










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.4 San José Dec. 2012
Recuperación de tres coberturas forestales de altura media en Costa Rica: análisis de los oligoquetos, el mantillo y suelo
Recovery of three tropical forest covers from mid-elevation sites in Costa Rica: oligochaetes, litter and soil analysis
Recovery of three tropical forest covers from mid-elevation sites in Costa Rica: oligochaetes, litter and soil analysis
*Dirección para correspondencia
Abstract
In Costa Rica, the region of Río Macho is a highly fragmented landscape with imminent risk of landslides. This area, which provides important environmental services, has been partially recovered to its original forest through intentional reforestation with exotic species or natural regeneration after abandonment. The aim of this study was to evaluate the bioindicator potential of oligochaete presence as well as some litter and soil characteristics. The ecosystem recovery of the two common restoration modes was measured within three different forest covers. For this, some substrate characteristics were analyzed and compared in a 50 years old secondary forest, a 13 years tacotal, and a 35 years cypress (Cupressus lusitanica) plantation. The three sites studied differed in density, biomass and average mass of oligochaetes, and in some litter (depth, nitrogen, phosphorus and C/N ratio of litter), and soil variables (soil water content (CA), pH, phosphorus, cation exchange capacity, and magnesium). The forest registered the lowest density of earthworms and soil pH, and the highest soil CA and phosphorus. CA was inversely related to the oligochaete density across sites. Besides, there were positive correlations between C/N and C/P ratios from the litter and soil pH, and inverse correlations of litter depth, litter N and P concentrations with soil P. Discriminant Analysis (AD) performed with all soil and litter variables, produced a sharp classification of the three forest cover types. AD suggests that site differences were mostly determined by soil CA and litter nitrogen concentration. Considering all the evaluated parameters, our results suggest in the first place, that oligochaetes are sensitive to changes in some soil and litter characteristics. Secondly, aside from the striking oligochaete differences between the old secondary forest and the other two sites, some soil and litter traits resulted good indicators of the present recovery of the three forest covers. In addition, comparing soil nutrients content (organic carbon, nitrogen, calcium, potassium and sulfur) among the three sites, our findings indicate that the cypress plantation had reached soil nutrient conditions similar to the old secondary forest, presumably by the accumulation of nutrients, as a result of low nutrient recirculation. In conclusion, ecosystem level studies throughout simple evaluation criteria (soils, oligochaetes and ground litter) can be used as rapid indicators of the state of some of the many and complex forest ecosystem compartments.
Key words: earthworms, bio-indicators, land use, soil nutrients, litter nutrients, Cupressus lusitanica, tropical cloud forest.
Resumen
La región Río Macho, un paisaje altamente fragmentado con riesgos inminentes de derrumbes e importantes servicios ambientales, ha sufrido una recuperación parcial de su vocación forestal por reforestación con especies exóticas y regeneración natural. Hemos comparado algunos rasgos del sustrato mediante la combinación del potencial bioindicador de los oligoquetos, rasgos del mantillo y características fisicoquímicos del suelo en tres sitios (bosque secundario viejo, tacotal y plantación Cupressus lusitanica). El bosque presentó la menor densidad de oligoquetos, el menor pH y el mayor contenido de humedad (CA) y fósforo del suelo. Hubo una correlación positiva entre los cocientes de C/N y C/P del mantillo y el pH del suelo, y negativa entre la profundidad, nitrógeno, fósforo del mantillo y fósforo del suelo. Los resultados sugieren que los oligoquetos son sensibles a cambios de los rasgos antes mencionados, que algunos de estos rasgos pueden ser buenos indicadores de la condición actual de regeneración de las coberturas estudiadas, y las coberturas han alcanzado concentraciones de varios nutrientes similares al bosque secundario viejo. Estos estudios a nivel ecosistémico a través de la evaluación de criterios sencillos pueden ayudarnos a inferir rápidamente el estado de algunos de los muchos y complejos compartimentos de los ecosistemas.
Palabras clave: lombrices, bioindicadoras, uso de la tierra, nutrientes del suelo, nutrientes del mantillo, Cupressus lusitanica, bosque tropical nuboso.
La deforestación es uno de los principales problemas que afecta directamente la dinámica natural de los ecosistemas tropicales (Quesada et al. 2009), lo que ha conducido en el último siglo a la conservación de remanentes de bosques maduros y la restauración de las zonas perturbadas. Las prácticas de restauración en los ecosistemas tropicales han estado dominadas por sistemas de regeneración natural y de reforestación con especies nativas y exóticas con valor comercial (Aide et al. 2000), tendencia que se ha repetido en Costa Rica (Quesada et al. 2009). La necesidad de evaluar éstos métodos de restauración sobre la dinámica de los procesos naturales de la recuperación nos ha llevado a enfocarnos en varios de los aspectos del funcionamiento del ecosistema. En este sentido, los oligoquetos, el mantillo y suelo, representan compartimentos claramente sensibles y por excelencia, indicadores de la salud del ecosistema terrestre (Jordan 1982, Facelli & Pickett 1991, Muys & Granval 1997).
De la fauna invertebrada del suelo, los oligoquetos son considerados los animales más importantes en el funcionamiento de los ecosistemas terrestres (Fraile 1986a, Fraile 1986b, León et al. 1988, Hendrix & Bohlen 2002), por su activa participación en el aumento de la fertilidad de los suelos y por su intervención en el incremento de la aireación, filtración, remoción y disponibilidad de nutrientes del suelo (Jones et al. 1994, Pashanasi et al. 1994). De este modo, estos organismos han sido muy utilizados en la vigilancia del estado y los cambios de la calidad del suelo en bosques y monocultivos (Muys & Granval 1997, Hendrix & Bohlen 2002). Específicamente, la abundancia de lombrices puede estar influenciada por aspectos como la compactación, la temperatura, la cantidad de nutrientes en el suelo y la estacionalidad (Fragoso & Lavelle 1992, Fragoso 2001). En la Amazonía central, Barros et al. (2004) encontraron que una vez que la tierra de uso pastoral es abandonada, ésta llega a estar dominada por una única especie de lombriz (Pontoscolex corethrurus). Esto demuestra cambios en la densidad y diversidad de los oligoquetos después de usos heterogéneos del suelo. Asimismo, un aumento en la población de lombrices se ha asociado con un incremento en la descomposición de la hojarasca, materia orgánica, cationes disponibles, y una disminución en la densidad del suelo (Tian et al. 1997, Tian et al. 2000, Jiménez et al. 2003). Además, se han encontrado diferencias contrastantes entre áreas usadas para plantación, áreas de bosque secundario y variaciones locales en la cantidad de oligoquetos en asociación con algunos tipos de vegetación (González & Zou 1999), lo que indica un efecto importante de la comunidad de plantas en la población de oligoquetos de diversas coberturas.
Por otro lado, el mantillo juega un papel transcendental en los procesos naturales del ecosistema, donde algunos investigadores han reconocido que el cambio de la cobertura vegetal tiene efectos directos sobre la constitución del mantillo, y este último determina la organización y estructura de las comunidades vegetales, a través de la germinación de las semillas, la sobrevivencia de plántulas y el desarrollo de retoños vegetales (Sydes & Grime 1981, Guzmán & Walker 1991). Asimismo, el mantillo es parte primordial en el proceso de almacenaje de carbono y elementos minerales en el suelo (Jordan 1982, Vogt et al. 1986), donde conjuntamente con la tasa de renovación de la biomasa subterránea constituye la vía principal de transferencia de nutrientes al suelo (Vitousek et al. 1995).
>El suelo sostiene una red trófica muy compleja cuyo funcionamiento resulta en último término en el ciclo de la materia orgánica y nutrientes (Domínguez et al. 2009), donde los nutrientes del suelo son producto del equilibrio de la biomasa vegetal depositada y la calidad de nutrientes que pueda contener, y donde los sitios ricos en nutrientes reflejan la biomasa y crecimiento de las plantas (Forde & Walch-Liu 2009). Por ejemplo, dada la heterogeneidad espacial de nutrientes del suelo, Zhang et al. (2010) encontraron una mayor proliferación de raíces en los sitios más ricos en nutrientes. Se ha demostrado que diferentes tipos de vegetación forman y mantienen diferentes calidades de suelos (Tanner 1980, Bach et al. 2003).
En este estudio, medimos algunas variables ligadas al compartimento del suelo en su capacidad de reflejar el estado de la restauración de diversos modos de recuperación forestal que representan dos prácticas antiguas de restauración boscosa en Costa Rica (un bosque secundario viejo, un tacotal y una plantación de Cupressus lusitanica Mill). Combinamos el potencial bioindicador de la presencia puntual de oligoquetos con algunas características específicas del mantillo y del suelo de tres coberturas forestales en recuperación, que representan estados contrastantes de regeneración natural y abandono. Intentamos comprobar si éstas coberturas difieren en cuanto a la densidad y biomasa de lombrices, estructura y nutrientes del mantillo y algunos rasgos fisicoquímicos del suelo. De esta manera, con la evaluación de estos criterios sencillos, se podría inferir rápidamente el estado de algunos rasgos del muy complejo sustrato de los ecosistemas forestales tropicales, y agilizar la toma de decisiones.
Materiales y Métodos
Sitio de estudio: Esta investigación se realizó en tres sitios situados en las faldas de las montañas al sur del Valle de Orosi, Costa Rica. El primer sitio contiene un bosque cercano a la Estación de Biología Tropical y Acuicultura de Río Macho (EBTARM, 9º45’52.64’’ N - 83º51’44.13’’ W), a 1 700m.n.s.m. y presenta una precipitación media anual de 2 416mm (Riba-Hernández 1996) y temperatura media mensual de 15.8ºC. Las especies arbóreas más frecuentes son: Viburnum costaricanum (Oerst.) Hemsl, Lozania mutisiana Schult, Symphonia globulifera L.F., Ilex lamprophylla Standl y Dendropanax arboreus (L.) Decne & Planch. El segundo sitio corresponde a un tacotal al costado oeste de la EBTARM (9º45’24.42” N - 83º50’59.16” W), cuya temperatura media mensual es de 16.5ºC. Algunas de las especies vegetales más frecuentes de este sitio son: Syzygium jambos (L.) Alston, Vismia baccifera (L.) Triana & Planch, Miconia dodecandra Cogn, Myrsine coriacea (Sw.) R. Br. ex Roem. & Schult y V. costaricanum. Según los registros de fotografías aéreas y encuestas a residentes de la zona, el bosque de referencia tiene aproximadamente 50 años de edad y el tacotal 13 años, ambos provenientes de potreros abandonados con suelos tipo ultisol (Tropohumult, iTCR 2008) y ubicados según Holdridge (1969) en la zona de vida de bosque muy húmedo montano bajo (bmh-MB). El último sitio corresponde a una plantación de ciprés (cipresal, C. lusitanica) sin manejo de al menos 35 años, establecida sobre un cafetal con suelos tipo inceptisol (Humitropept, iTCR 2008), rodeada de bosque ribereño y cafetales en producción, ubicada en la Finca La Laja, en Orosi (9º43’37.16” N - 83º52’00.38” W), a 1 300m.s.n.m. y en la misma vertiente de los sitios anteriores, cuya temperatura media mensual es de 18.4°C y la zona de vida corresponde al bosque muy húmedo premontano (bmh-P, Holdridge 1969). El sotobosque de este sitio esta colonizado por muchos individuos de algunas especies como: Miconia prasina (Sw.) DC., Miconia donaeana Naudin, V. costaricanum, Coffea arabica L. y S. jambos. A pesar de que este último sitio representa una zona de vida diferente y se encuentra a casi 400m más abajo, contiene un modo de restauración que ha sido controversial (Cavalier & Tobler 1998) y que merece ser evaluado bajo los parámetros establecidos en este estudio.
Diseño experimental: Se realizaron tres visitas a cada sitio durante julio y agosto 2009. En cada sitio se estableció una parcela de 5 000m2, dividida en subparcelas continuas de 100m2. En el centro de diez subparcelas escogidas al azar se delimitó un cuadrante de 0.0625m2, donde se midió la profundidad de la hojarasca y se recolectó el mantillo, definido aquí como la capa de residuos orgánicos que yace sobre el suelo de la comunidad vegetal (Facelli & Pickett 1991). Las muestras de mantillo se secaron a 60ºC durante 72hr y posteriormente se midió la biomasa seca separada en sus componentes de hojarasca y material leñoso (Sánchez-De León 2003). A cuatro muestras de la hojarasca del mantillo de los cuadrantes muestreados, se les analizó el contenido de carbono orgánico (CO), nitrógeno total, fósforo extraíble, calcio y magnesio por sitio. En los cuadrantes se removió el suelo hasta los 25cm de profundidad, y se separaron manualmente los oligoquetos (González et al. 2007), se lavaron con abundante agua, se contaron (n) y se secaron superficialmente con papel absorbente antes de obtener su biomasa fresca (b). Se calculó la densidad (ind/m2), biomasa fresca (g/m2) y la masa promedio de oligoqueto como b/n por muestra. La débil situación de la taxonomía de oligoquetos de Costa Rica nos previno de identificar las posibles especies encontradas en los muestreos. El único trabajo taxonómico en Costa Rica tiene 16 especies distribuidos en siete géneros de cuatro familias de regiones muy restringidas del país (Righi & Fraile 1987), un número no aceptable para la diversidad que podría existir en el trópico (Reynolds 1994).
El porcentaje del contenido de agua (CA) y pH del suelo se obtuvo de todos los hoyos donde se extrajeron los oligoquetos luego de ser secado en un horno a 100ºC durante 72hr (Anderson & Ingram 1993). Asimismo, en cada sitio se realizó el análisis de la materia orgánica (MO), carbono orgánico (CO), nitrógeno total, fósforo extraíble, azufre, acidez, capacidad de intercambio catiónico (CiC) y bases (calcio, potasio y magnesio), a cuatro muestras de suelo, de los hoyos de donde se extrajeron los oligoquetos y de donde se tomaron las muestras de hojarasca para análisis químicos.
Los análisis fisicoquímicos se efectuaron según Anderson & Ingram (1993), de la siguiente manera: La MO y el CO del suelo por digestión acida; el azufre, potasio y fósforo del suelo por el método de Olsen; el nitrógeno total del suelo y de la hojarasca por el método Kjeldahl; la acidez, calcio y magnesio del suelo por titulación con KCl; la CiC dado por la suma de bases; el CO de la hojarasca por incineración; el calcio y magnesio de la hojarasca por medio de absorción atómica; el fósforo de la hojarasca por el método de Molibdato-Vanadato de Amonio (Anderson & ingram 1993). Estos análisis fueron realizados y certificados por el Laboratorio de Química Marina de la Universidad Nacional de Costa Rica.
Todas las variables de los oligoquetos, mantillo y suelo fueron estudiadas mediante análisis de varianza de una vía (ANDEVA) clasificados por sitio. Los promedios de las variables medidas se confrontaron a posteriori con Pruebas de Bonferroni. Se realizaron correlaciones de Pearson entre todos los pares de variables de los tres sitios. Todas las variables del mantillo, suelo y parámetros químicos de la hojarasca se sometieron a un análisis multivariable en tres grupos (Bosque, tacotal y cipresal), definidos a priori en un espacio conformado por dos ejes en un análisis discriminante lineal (AD). La densidad y biomasa fresca de oligoquetos se transformaron al logaritmo base 10 para ajustar normalidad. Todos los resultados estadísticos fueron realizados con el programa R versión 2.12.1 (R Development Core Team 2010, Crawley 2007).
Resultados
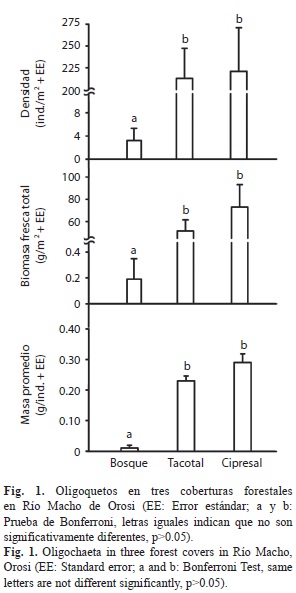
El bosque presentó la menor densidad, biomasa fresca y masa promedio de oligoquetos con respecto al tacotal y la plantación de ciprés (F2,27=71.0, R2=0.84, p<0.001; F2,27=55.4, R2=0.80, p<0.001 y F2,27=71.0, R2=0.84, p<0.001; respectivamente, Fig. 1), mientras que el tacotal y el cipresal no fueron diferentes en estas variables (Fig. 1).
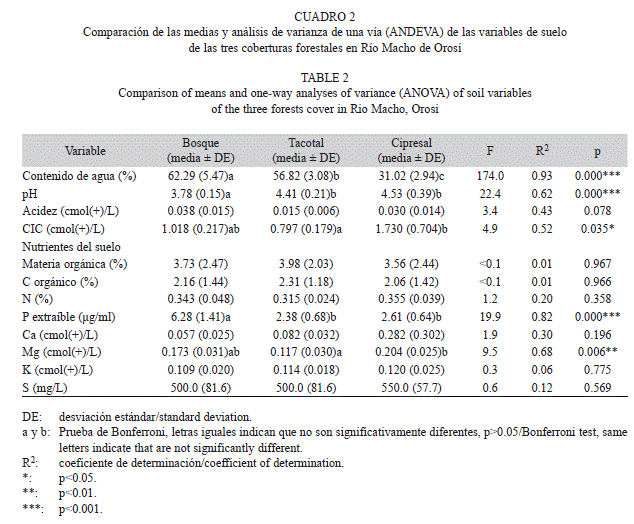
La cantidad de biomasa seca total del mantillo y sus componentes en hojarasca y material leñoso fueron similares entre los tres sitios (Cuadro 1), pese a esto, la profundidad del mantillo en el cipresal fue menor que en el bosque y en el tacotal. Estas dos últimas coberturas no fueron diferentes en la profundidad del mantillo (Cuadro 1). La concentración de nutrientes de la hojarasca presentó algunas diferencias entre los sitios, pero los ámbitos de variación fueron relativamente estrechos. La concentración de nitrógeno de la hojarasca fue similar en el tacotal y el cipresal, pero en ambos casos menores que en el bosque (Cuadro 1). El fósforo en la hojarasca del bosque fue mayor que el tacotal y el cipresal que tuvo concentraciones intermedias (Cuadro 1). Un patrón opuesto al anterior fue obtenido para la relación C/N en la hojarasca (Cuadro 1). El CO, el calcio, el magnesio y la relación C/P de la hojarasca no exhibieron diferencias significativas entre los tres sitios (Cuadro 1).
El bosque secundario presentó el menor pH del suelo en comparación con el tacotal y el cipresal, mientras que estas dos últimas coberturas fueron similares entre sí (Cuadro 2). Sin embargo, el bosque presentó el mayor porcentaje de CA y de P del suelo, seguido por el tacotal y por último el cipresal (Cuadro 2). En contraste, el cipresal destacó con el mayor CiC y magnesio del suelo con relación al tacotal, y el bosque presentó valores intermedios. La acidez, MO, CO, calcio, nitrógeno, potasio y azufre del suelo no mostraron diferencias significativas entre los tres sitios (Cuadro 2).
Hubo correlaciones positivas (Pearson, p<0.05) de los oligoquetos con las proporciones C/N y C/P de la hojarasca y pH del suelo (r>0.60, >0.60 y >0.40, respectivamente, Fig. 2), e inversas con la profundidad del mantillo, concentraciones de nitrógeno y fósforo de la hojarasca y fósforo del suelo (r<-0.37, <-0.65, <-0.65 y <-0.71, respectivamente, Fig. 2). Asimismo, hubo correlaciones entre algunos pares de nutrientes de la hojarasca y
y entre algunos nutrientes de hojarasca y del suelo (Pearson, p<0.05, Fig. 2).
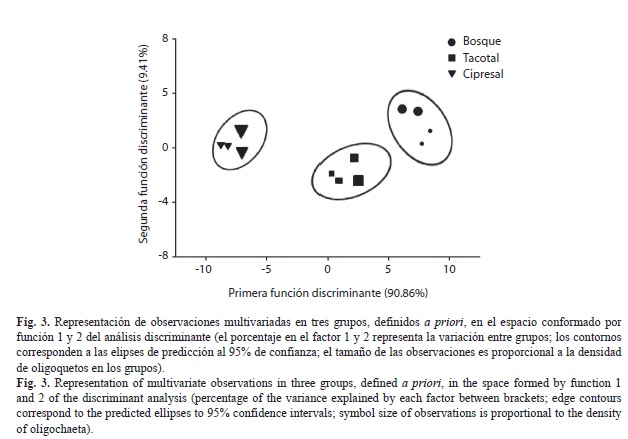
El AD realizado con todas las variables del suelo y mantillo produjo una clasificación muy acertada de los tipos de coberturas forestales (Fig. 3). De las 23 variables empleadas en el AD, sólo dos resultaron estadísticamente significativas (p<0.001): el porcentaje de CA del suelo (F=205.8, Lambda de Wilk=0.021) y el porcentaje de nitrógeno en la hojarasca (F=65.2, Lambda de Wilk=0.003). La primera función discriminante explicó el 90.86% (auto- valor=49.26) de la variación entre grupos y la segunda función el 9.14% (autovalor=4.96) restante (Fig. 3). Todas las observaciones fueron clasificadas correctamente en los sitios.
Discusión
La literatura reciente coincide en que los ecosistemas inalterados presentan una menor abundancia y biomasa de oligoquetos en comparación con zonas perturbadas (González et al. 1996, Zou & González 1997, Tondoh et al. 2011). Nuestros resultados apuntan hacia este mismo patrón. No obstante, la densidad y biomasa de oligoquetos para el bosque de referencia (3.2ind./m2 y 0.19g/m2, respectivamente) estuvo muy por debajo del promedio reportado por González et al. (2007), quienes encontraron hasta 110 individuos por m2 en un bosque de altura (1 010m.s.n.m.) en Puerto Rico. Por ejemplo, nuestro promedio en el bosque se asemeja más al obtenido en un ambiente extremo de un manglar (2.7ind/m2, González et al. 2007). La mayor abundancia y biomasa de oligoquetos en sistemas degradados puede deberse a la alta adaptabilidad de sus poblaciones en estos sistemas y a la predominancia de oligoquetos endógeas en todo el paisaje (Tondoh et al. 2011). Igualmente, algunas condiciones abióticas del suelo como lo es la disminución del pH, carbono orgánico, nitrógeno y fósforo se han visto asociados con la reducción de la abundancia de oligoquetos (Wang et al. 2005, Nelson et al. 2009). Por ejemplo, el bosque secundario viejo tuvo un pH del suelo bastante bajo (promedio 3.78, desv. estándar 0.15). Varios estudios ecológicos del suelo describen a los oligoquetos como ausentes en suelos ácidos (pH<3.5, Curry 1998, Mele & Carter 1999). La correlación inversa entre la densidad de oligoquetos y el pH del suelo confirma esta tendencia, la cual ha sido informada antes (González et al. 2007). Por otro lado, la complejidad y la diversidad de algunos ecosistemas favorecen la coexistencia a cambio de una reducción de la representación de algunos grupos de la comunidad (Aarssen 1983, Aarssen 1984, Worm & Karez 2002). En este sentido, la reducida abundancia de los oligoquetos en el bosque de referencia podría ser el producto de una mayor diversidad de otros macro-invertebrados. Por ejemplo, este tipo de cambio parece ir ocurriendo conforme avanzan los estados sucesionales a partir de pastizales abandonados (Sánchez-De León 2003)
La comunidad vegetal sobre el suelo es un precursor importante de la comunidad de oligoquetos (González & Zou 1999, Dechaine et al. 2005). Por ejemplo, algunos rasgos de la comunidad vegetal, tal como su composición y la tasa de incorporación de biomasa de hojarasca, pueden ser determinantes de la profundidad y calidad del mantillo (Facelli & Pickett 1991). A pesar de no haber diferencias entre los tres sitios en la biomasa de mantillo, si encontramos diferencias en su profundidad. La menor profundidad del mantillo del cipresal parece estar directamente relacionada con la estructura del material vegetal producida por el dosel (aproximadamente el 90% esta compuesto por ramas finas con hojas escamiformes). Éste tipo de material se deposita de manera compacta sobre el suelo, y se desarrolla en una capa de material vegetal de 10-30mm de espesor. De este modo, el gradiente de abundancia de oligoquetos en este estudio no esta directamente ligado a la cantidad ni estructura del mantillo (González et al. 2007). Es posible suponer que la supuesta acidificación del suelo por parte de los cipreses hubiera determinado una baja abundancia de oligoquetos en el cipresal, sin embargo esto no ocurrió. Del mismo modo, la baja concentración de nutrientes en la hojarasca de las coníferas (Lugo 1992a) y la del mantillo del tacotal con respecto al bosque en este estudio, y en especies sucesionales tempranas en otros estudios (Sprugel 1984, Kappelle & Leal 1996), no coinciden con la alta densidad de oligoquetos encontrados en estos sitios. Por último, la correlación inversa del porcentaje del CA en el suelo con la densidad, biomasa fresca y masa promedio de oligoquetos, es contraria a lo informado por otros investigadores (González et al. 2007). Según Fraile (1986b), cuando la humedad bajo la hojarasca se mantiene bastante alta es probable encontrar un aumento en la densidad de oligoquetos. Fraile (1986b) aclara que si la humedad incrementa demasiado los oligoquetos huyen a sitios menos húmedos. Sin embargo, los estudios de densidad de oligoquetos y humedad del suelo son controversiales debido a que no es sino hasta valores muy altos de humedad (>60%) donde las lombrices desaparecen (Bohlman et al. 1995, Sollins et al. 1993), lo que coincide con los altos valores de CA en nuestro bosque de referencia.
Los nutrientes del suelo como calcio, nitrógeno, potasio y azufre no fueron diferentes entre las tres coberturas forestales, aunque solo el cipresal difiere en el tipo de suelo. Los suelos de las plantaciones forestales viejas pueden contener concentraciones de nutrientes similares a bosques continuos, pese a las bajas concentraciones de nutrientes que puede contener la hojarasca depositada en los suelos de las plantaciones (Lugo 1992a). Es posible que la lenta recirculación de nutrientes en plantaciones provoque un aumento en el almacenamiento de nutrientes, como se ha sugerido en estudios similares con plantaciones exóticas (Lugo 1992a, Cavalier & Tobler 1998), lo que redunda en un acercamiento de los valores entre estos tres sitios.
La disponibilidad de fósforo del suelo fue mayor en el bosque (6.28μg/mL) que en el tacotal (2.5 veces) y la plantación de ciprés (2.4 veces). Cavalier & Tobler (1998) informan una concentración de fósforo de 5.3; 0.67μg/mL, muy por arriba a la informada en este estudio para nuestro cipresal (2.61; 0.64μg/mL). Es posible que el bosque secundario viejo con una mayor proporción de especies arborescentes primarias contenga también una mayor proporción de especies micorrízicas. La formación de micorrizas reduce la limitación debida a la baja movilidad del P (Semones 2000), que puede haber favorecido una mayor disponibilidad del fósforo hacia el suelo, el cual ha tenido tiempo de retornar más rápidamente en forma orgánica de nuevo al suelo (Bolan 1991). El contenido de nitrógeno y fósforo en la hojarasca sobre el suelo en las tres coberturas se correlacionó inversamente con la densidad, biomasa fresca y masa promedio de oligoquetos. Según Iordache & Borza (2010), el suplemento del fósforo en condiciones controladas de cultivos tienen un efecto negativo sobre la comunidad de oligoquetos, pero la adición de nitrógeno un tendencia positiva al incremento de su densidad, contario a lo encontrado en este estudio. El fósforo del suelo exhibió una correlación inversa con la densidad, y biomasa fresca de oligoqueto. Según Iordache & Borza (2010), el aumento del contenido de fósforo en el suelo tiene un efecto sobre la disminución de la biomasa de oligoquetos, aunque no siempre ocurre (Tian et al. 2000, Suárez et al. 2003).
El AD produjo una clasificación acertada de los tipos de coberturas forestales, con una mayor similitud del tacotal y el bosque, los cuales comparten el mismo tipo de suelo. Sin embargo, porcentaje del CA del suelo y nitrógeno en la hojarasca fueron las variables de mayor peso que separan claramente las tres coberturas, y que tienen una alta significancia sobre la comunidad de oligoquetos (Fraile 1986b, Wang et al. 2005, iordache & Borza 2010). Este análisis resalta, en última instancia, la importancia de la composición de la vegetación en la calidad de la hojarasca y cómo esta difiere según el estado sucesional del sitio. Además, a pesar de que la profundidad del mantillo, pH y el fósforo del suelo no fueron seleccionadas por el AD, no debemos descartar su consideración en la comparación de éstas u otras coberturas forestales, ya que ofrecen posibles explicaciones de procesos relacionados con la salud de estos ecosistemas.
Por último, es importante discutir algunos aspectos de las plantaciones de ciprés. Este estudio no apoya el paradigma sobre la disminución del pH del suelo por las plantaciones de coníferas (Grieve 1978, Sawyer 1993). Suponemos que el tiempo de regeneración del suelo y el efecto antropogénico de la agricultura anterior a la plantación y actualmente alrededor con cafetales activos, son las principales propuestas que explicarían porqué el pH del suelo no fue menor en la plantación en comparación al bosque. Por ejemplo, Cavellier & Tobler (1998) no encontraron cambios en el pH del suelo después de 17 años en una plantación de 21 años de edad de C. lusitanica que anteriormente fue usada como potrero. Esto podría indicar que el efecto antropogénico previo puede incidir sobre la dinámica del suelo incluso a largo plazo. Por otro lado, las plantaciones de coníferas como ecosistemas son catalogadas negativamente dada la baja riqueza de especies vegetales y animales que contienen, y por sus supuestas consecuencias sobre el aumento de la acidez del suelo (Grieve 1978, Sawyer 1993) y la podzolización (Savill & Evans 1986, Sawyer 1993). Pese a esto, algunos investigadores le atribuyen rasgos positivos (Cavellier & Tobler 1998), dado que las plantaciones tienden a acumular mayor biomasa y nutrientes sobre el suelo, contribuyendo más rápidamente a la restauración de la fertilidad de las mismas (Lugo 1992b). Además, plantaciones con baja densidad de árboles normalmente no excluyen a las especies nativas en el sotobosque (Lugo 1992b). En Costa Rica, se han contabilizado hasta 47 especies de árboles nativos en regeneración en el estrato inferior de cipresales viejos (Chaverri et al. 1998). De este modo, algunos cipresales abandonados podrían estar actuando como refugio de la riqueza de especies autóctonas, e incluso llegar a ofrecer algunos servicios ambientales al mismo nivel que ciertas áreas en estados secundarios de sucesión.
Varias conclusiones emergen de la combinación del potencial bioindicador de la presencia de oligoquetos, las características del mantillo y del suelo en una dinámica contrastante de restauración forestal. En primer lugar, los oligoquetos resultaron sensibles a cambios en algunas de las características del suelo (porcentaje CA, pH y fósforo) y del mantillo (nitrógeno, fósforo y relación C/N y C/P). Segundo, aparte de los oligoquetos, algunos rasgos del suelo (porcentaje de CA, pH, CiC, magnesio y fósforo) y mantillo (profundidad, nitrógeno, fósforo y proporción C/N) pueden servir por sí solos o en combinación, como indicadores del estado de las tres coberturas estudiadas. Por último, las similitudes de las concentraciones de nutrientes del suelo (carbono orgánico, nitrógeno, calcio, potasio y azufre) de los tres sitios, evidencian que el lapso de regeneración del cipresal (35 años) ha alcanzado condiciones de nutrientes similares al bosque secundario viejo, lo que supone una baja recirculación de nutrientes (Lugo 1992a). En el caso del tacotal, el uso anterior al abandono no parece haber degradado el suelo como para afectar los valores de estas variables.
En resumen, la restauración de los bosques tropicales requiere una cuidadosa valoración de las condiciones ambientales (es decir de suelos y clima), la selección de especies, y las condiciones de siembra (Cavalier & Tobler 1998). No podemos descartar que cipresales viejos, abandonados y autoraleados puedan llegar a ofrecer algunos servicios ecosistémicos e incluso ofrecer algunas condiciones similares a estadios secundarios jóvenes y viejos de sucesión forestal. Sin embargo, creemos que la plantación con árboles exóticos y estados iniciales e intermedios de recuperación natural podrían potenciar sus efectos ecosistémicos si se manejan con prácticas sencillas de restauración asistida que dependerían de las características del lugar y un plan estratégico de seguimiento y gestión (Cavalier & Tobler 1998, Yirdaw 2001).
Agradecimientos
Dina Solís del Laboratorio de Química Marina de la Universidad Nacional de Costa Rica (UNA) por la medición de los parámetros químicos del suelo y hojarasca. A Alicia Fonseca y Helga Madrigal del Laboratorio de Hidrología de la UNA. A cuatro revisores anónimos por sugerencias al manuscrito inicial. Esta investigación fue financiada con una beca del Fondo Especial para la Educación Superior (FEES) asignada por la Comisión Nacional de Rectores (CONARE) de Costa Rica a R.A.C.S.
Referencias
Aarssen, L.W. 1983. Ecological combining ability and competitive combining ability in plants: Toward a general evolutionary theory of coexistence in systems of competition. Am. Nat. 33: 67-83. [ Links ]
Aarssen, L.W. 1984. On the distinction between niche and competitive ability: implication for coexistence theory. Acta Biotheor. 33: 67-83. [ Links ]
Aide, T.M., J.K. Zimmerman, J.B. Pascarella, M. Rivera & H. Marcano-Vega. 2000. Forest regeneration in a chronosequence of tropical abandoned pastures: implications for restoration ecology. Restor. Ecol. 8: 328-338. [ Links ]
Anderson, J.M. & J.S.i. Ingram. 1993. Tropical soil biology and fertility: A handbook of methods. CAB international, Wallingford, Reino Unido. [ Links ]
Bach, K., M. Schawe, S. Beck, G. Gerold, S.R. Gradstein & M.R. Moraes. 2003. Vegetación, suelos y clima en los diferentes pisos altitudinales de un bosque montano de Yungas, Bolivia: Primeros resultados. Ecol. Bolivia 38: 3-14. [ Links ]
Barros, E., M. Grimaldi, M. Sarrazin, A. Chauvel, D. Mitja, T. Desjardins & P. Lavelle. 2004. Soil physical degradation and changes in macrofaunal communities in Central Amazon. Appl. Soil Ecol. 26: 157-168. [ Links ]
Bohlman, S.A., T.J. Matelson & N.M. Nadkarni. 1995. Moisture and temperature patterns of canopy humus and forest floor soil of a montane cloud forest, Costa Rica. Biotropica 27: 13-19. [ Links ]
Bolan, N.S. 1991. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 134: 189-207. [ Links ]
Cavalier, J. & A. Tobler. 1998. The effect of abandoned plantations of Pinus patula and Cupressus lusitanica on soils and regeneration of a tropical montane rain forest in Colombia. Biodivers. Conserv. 7: 335-347. [ Links ]
Chaverri, A., N. Zamora, V. Aguilar & J. Gutiérrez. 1998. Regeneración natural de especies nativas latifoliadas y de ciprés (Cupressus lusitanica) bajo una plantación de ciprés en San José de la montaña, Costa Rica. Agron. Costarricense 22: 7-17. [ Links ]
Crawley, W. 2007. The R Book. Wiley, Chichester, Inglaterra. [ Links ]
Curry, J.P. 1998. Factors affecting earthworm abundance in soils, p. 37-64. In C.A. Edwards (ed.). Earthworm ecology. Santa Lucia, Boca Ratón, Florida, EEUU. [ Links ]
Dechaine, J., H. Ruan, Y. Sanchez-De León & x. Zou. 2005. Correlation between earthworms and plant litter decomposition in a tropical wet forest of Puerto Rico. Pedobiologia 49: 601-607. [ Links ]
Domínguez, J., M. Aira & M. Gómez-Brandón. 2009. El papel de las lombrices de tierra en la descomposición de la materia orgánica y el ciclo de nutrientes. Ecosistemas 18: 20-31. [ Links ]
Facelli, J.M. & S.T.A. Pickett. 1991. Plant litter: its dynamics and effects on plant community structure. Bot. Rev. 57: 1-32. [ Links ]
Forde, B.G. & P. Walch-Liu. 2009. Nitrate and glutamate as environmental cues for behavioural responses in plant roots. Plant. Cell. Environ. 32: 682-693. [ Links ]
Fragoso, C. 2001. Las lombrices de tierra de México (Annelida, Oligochaeta): Diversidad, ecología y manejo. Acta Zool. Mex. 1: 131-171. [ Links ]
Fragoso, C. & P. Lavelle. 1992. Earthworm communities of tropical rain forests. Soil Biol. Biochem. 24: 1397-1408. [ Links ]
Fraile, J. 1986a. Las lombrices de tierra contribuyen al aprovechamiento de los recursos naturales. Biocenosis 2: 46-50. [ Links ]
Fraile, J. 1986b. Participación de las lombrices de tierra en el ciclo del nitrógeno, con énfasis en los sistemas agroforestales de Costa Rica. Uniciencia 3: 119-124. [ Links ]
González, G., E. García, V. Cruz, S. Borges, M. Zalamea & M. Rivera. 2007. Earthworm communities along an elevation gradient in Northeastern Puerto Rico. Europe. J. Soil Biol. 43: S24-S32. [ Links ]
González, G. & x. Zou. 1999. Plant and litter influence in earthworm abundance and community structure in a tropical wet forest. Biotropica 31: 486-493. [ Links ]
González, G., x. Zou & S. Borges. 1996. Earthworm abundance and species composition in abandoned tropical croplands: comparisons of tree plantations and secondary forests. Pedobiologia 40: 385-391. [ Links ]
Grieve i.C. 1978. Some effects of the plantation of conifers on a freely-drained lowland soil, Forest of Dean. Forestry 51: 21-8. [ Links ]
Guzmán, S.M. & L.R. Walker. 1991. Differential seeding responses to litter after hurricane Hugo in the Luquillo Experimental Forest, Puerto Rico. Biotropica 23: 407-413. [ Links ]
Hendrix, P.F. & P.J. Bohlen. 2002. Exotic earthworm invasions in North America: Ecological and policy implications. Bioscience 52: 801-811. [ Links ]
Holdridge, L.R. 1967. Life zone ecology. Tropical Science Center, San José, Costa Rica. [ Links ]
Iordache, M. & i. Borza. 2010. Relation between chemical indices of soil and earthworm abundance under chemical fertilization. Plant Soil Environ. 56: 401-407. [ Links ]
ITCR. 2008. Atlas de Costa Rica. Laboratorio de Sistemas de información Geográfica, instituto Tecnológico de Costa Rica, Cartago, Costa Rica. [ Links ]
Jiménez, J.J., A. Cepeda, T. Decaëns, A. Oberson & D.K. Friesen. 2003. Phosphorus fractions and dynamics in surface earthworm casts under native and improved grasslands in a Colombian savanna Oxisol. Soil. Biol. Biochem. 35: 715-727. [ Links ]
Jones, C.G., J.H. Lawton & M. Shachak. 1994. Organisms as ecosystem engineers. Oikos 69: 373-386. [ Links ]
Jordan, C.F. 1982. The nutrient balance of an Amazonian rain forest. Ecology 63: 647-654. [ Links ]
Kappelle, M. & M.E. Leal. 1996. Changes in leaf morphology and foliar nutrient status along a successional gradient in a Costa Rica upper montane Quercus Forest. Biotropica 28: 331-344. [ Links ]
León, S., N. González, M. Lobo & J. Fraile. 1988. Poblaciones de lombrices de tierra en sitios de acumulación de desechos orgánicos en el Valle Central de Costa Rica. Uniciencia 5: 61-68. [ Links ]
Lugo, A.E. 1992a. Comparison of tropical tree plantations with secondary forests of similar age. Ecol. Monogr. 62: 1-41. [ Links ]
Lugo, A.E. 1992b. Tree plantations for rehabilitating damaged Forest Lands in the tropics, p. 247-255. In M.K. Balli (ed.). Ecosystem rehabilitation: Ecosystem analysis and synthesis. SPB, La Haya, Holanda. [ Links ]
Mele, P.M. & M.R. Carter. 1999. Impact of crop management factors in conservation tillage farming on earthworm density, age structure and species abundance in south-eastern Australia. Soil Till. Res. 50: 1-10. [ Links ]
Muys, B. & P.H. Granval. 1997. Earthworms as bioindicators of forest site quality. Soil Biol. Biochem. 29: 323-328. [ Links ]
Nelson, K.L., D.H. Lynch & G. Boiteau. 2009. Assessment of changes in soil health throughout organic potato rotation sequences. Agr. Ecosyst. Environ. 131: 220-228. [ Links ]
Pashanasi, B., P. Lavalle & J. Alegre. 1994. Efecto de lombrices de tierra (Pontoscolex corethrurus) sobre el crecimiento de cultivos anuales y características físicas y químicas en suelos de Yurimaguas. Fol. Amazon. 6: 5-46. [ Links ]
Quesada, M., G.A. Sanchez-Azofeifa, M. Alvarez-Anorve, K.E. Stoner, L. Avila-Cabadilla, J. Calvo-Alvarado, A. Castillo, M.M. Espirito-Santo, M. Fagundes, G.W. Fernández, J. Gamon, M. Lopezaraiza-Mikel, D. Lawrence, P. Morellato, J. Powers, F.S. Neves, V. Rosas-Guerrero, R. Sayago & G. Sanchez-Montoya. 2009. Succession and management of tropical dry forests in the Americas: Review and new perspectives. Forest. Ecol. Manag. 258: 1014-1024. [ Links ]
R. Development Core Team. 2010. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (Consultado: 28 agosto, 2011, www.R-project.org/. [ Links ]).
Reynolds, J. 1994. Earthworms of the world. Global Biodiversity 4: 11-16. [ Links ]
Riba-Hernández, P. 1996. Notas sobre los murciélagos de la Estación Biológica Río Macho, Costa Rica. Brenesia 45: 183-186. [ Links ]
Righi, G. & J.M. Fraile. 1987. Alguns oligochaeta de Costa Rica. Rev. Brasil. Biol. 47: 535-548. [ Links ]
Sánchez-De León, Y., x. Zou, S. Borges & H. Ruan. 2003. Recovery of native earthworms in abandoned tropical pastures. Conserv. Biol. 17: 999-1006. [ Links ]
Savill, P.S. & J. Evans. 1986. Plantation Sylviculture in Temperate Regions: with Special Reference to the British Isles. Oxford, Clarendon, EEUU. [ Links ]
Sawyer, J. 1993. Plantations in the tropics: Environmental concerns. UICN, Gland, Switzerland and Cambridge, United Kingdom in collaboration with UNEP and WWF. [ Links ]
Semones, S. 2000. Mycorrhizae and plant stress physiology, p. 108-175. In D.M. Orcut & E.T. Nilsen (eds.). Physiology of plants under stress. Wilson, Virginia, EEUU. [ Links ]
Sollins, P.F., F.M. Sancho, R.C. Mata & R.L. Sanford. 1993. Soils and soil process research, p. 40-41. In. L.A. McDade, K.S. Bawa, H.A. Hespenheide & G.S. Hartshorn (eds.). La Selva: ecology and natural history of a neotropical rainforest. University of Chicago, Chicago, Illinois, EEUU. [ Links ]
Sprugel, D. 1984. Density, biomass, productivity, and nutrient-cycling changes during stand development in wave-regenerated Balsam fir forest. Ecol. Monogr. 54: 165-186. [ Links ]
Suárez, E.R., D.M. Pelletier, T.J. Fahey, P.M. Groffman, P.J. Bohlen & M.C. Fisk. 2003. Effects of exotic earthworms on soil phosphorus cycling in two broadleaf temperate forests. Ecosystems 7: 28-44. [ Links ]
Sydes, C. & J.P. Grime. 1981. Effects of tree leaf litter on herbaceous vegetation in deciduous woodland: ii. An experimental investigation. J. Ecol. 69: 249-262. [ Links ]
Tanner, E.V.J. 1980. Studies on the biomass and productivity in a series of montane rain forests in Jamaica. J. Ecol. 68: 573-588. [ Links ]
Tian, G., B.T. Kang & L. Brussaard. 1997. Effect of mulch quality on earthworm activity and nutrient supply in the humic tropics. Soil Biol. Biochem. 29: 369-373. [ Links ]
Tian G., J.A. Olimaha, G.O. Adeoye & B.T. Kang. 2000. Regeneration of earthworm populations in a degraded soil by natural and planted fallows under humid tropical conditions. Soil Sci. Soc. Am. J. 64: 222-228. [ Links ]
Tondoh, J.E., A.M. Guéi, C. Csuzdi & P. Okoth. 2011. Effect of land-use on the earthworm assemblages in semi-deciduous forests of Central-West Ivory Coast. Biodivers. Conserv. 20: 169-184. [ Links ]
Vázquez, C.Y., A.S. Orozco, E. Rincón, M.C. Sánchez, P. Huante, J.R. Toledo & V.L. Barradas. 1990. Light beneath the litter in a tropical forest: Effect on seed germination. Ecology 71: 1952-1958. [ Links ]
Vitousek, P.M., G. Gerrish, D.R. Turner, L.R. Walker & D. Mueller-Dumbois. 1995. Litter fall and nutrient cycling in four Hawaiian montane rainforest. Trop. Ecol. 11: 189-203. [ Links ]
Vogt, K.A., C.C. Grier & D.L. Vogt. 1986. Production, turnover and nutrient dynamics of above-ground detritus of world forests. Adv. Ecol. Res. 15: 303-377. [ Links ]
Wang, x., F. Hu & H. Li. 2005. Contribution of earthworm activity to the infiltration of nitrogen in a wheat agroecosystem. Biol. Fertil. Soils 41: 284-287. [ Links ]
Worm, B. & R. Karez. 2002. Competition, coexistence and diversity on rocky shores, p. 144-157. In U. Sommer & B. Worm (eds.). Competition and coexistence. Springer, Alemania. [ Links ]
Yirdaw, E. 2001. Diversity of naturally-regenerated native woody species in forest plantations in the Ethiopian highlands. New Forests 22: 159-177. [ Links ]
Zhang, Y., Z. Zhou, x. Ma & G. Jin. 2010. Foraging ability and growth performance of four subtropical tree species in response to heterogeneous nutrient environments. J. Forest. Res. 15: 91-98. [ Links ]
Zou, x. & G. González. 1997. Changes in earthworm density and community structure during secondary succession in abandoned tropical pastures. Soil Biol. Biochem. 29: 627-629. [ Links ]
*Correspondencia: Junior Pastor Pérez-Molina & Roberto A. Cordero Solórzano: Laboratorio de Ecología Vegetal Funcional (LEVEF), Escuela de Ciencias Biológicas, Universidad Nacional de Costa Rica, Campus Omar Dengo, Heredia, Costa Rica; jpastorpm@gmail.com, ticolamb@gmail.com
1. Laboratorio de Ecología Vegetal Funcional (LEVEF), Escuela de Ciencias Biológicas, Universidad Nacional de Costa Rica, Campus Omar Dengo, Heredia, Costa Rica; jpastorpm@gmail.com, ticolamb@gmail.com
Recibido 26-ix-2011. Corregido 18-iii-2012 Aceptado 17-iV-2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}