José Dec. 2012
Chemosensory age discrimination in the snake Boa constrictor (Serpentes: Boidae)
*Dirección para correspondencia
Abstract
Many snakes are able to use their chemosensory system to detect scent of conspecifics, which is important in many social contexts. Age discrimination based on chemical cues may be especially important to ensure access to sexually mature potential partners. In this study, we used 24 individual Boa constrictor snakes (12 adults mature and 12 non-mature individuals) that had been captured in different areas of Ecuador, and were maintained in captivity at the Vivarium of Quito. We used tongue-flick experiments to examine whether these snakes were able to discriminate between scents from mature and non-mature individuals. Results showed that B. constrictor snakes used chemical cues to recognize conspecifics and that the scent of individuals of different ages elicited chemosensory responses of different magnitudes. The scents from adult conspecifics elicited the quickest and highest chemosensory responses (i.e., short latency times and high tongue-flick rates), although we did not find differential responses to scent of males and females. The magnitude of the responses was lower to scent of sub adult individuals, and then even lower to scent of juvenile snakes, but in all cases the scent of snakes was discriminated from a blank control. We discuss the potential chemical mechanisms that may allow age recognition and its implications for social and sexual behavior of this snake species.
Key words: age recognition, Boa constrictor, chemoreception, Ecuador, snakes.
Resumen
Muchas serpientes son capaces de usar su sistema quimiosensorial para detectar el olor de individuos coespecíficos, lo que es importante en muchos contextos sociales. La discriminación de la edad basada en señales químicas puede ser especialmente importante para asegurar el acceso a parejas potenciales que sean sexualmente maduras. En este estudio, usamos 24 individuos de una especie de boa (Boa constrictor) (12 individuos adultos y 12 inmaduros) que habían sido capturados en diferentes partes de Ecuador y eran mantenidos en cautividad el Vivarium de Quito. Usamos experimentos de protusiones linguales para examinar si esta serpiente es capaz de discriminar entre el olor de individuos maduros y no maduros. Los resultados mostraron que B. constrictor usa señales químicas para reconocer co-específicos y que el olor de individuos de distinta edad provoca respuestas quimiosensoriales de diferente magnitud. El olor de individuos adultos provocó las respuestas más rápidas y elevadas (esto es, tiempos de latencia más cortos y tasas más altas de protusiones linguales), aunque no encontramos diferencias en las respuestas a olores de machos y hembras. La magnitud de las respuestas fue más baja a olores de sub adultos, e incluso más baja a olor de juveniles, pero en todos los casos el olor de una serpiente era discriminado de un control no oloroso. Discutimos los posibles mecanismos químicos que pueden permitir esta discriminación de la edad y sus implicaciones para el comportamiento social y sexual de esta serpiente.
Palabras clave: Boa constrictor, Ecuador, quimio recepción, reconocimiento de la edad, serpientes.
Most snakes have the chemosensory ability to discriminate substrate deposited scent trails of conspecifics, which is useful within social, foraging or anti predatory contexts (Ford & Low 1984, Ford 1986, Greene et al. 2001, LeMaster et al. 2001, reviewed in Mason 1992, Mason & Parker 2010). Pheromones in scent marks may inform on the species identity, sex, sexual attractiveness or mating status of the signaler individual (Ford & Schofield 1984, LeMaster & Mason 2001, O’Donnell et al. 2004). The ability of snakes to locate mates based on pheromonal cues in the environment is very important because it can increase their reproductive success (Shine et al. 2005).
Among the many potential chemosensory abilities, age discrimination based on chemical cues may be determinant in several circumstances. For example during the mating season; to ensure access to potential partners, animals should be able to discriminate sexually mature from non mature individuals (O’Donnell et al. 2004). Also, in many cases females should prefer to mate with older males because viability selection leads to older males of higher genotypic quality than younger males (Brooks & Kemp 2001, López et al. 2003). Similarly, males may prefer older/larger females because these may produce more offspring (Gregory 1977). In snakes, mature and young individuals differ in body size, but in addition to this simple visual cue, chemical cues signaling age or sexual maturity could be important to identify potential mates (Shine & Mason 2001). For example, male garter snakes can assess the body size (i.e., age) and body condition of females based solely on female pheromone cues alone (Shine et al. 2003). This chemosensory ability may be especially useful to locate hidden potential partners in snake species with secretive habits or that live in complex environments.
There are many studies of chemical communication in European and North American snakes (e.g., Andren 1982, Mason 1993, LeMaster & Mason 2001, Mason & Parker 2010). However, the ecology and biology of Neotropical snakes is poorly known. Especially, there is little knowledge on the use of chemical signals by most snake species from South America. In Epicrates cenchria, both sexes can detect and discriminate chemical cues of conspecifcs from other boid species (Briguera et al. 1994, 1998). Other study showed that male and female Boa constrictor occidentalis detect conspecifics odors and discriminate between sexes based on chemical cues from the skin and the cloacal glands (Chiaraviglio & Briguera 2001). These chemosensory abilities may help males to follow scent trails of conspecific females (Briguera et al. 1997, Cervantes & Chiaraviglio 1999). Also, the South American colubrid Waglerophis merremii can discriminate sex of conspecifics (Chiaraviglio & Gutiérrez 1994).
In this study, we examined whether a boa snake species (Boa constrictor) from Ecuador is able to discriminate between scents from mature and non-mature individuals We used tongue flicks experiments to measure the chemosensory responses of snakes (latency time to respond and number of tongue flicks) to scents of conspecifics of different ages (adults, sub adults and juveniles). We hypothesized that the abundance, composition or proportion of chemical compounds in scent should differ with the age of snakes (e.g., LeMaster & Mason 2002), and this would allow chemosensory age recognition.
Materials and Methods
Study species: We studied a snake species, B. constrictor (L., 1758), member of the family Boidae. This is a boa species mainly found in Central and South America and some Antillean islands. These snakes inhabit a wide variety of environmental conditions, from tropical rainforests to arid areas. They have a highly variable color pattern and may grow to become quite large (maximum 4m). Small individuals may climb into trees and shrubs to forage, but they become mostly terrestrial as they become older and heavier (Mehrtens 1987, Valencia et al. 2008). Boas give birth to live young of approx. 0.5m long and sexual maturity occurs at a length of 1.5-2m (Greene 1983).
In this study, we used 24 individual B. constrictor snakes that had been captured in different areas of Ecuador, and were maintained in captivity at the Vivarium of Quito, Fundación Herpetológica Gustavo Orcés (Av. Amazonas N.º 3008 y Rumipamba, Quito, Ecuador). All individuals were maintained separately in their own terraria, with the adequate temperature (24-26°C) and humidity conditions (58-78%). Snakes were fed weekly chicken or mice (the type and number of prey items depended on the size of each snake). We considered adult mature individuals (n=12, six males and six females) to those with a body length of at least >1.5m (Greene 1983), although most individuals were much larger. Non-mature individuals (n=12, seven males and five females) included snakes of two age classes: sub adults (between 1-1.5m of body length, n=7) and juveniles (body length <1m, n=5).
Scent recognition experiments: Snakes have been shown to react to a variety of chemical stimuli with increased and differential rates of tongue extrusions. Thus, tongue-flick (TF) rate can be used as a quantitative bioassay of detection and discrimination of chemical cues (Cooper & Burghardt 1990, Schulterbrandt et al. 2008). To test for differential responses to scents we made comparisons of TF rate by male and female B. constrictor snakes of different ages in response to chemical stimuli arising from cotton swab applicators impregnated with scents of conspecific adult males or females, sub adults, juveniles, or with deionized water (odorless control). Water was used to gauge baseline TF rates in the experimental situation (Cooper & Burghardt 1990). Also, as a control of responses to scent of a heterospecific snake, we used scent from adult males and females (n=7) of another snake species from Ecuador, the Amazon tree boa (Corallus hortulanus) (Mehrtens 1987, Valencia et al. 2008). These snakes were also maintained in the Vivarium of Quito. We obtained snake scents from the cloacal area and the skin surrounding it of males or females, because these body parts produce chemicals that are deposited on substrate trails, and can be explored by other snakes (Mason & Parker 2010). Therefore, after first dipping the cotton tip (1cm) of a wooden applicator attached to a long stick (50cm) in deionized water, we rolled the tip over the cloaca and the skin area 2cm around the cloacal area (of one species, age and sex per applicator). A new swab was used in each trial.
Trials were conducted during October-November 2008. Prior to the tests, snakes were gently taken from their terraria and kept individually in clean plastic cages in an experimental room, where snakes were left for 30min for acclimatization before the trials. Because temperature may strongly influence tongueflick rates of snakes (Stevenson et al. 1985), snakes were maintained at their optimal body temperatures (around 25°C) inside each terrarium and in the experimental room at least one hour before and during the experiments. Every B. constrictor snake (n=24) was exposed to each scent stimulus and order of presentation was randomized. One trial per each animal was conducted each day.
To begin a trial, the experimenter slowly approached the cage and slowly moved the cotton swab to a position 1cm anterior to the snake`s snout. Snakes usually did not flee from the swab, but explored it repeatedly by tongueflicking or ignored it after the firsts TFs. The numbers of TFs directed to the swab were recorded for 60s beginning with the first TF. Latency to the first TF was computed as the number of seconds elapsed between presentations of the cotton swab to the first TF directed to the swab.
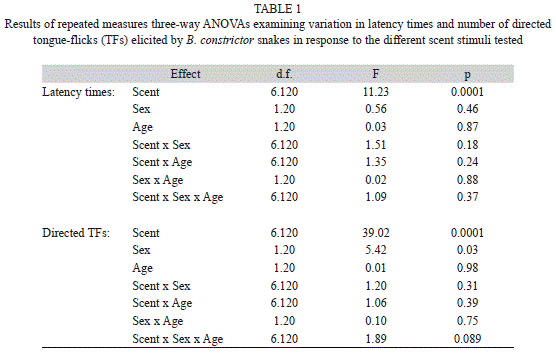
To examine differences in latency times and number of directed TFs (both log-transformed) among scent stimuli presented, we used three-way repeated measures analyses of variance (ANOVAs) with scent stimuli as a within factor, and with the sex (male vs. female) and age (adult vs. non-mature) of the responding snake as between factors. Pairwise post-hoc comparisons were planned using Tukey’s honestly significant difference (HSD) tests (Sokal & Rohlf 1995).
Results
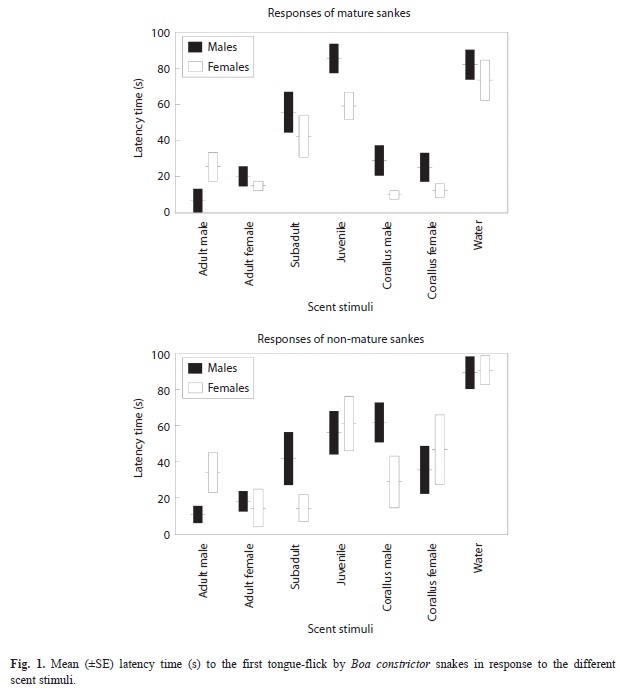
Latency time: There were significant differences in latencies to the first TF between scent stimuli tested, but B. constrictor snakes of different ages and sexes responded in a similar way to the different stimuli (i.e., the interactions were not significant) (Table 1, Fig. 1). Latencies to water were significantly longer than to any other stimuli (Tukey’s tests, p<0.02 in all cases). Latencies to scent of adult male and female B. constrictor were not significantly different (p=0.98), and both were significantly shorter than to scent from sub adult and juvenile B. constrictor snakes (p<0.01 in all cases), which did not significantly differ (p=0.86). Latencies to scent of male and female C. hortulanus did not significantly differ from latencies to scents of B. constrictor snakes of any sex or age (p>0.25 in all cases) (Fig. 1).
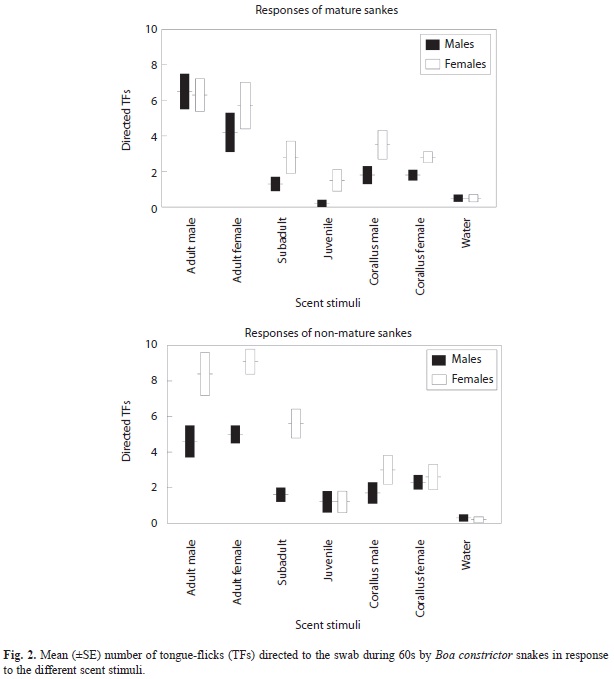
Tongue flicks directed to the swab: The numbers of TFs directed to the swab by B. constrictor snakes were significantly different depending on the scent stimuli, and female snakes had a significantly higher overall TF rate than males. However, snakes of all sexes and ages responded in a similar way to the different stimuli (i.e., the interactions were not significant) (Table 1, Fig. 2).
Tongue flick rate to water was significantly lower than to any other stimuli (Tukey’s tests, p<0.001 in all cases). Responses to adult male and female B. constrictor were not significantly different (p=0.99), both were significantly higher than to scent from subadult and juvenile B. constrictor snakes (p=0.00012 in all cases), and TF rates to scent of sub adults were significantly higher than to scent of juveniles (p=0.02). Responses to scent of adult male and female C. hortulanus were not significantly different (p=0.99). Both were significantly greater than to scent of juveniles (p<0.05), did not differ from responses to sub adults (p=0.99) and were significantly lower than responses to adult male and female B. constrictor (p=0.00014 in all cases) (Fig. 2).
Discussion
Our results showed that B. constrictor snakes used chemical cues to recognize conspecifics and that the scent of individuals of different ages elicited chemosensory responses of different magnitude. The chemosensory tests first showed that individuals of any sex or age responded quicker with tongue-flicks and had higher responses to cotton swabs impregnated with any snakes’ scents than to cotton swabs with water alone. This confirms previous findings with other subspecies of this boa snake (Chiaraviglio & Briguera 2001) that indicate that this species has well developed the chemosensory system and can distinguish the odor of a snake from a blank control (Halpern 1992, Cooper & Burghardt 1990).
Further, results of this study showed that the scents from adult conspecifics elicited the quickest and highest chemosensory responses although we did not find differential responses to scent of males and females, in contrast to previous studies where male B. constrictor show higher TF responses to female scent than to male scent (Chiaraviglio & Briguera 2001). Something similar occurs in other Boidae species, E. cenchria (Briguera et al. 1994, 1997). The apparent lack of sex discrimination in our study might be explained because chemosensory responses of snakes to conspecific scents may depend on their reproductive state (Chiaraviglio & Gutiérrez 1994).
In our study, the magnitude of the chemosensory responses was lower to scent of sub adult individuals, and even lower to scent of juvenile snakes. Nevertheless, as indicated above, all of these scents were detected by snakes. Interestingly, the scent of adults of a heterospecific snake was quickly detected but later elicited low TF responses that did not differ from responses to non-mature conspecifics. The clear differences in TF rates to scent of adult B. constrictor and to scent of adult C. hortulanus indicated that there is species recognition. These results could suggest that although all these scents could be detected, after being identified, some of them were not further considered for more detailed chemosensory exploration, resulting in lower TF rates.
These differences in the magnitude of the responses are probably related to the different importance that finding individuals of different ages may have in social and sexual behavior of this snake. For example, in male Boiga irregularis snakes, adult female skin lipid pheromones induce courtship behavior, while adult male skin lipid pheromones induce combat behaviors (Greene & Mason 1998, 2000). Nonmature individuals are not involved in sexual behavior and, thus, finding their scent should not elicit further exploratory behavior. Age recognition based on chemical cues also occur in other animals, such as lizards and mammals (López et al. 2003, Osada et al. 2003), probably because it may have important reproductive consequences. However, to understand the role of chemical cues in social behavior of B. constrictor snakes we require further experiments.
Mature and non-mature B. constrictor snakes responded in a similar way to the different scent stimuli, with high responses to scent of adult individuals and low responses to scent of juveniles. Although when finding a scent the “motivation” and posterior behavioral responses should vary with the age of the responding snake, it is likely that the same chemical stimuli induced similar tongue-flick responses in the vomeronasal system of any conspecific snake (Halpern 1992). Nevertheless, female snakes had higher overall TF rates to most stimuli than males, which could be due to sexual differences in the chemosensory system or tongue-flick behavior. However, further studies are needed to understand the inter-individual variation in these responses.
The chemosensory age discrimination may simply respond to differences in the amount of chemical secretions that individuals of different age produce, with juvenile snakes producing the lowest amounts of secretions. The production of secretions could be under control of gonadal steroid hormones that would be only active in sexually mature individuals (Mason 1993, Mason & Parker 2010). This would explain the decreasing latency times and increasing TF rates when age of the donor of the scent increased. However, it is also likely that there were qualitative differences in the compounds found in scent. For example, in female garter snakes, Thamnophis sirtalis, the methyl ketone pheromone blend found in the scent of the skin becomes dominated by the longest chain, unsaturated methyl ketones with increasing body length (LeMaster & Mason 2002). These differences allow males to use scent of females to discriminate the size and body condition of different individual females (Shine et al. 2003).
In summary, in this study we show that B. constrictor snakes are able to use chemical cues of conspecifics in intraspecific recognition and age discrimination. Further studies are needed to clarify how these chemosensory abilities are used in social and sexual behavior of this snake, and to know the specific chemical compounds involved in the responses. Also, more studies with other snake species are necessary to know the extent of generalization of chemical senses in snakes, and to have a better understanding of the functional and evolutionary role of pheromones in reptiles.
Acknowledgments
We thank three anonymous reviewers for helpful comments, and K. Garzón and M.E. Barragán, from the Vivarium de Quito, Fundación Herpetológica Gustavo Orcés, for access and facilities to work with the snakes. Financial support was provided by the projects MCI-CGL2008-02119/BOS and MICIIN-CGL2011-24150/BOS, and by an “El Ventorrillo” CSIC grant to MG. The experiments enforced all the present Ecuatorian laws and were performed under license from the Ministerio del Ambiente, Ecuador.
References
Andren, C. 1982. The role of the vomeronasal organs in the reproductive behavior of the adder Vipera berus. Copeia 1982: 148-157. [ Links ]
Briguera, V., M. Chiaraviglio & M. Gutiérrez. 1994. Comunicación química en la boa arco iris Epicrates cenchria alvarezi (Serpentes, Boidae). Cuad. Herp. 8: 173-176. [ Links ]
Briguera, V., M. Chiaraviglio & M. Gutiérrez. 1997. Experiencias de seguimiento de rastro por los machos de boa arco iris Epicrates cenchria alvarezii (Serpentes-Boidae) mediado por mensajes químicos. Cuad. Herp. 11: 7-12. [ Links ]
Briguera, V., M. Chiaraviglio & M. Gutiérrez. 1998. Observaciones sobre la discriminación de señales químicas heteroespecíficas de reconocimiento en Epicrates cenchria alvarezii, Abalos, Báez y Nader (Serpentes: Boidae). Gayana Zool. 62: 215-220. [ Links ]
Brooks, R. & D.J. Kemp. 2001. Can older males deliver the good genes? Trends Ecol. Evol. 6: 308-313. [ Links ]
Cervantes, R.S. & M. Chiaraviglio. 1999. Actividad de rastreo en Boa constrictor occidentalis (Serpentes: Boidae), un mecanismo de localización de la especie. Cuad. Herp. 13: 3-10. [ Links ]
Chiaraviglio, M. & V. Briguera. 2001. Participation of chemical cues in conspecific detection and sexual discrimination in Boa constrictor occidentalis (Serpentes: Boidae). Gayana 65: 5-10. [ Links ]
Chiaraviglio, M. & M. Gutiérrez. 1994. Reconocimiento de olor feromonal conespecífico en Waglerophis merremii (Serpentes, Colubridae), influencia estacional y hormonal. Cuad. Herp. 8: 126-130. [ Links ]
Cooper, W.E. & G.M. Burghardt. 1990. A comparative analysis of scoring methods for chemical discrimination of prey by squamate reptiles. J. Chem. Ecol. 16: 45-65. [ Links ]
Ford, N.B. 1986. The role of pheromone trails in the sociobiology of snakes, p. 261-278. In D. Duvall, D. Muller-Schwarze & R.M. Silverstein (eds.). Chemical signals in Vertebrates IV. Ecology, evolution and comparative biology. Plenum, New York, USA. [ Links ]
Ford, N.B. & J.R. Low. 1984. Sex pheromone source location by garter snakes: a mechanism for detection of direction in nonvolatile trails. J. Chem. Ecol. 10: 1193-1199. [ Links ]
Ford, N.B. & C.W. Schofield. 1984. Species specificity of pheromone trails in the plain garter snakes, Thamnophis radix. Herpetologica 40: 51-55. [ Links ]
Greene, H.W. 1983. Boa constrictor (boa, béquer, boa constrictor), p. 380-382. In D.H. Janzen (ed.). Costa Rican natural history. University of Chicago, Chicago, USA. [ Links ]
Greene, M.J. & R.T. Mason. 1998. Chemically mediated sexual behavior of the brown tree snake, Boiga irregularis. EcoSci. 5: 405-409. [ Links ]
Greene, M.J. & R.T. Mason. 2000. Courtship, mating, and male combat of the brown tree snake, Boiga irregularis. Herpetologica 56: 166-175. [ Links ]
Greene, M.J., S.L. Stark & R.T. Mason. 2001. Pheromone trailing behavior of the brown tree snake, Boiga irregularis. J. Chem. Ecol. 27: 2193-2201. [ Links ]
Gregory, P.T. 1977. Life-history parameters of the redsided garter snake (Thamnophis sirtalis parietalis) in an extreme environment, the Interlake region of Manitoba. Nat. Mus. Can. Pub. Zool. 13: 1-44. [ Links ]
Halpern, M. 1992. Nasal chemical senses in reptiles: structure and function, p. 423-523. In C. Gans & D. Crews (eds.). Biology of the Reptilia, Vol. 18. Brain, hormones, and behavior. University of Chicago, Chicago, USA. [ Links ]
LeMaster, M.P. & R.T. Mason. 2001. Evidence for a female sex pheromone mediating male trailing behavior in the red–sided garter snake, Thamnophis sirtalis parietalis. Chemoecology 11: 49-152. [ Links ]
LeMaster, M.P. & R.T. Mason. 2002. Variation in a female sexual attractiveness pheromone controls male mate choice in garter snakes. J. Chem. Ecol. 28: 1269-1285. [ Links ]
LeMaster, M.P., I.T. Moore & R.T. Mason. 2001. Conspecific trailing behavior of red–sided garter snakes, Thamnophis sirtalis parietalis, in the natural environment. Anim. Behav. 61: 827-833. [ Links ]
López, P., P. Aragón & J. Martín. 2003. Responses of female lizards, Lacerta monticola, to males’ chemical cues reflect their mating preference for older males. Behav. Ecol. Sociobiol. 55: 73-79. [ Links ]
Mason, R.T. 1992. Reptilian pheromones, p. 114-228. In C. Gans & D. Crews (eds.). Biology of the Reptilia, Vol. 18. Brain, hormones, and behavior. University of Chicago, Chicago, USA. [ Links ]
Mason, R.T. 1993. Chemical ecology of the red-sided garter snake, Thamnophis sirtalis parietalis. Brain Behav. Evol. 41: 261-268. [ Links ]
Mason, R.T. & M.R. Parker. 2010. Social behavior and pheromonal communication in reptiles. J. Comp. Physiol. A 196: 729-749. [ Links ]
Mehrtens, J.M. 1987. Living snakes of the world in color. Sterling, New York, USA. [ Links ]
O’Donnell, R.P., N.B. Ford, R. Shine & R.T. Mason. 2004. Male red-sided garter snakes, Thamnophis sirtalis parietalis, determine female mating status from pheromone trails. Anim. Behav. 68: 677-683. [ Links ]
Osada, K., K. Yamazaki, M. Curran, J. Bard, B.P.C. Smith, G.K. Beauchamp. 2003. The scent of age. Proc. R. Soc. Lond. B 270: 929-933. [ Links ]
Schulterbrandt, T.G., J. Kubie, H. von Gizycki, I. Zuri & M. Halpern. 2008. Patterns of tongue-flicking by garter snakes (Thamnophis sirtalis) during presentation of chemicals under varying conditions, p. 345-356. In J.L. Hurst, R.J. Beynon, S.C. Roberts & T.D. Wyatt (eds.). Chemical signals in Vertebrates. Vol. 11. Springer, New York, USA. [ Links ]
Shine, R. & R.T. Mason. 2001. Courting male garter snakes (Thamnophis sirtalis parietalis) use multiple cues to identify potential mates. Behav. Ecol. Sociobiol. 49: 465-473. [ Links ]
Shine, R., B. Phillips, H. Waye, M.P. LeMaster & R.T. Mason. 2003. Chemosensory cues allow courting male garter snakes to assess body length and body condition of potential mates. Behav. Ecol. Sociobiol. 54: 162-166. [ Links ]
Shine, R., R.P. O’Donnell, T. Langkilde, M.D. Wall & R.T. Mason. 2005. Snakes in search of sex: the relation between mate-locating ability and mating success in male garter snakes. Anim. Behav. 69: 1251-1258. [ Links ]
Sokal, R.R. & F.J. Rohlf. 1995. Biometry. Freeman, New York, USA. [ Links ]
Stevenson, R.D., C.R. Peterson & J.S. Tsuji. 1985. The thermal dependence of locomotion, tongue flicking, digestion, and oxygen consumption in the wandering water snake. Physiol. Zool. 58: 46-57. [ Links ]
Valencia, J.H., E. Toral, M. Morales, R. Betancourt & A. Barahona. 2008. Guía de campo de reptiles de Ecuador. Fundación Herpetológica Gustavo Orcés, Simbioe, Quito, Ecuador. [ Links ]
*Correspondencia: Marianne Gabirot: Departamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales, CSIC, José Gutiérrez Abascal 2, 28006 Madrid, Spain; marianne.gabirot@cefe.cnrs.fr . Department Ecologie Comportementale, (U.M.R. 5175), CEFE-CNRS, 1919 Route de Mende, F34293 Montpellier, Cedex 5, France.
Pablo Picerno: Fundacion Herpetológica Gustavo Orces, Vivarium de Quito, Av. Amazonas 3008 y Rumipamba, Quito, Ecuador; pablopicerno@yahoo.com
Jorge Valencia: Fundacion Herpetológica Gustavo Orces, Vivarium de Quito, Av. Amazonas 3008 y Rumipamba, Quito, Ecuador; jvalencia@fhgo.org.ec . Pontificia Universidad Católica del Ecuador, Escuela de Biología, Museo de Zoología. Avenida 12 de Octubre y Roca, Apartado 17-01-2184. Quito, Ecuador.
Pilar Lopez: Departamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales, CSIC, José Gutiérrez Abascal 2, 28006 Madrid, Spain; pilar.lopez@mncn.csic.es
José Martin: Departamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales, CSIC, José Gutiérrez Abascal 2, 28006 Madrid, Spain; jose.nartin@mncn.csic.es
1. Departamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales, CSIC, José Gutiérrez Abascal 2, 28006 Madrid, Spain; jose.nartin@mncn.csic.es, pilar.lopez@mncn.csic.es
2. Department Ecologie Comportementale, (U.M.R. 5175), CEFE-CNRS, 1919 Route de Mende, F34293 Montpellier, Cedex 5, France; marianne.gabirot@cefe.cnrs.fr
3. Fundacion Herpetológica Gustavo Orces, Vivarium de Quito, Av. Amazonas 3008 y Rumipamba, Quito, Ecuador; pablopicerno@yahoo.com
4. Pontificia Universidad Católica del Ecuador, Escuela de Biología, Museo de Zoología. Avenida 12 de Octubre y Roca, Apartado 17-01-2184. Quito, Ecuador; jvalencia@fhgo.org.ec
Received 01-XI-2011.Corrected 04-V-2012.Accepted 05-VI-2012.


{kind=link}
{kind=link}
{kind=link}