










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Composition and abundance of small mammal communities in forest fragments and vegetation corridors in Southern Minas Gerais, Brazil
*Dirección para correspondencia
Abstract
Habitat fragmentation leads to isolation and reduce habitat areas, in addition to a series of negative effects on natural populations, affecting richness, abundance and distribution of animal species. In such a text, habitat corridors serve as an alternative for connectivity in fragmented landscapes, minimizing the effects of structural isolation of different habitat areas. This study evaluated the richness, composition and abundance of small mammal communities in forest fragments and in the relevant vegetation corridors that connect these fragments, located in Southern Minas Gerais, Southeastern Brazil. Ten sites were sampled (five forest fragments and five vegetation corridors) using the capture-mark-recapture method, from April 2007-March 2008. A total sampling effort of 6 300 trapnights resulted in 656 captures of 249 individuals. Across the 10 sites sampled, 11 small mammal species were recorded. Multidimensional scaling (MDS) ordinations and ANOSIM based on the composition of small mammal communities within the corridor and fragment revealed a qualitative difference between the two environments. Regarding abundance, there was no significant difference between corridors and fragments. In comparing mean values of abundance per species in each environment, only Cerradomys subflavus showed a significant difference, being more abundant in the corridor environment. Results suggest that the presence of several small mammal species in the corridor environment, in relatively high abundances, could indicate corridors use as habitat, though they might also facilitate and/or allow the movement of individuals using different habitat patches (fragments).
Key words: rodents, marsupials, fragmentation, vegetation corridors, Brazilian Atlantic forest.
Resumen
La fragmentación del hábitat conduce al aislamiento y la reducción de los hábitats, además provoca una serie de efectos negativos sobre las poblaciones naturales, afectando la riqueza, abundancia y distribución de las especies de animales. Dentro de este contexto, los corredores biológicos sirven como una alternativa para la conectividad de los paisajes fragmentados, minimizando los efectos del aislamiento estructural de las áreas con diferentes hábitats. Este estudio evaluó la riqueza, la composición y la abundancia de las comunidades de mamíferos pequeños en fragmentos de bosque y en los corredores de vegetación relevantes que conectan estos fragmentos, localizados en el sur de Minas Gerais, sudeste de Brasil. Diez sitios fueron muestraeados (cinco fragamentos de bosque y cinco corredores biológicos ) usando el método de captura-marcaje-recaptura, desde abril de 2007-marzo de 2008. Un esfuerzo total de muestreo de 6 300 trampas nocturnas resultó en 656 capturas de 249 individuos. En los 10 sitios muestreados, se registraron 11 especies de mamíferos pequeños. Las ordenaciones del escalamiento Multidimensional (MDS) y el ANOSIM basados en la composición de las comunidades de mamíferos pequeños dentro de los corredores y los fragmentos revelan una diferencia cualitativa entre estos dos ambientes. En cuanto a la abundancia, no hubo una diferencia significativa entre los corredores y los fragmentos. Al comparar los valores promedio de abundancia por especie en cada ambiente, sólo Cerradomys subflavus mostró una diferencia significativa, siendo más abundante en el ambiente del corredor biológico. Los resultados sugieren que la presencia de varias especies de mamíferos pequeños en el entorno del corredor biológico, en abundancias relativamente altas, podría indicar el uso de los corredores como hábitat, aunque estos también podrían facilitar y/o permitir el movimiento de individuos que utilizan los diferentes parches de hábitat (fragmentos).
Palabras clave: roedores, marsupiales, fragmentación, corredores biológicos, bosque Atlántico brasileño.
Habitat fragmentation leads to isolation and reduce habitat areas, in addition to a series of negative effects on natural populations, affecting richness, abundance and distribution of animal species. In such a text, habitat corridors serve as an alternative for connectivity in fragmented landscapes, minimizing the effects of structural isolation of different habitat areas. This study evaluated the richness, composition and abundance of small mammal communities in forest fragments and in the relevant vegetation corridors that connect these fragments, located in Southern Minas Gerais, Southeastern Brazil. Ten sites were sampled (five forest fragments and five vegetation corridors) using the capture-mark-recapture method, from April 2007-March 2008. A total sampling effort of 6 300 trapnights resulted in 656 captures of 249 individuals. Across the 10 sites sampled, 11 small mammal species were recorded. Multidimensional scaling (MDS) ordinations and ANOSIM based on the composition of small mammal communities within the corridor and fragment revealed a qualitative difference between the two environments. Regarding abundance, there was no significant difference between corridors and fragments. In comparing mean values of abundance per species in each environment, only Cerradomys subflavus showed a significant difference, being more abundant in the corridor environment. Results suggest that the presence of several small mammal species in the corridor environment, in relatively high abundances, could indicate corridors use as habitat, though they might also facilitate and/or allow the movement of individuals using different habitat patches (fragments).
Key words: rodents, marsupials, fragmentation, vegetation corridors, Brazilian Atlantic forest.
Resumen
La fragmentación del hábitat conduce al aislamiento y la reducción de los hábitats, además provoca una serie de efectos negativos sobre las poblaciones naturales, afectando la riqueza, abundancia y distribución de las especies de animales. Dentro de este contexto, los corredores biológicos sirven como una alternativa para la conectividad de los paisajes fragmentados, minimizando los efectos del aislamiento estructural de las áreas con diferentes hábitats. Este estudio evaluó la riqueza, la composición y la abundancia de las comunidades de mamíferos pequeños en fragmentos de bosque y en los corredores de vegetación relevantes que conectan estos fragmentos, localizados en el sur de Minas Gerais, sudeste de Brasil. Diez sitios fueron muestraeados (cinco fragamentos de bosque y cinco corredores biológicos ) usando el método de captura-marcaje-recaptura, desde abril de 2007-marzo de 2008. Un esfuerzo total de muestreo de 6 300 trampas nocturnas resultó en 656 capturas de 249 individuos. En los 10 sitios muestreados, se registraron 11 especies de mamíferos pequeños. Las ordenaciones del escalamiento Multidimensional (MDS) y el ANOSIM basados en la composición de las comunidades de mamíferos pequeños dentro de los corredores y los fragmentos revelan una diferencia cualitativa entre estos dos ambientes. En cuanto a la abundancia, no hubo una diferencia significativa entre los corredores y los fragmentos. Al comparar los valores promedio de abundancia por especie en cada ambiente, sólo Cerradomys subflavus mostró una diferencia significativa, siendo más abundante en el ambiente del corredor biológico. Los resultados sugieren que la presencia de varias especies de mamíferos pequeños en el entorno del corredor biológico, en abundancias relativamente altas, podría indicar el uso de los corredores como hábitat, aunque estos también podrían facilitar y/o permitir el movimiento de individuos que utilizan los diferentes parches de hábitat (fragmentos).
Palabras clave: roedores, marsupiales, fragmentación, corredores biológicos, bosque Atlántico brasileño.
Brazil is among the richest countries in the world in animal species diversity, and it is the richest in mammal species (Costa et al. 2005). Despite high rates of deforestation, the Atlantic forest of Brazil host an enormous biological diversity and a very high degree of endemism, being considered one of the most important biodiversity hotspots (Myers et al. 2000, Brooks et al. 2002, Orme et al. 2005, Ceballos & Ehrlich 2006). Forest fragmentation and loss of habitat constitute major threats to biodiversity, particularly in tropical regions, where population density and outspread agriculture are increasing rapidly (Saunders et al. 1991, Turner 1996, Cuarón 2000, Fahrig 2003, Bennett et al. 2006). Fragmentation leads to isolation too and triggers a number of negative effects on wildlife populations, affecting the richness, abundance, and distribution of species (Brooks et al. 2002, Fahrig 2002, Fischer & Lindenmayer 2007, Pardini et al. 2009, vieira et al. 2009). On a broader time scale, fragmentation also affects genetic variability and persistence of populations, potentially leading to local extinctions (Fahrig & Merriam 1994, Gibbs 2001, viveiros de Castro & Fernandez 2004). To survive in fragmented landscapes the individuals need to move between patches of habitat in search of conditions and resources, and many species are unable to travel across open matrix areas (Knaapen et al. 1992, Gascon et al. 1999, Umetsu & Pardini 2007, Passamani & Fernandez 2011a,b). Studies suggest that vegetation corridors could act as habitat or facilitators of dispersion and genetic exchange between populations on fragmented landscapes (Bennett 1990, Aars & Ims 1999, Mech & Hallet 2001).
One of the focuses of related research has been the use of vegetation corridors by small mammals especially in temperate countries (Hobbs 1992, Aars & Ims 1999, Haddad et al. 2003) while in tropical forests there are few studies (Bennett 1990, Downes et al. 1997, Pardini et al. 2005, Rocha et al. 2011). The small mammals responses to environmental changes like fragmentation, edge effect, matrix quality, and corridor presence (Saunders et al. 1991, Malcolm 1994, Lidicker 1999, Pardini et al. 2005, Umetsu & Pardini 2007, Passamani & Fernandez 2011a) and their responses, can help researchers understand ecological processes like population dynamics and community structure, and in particular the role of corridors in metapopulation dynamics (Lidicker 1999). Overall, generalist species respond positively to fragmentation, persisting in modified areas and increasing their densities, while species that are more selective in habitat and diet reduce their densities and often become locally extinct (Pardini 2004, viveiros de Castro & Fernandez 2004, Pütker et al. 2008, Pardini et al. 2010). In Southern Minas Gerais, Brazil, the landscape consists of many small forest fragments imersed in an anthropogenic matrix (agricultural crops and pastures) and some of these fragments are connected through vegetation corridors. However studies concerning the composition and abundance on faunal species in these corridors, are incipients. Thus, the aim of this study was to evaluate the richness, composition, and abundance of small-mammal communities in forest fragments and in connecting vegetation corridors.
Materials and methods
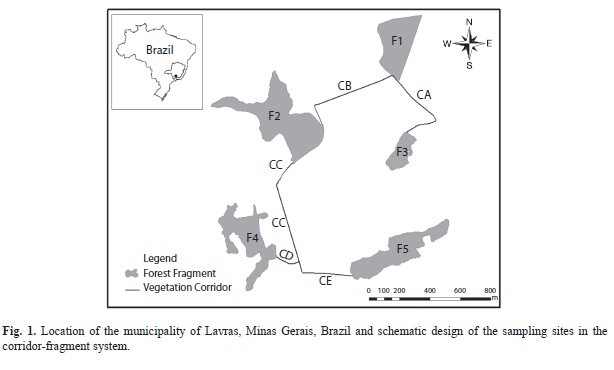
Study site: The corridor-fragment system studied is located in the municipality of Lavras, in Southern Minas Gerais, Brazil (21º17’15.1’’ S - 44°58’59.3’’ W), at ca. 950m of altitude (Castro 2004). The climate of the region is the Cwa of Koeppen’s classification, with dry winters and wet summers, and the average annual temperature is 20.4°C. The average annual precipitation is 1 460mm, with a rainy season from November through February (Dantas et al. 2007). The local vegetation is defined as being a disjointed portion of Cerrado biome, inserted into the semideciduous, seasonal Atlantic forest region of Southeastern Brazil (Oliveira-Filho et al. 1994), and the areas sampled are comprised of secondary forests at different regeneration stages. The small mammals were sampled in five forest fragments (F1, F2, F3, F4 and F5) and five vegetation corridors (CA, CB, CC, CD and CE) connecting them (Fig. 1). The forest fragments size was F1=7.2ha, F2=11.8ha, F3=1.03ha, F4=7.4ha and F5=7.8ha. According to Castro (2004) the origin of the vegetation corridors in Southern Minas Gerais can be connected to two different factors: One of them would be the linear strips of remnant vegetation left after clearcut in the forests to create boundaries between rural properties (also known as fencerows or headgerows). The other factor would be the natural trees colonization of ditches, excavations built by slaves in the late 19th Century to divide rural properties. The vegetation corridor studied has 4m width and ca. 1.5m depth and length was CA=568m, CB=667m, CC=789m, CD=170m and CE=333m.
Sampling: In each fragment, two transects of 100m were established, 50m apart, with one capture station placed along each transect at 20m spacing. Two traps were set in each capture station: one on the ground and one in the understory, fastened in tree branches (between 1-2m height). In each corridor, one transect of 100m was established, and traps were distributed as with the fragments. On the ground of each transect two Tomahawk traps (45.0×16.0×16.0cm), two Sherman small traps (25.0× 9.0×8.0cm), and two Sherman big traps (43.0×12.5×14.5cm), were placed alternately. For the understory portion, only Sherman small traps were used. Traps were baited with banana, commercial codfish oil, peanut butter and corn flour that were replaced every two days. The traps were placed at dusk and checked the following morning for five consecutive nights per month, during April 2007-March 2008. In the first two months, the corridors and fragments were sampled together, but over the 10 subsequent months, the sites were sampled alternately, with a total of seven sampling months in each site. The capture-mark-recapture method was used, and the animals captured were marked with numbered ear tags (National Band & Tag Co., Newport, KY, USA) and then released at the same point of capture. The marsupials were identified according to Rossi et al. (2006) and the rodents according to Oliveira & Bonvicino (2006), except Cerradomys subflavus and Calomys cerqueirai, for which the classification proposed by Weksler et al. (2006) and Bonvicino et al. (2010) was used, respectively. Some specimens of all species were collected and hosted to the collection of the Laboratory of Ecology and Conservation of Mammals of the Universidade Federal de Lavras.
To compare the richness of corridor and fragment sites, rarefaction curves were constructed using EstimateS 8.0 software (Colwell 2008). To evaluate abundance, only the first capture of each individual was considered. To verify if there were differences between the total abundance of fragments and corridors, and between the abundance of each species in these two environments, the Student’s t-test was performed using STATISTICA 6.0 software (StatSoft 2001) with a significance level of 0.05. The extrapolation of group patterns of the multidimensional scaling (MDS) ordination was used for analyze community composition and abundance in the fragments and corridors, and similarity matrices of communities were constructed using the Bray-Curtis similarity index. Additionally, we used one way analysis of similarities (ANOSIM) to test for the significant differences in composition and abundance of small-mammal communities between corridor and fragment sites. Both analyses were performed using PRIMER v-5 software (Clarke & Gorley 2001).
Results
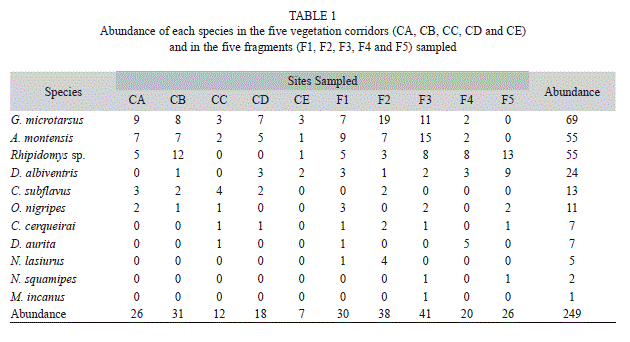
A total sampling effort of 6 300 trapnights resulted in 656 captures of 249 individuals, corresponding to a capture success rate of 10.4%. Overall, 11 small mammal species were recorded: Akodon montensis, Calomys cerqueirai, Cerradomys subflavus, Necromys lasiurus, Nectomys squamipes, Oligoryzomys nigripes, Rhipidomys sp., Didelphis albiventris, D. aurita, Gracilinanus microtarsus and Marmosops incanus.
The rarefaction curves (Fig. 2) produced from the sampling effort of 70 individuals showed that fragments have higher species richness, with around 30% more species sampled than in vegetation corridors. Out of the total number of captures, 59% were rodents and 41% were marsupials, with rodents being more abundant both in corridors (61%) and in fragments (59%). The most abundant species were G. microtarsus (69 individuals), A. montensis (55) and Rhipidomys sp. (55), while the least abundant were N. lasiurus (5), N. squamipes (2) and M. incanus (1) (Table 1).
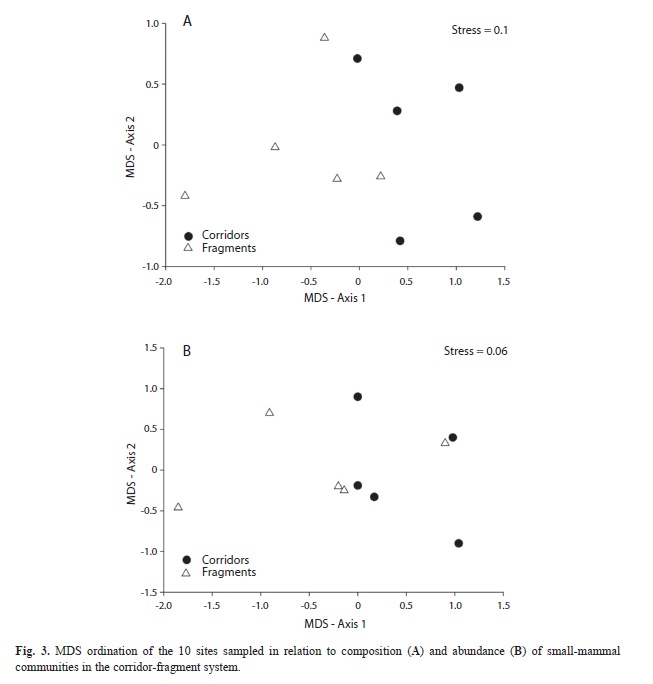
Considering each type of environment, the abundance of small mammals in corridors was 4.48 (SD=±2.34) and in fragments was 3.69 (SD=±1.02), but the difference was not significant (t=0.69, p=0.50). When comparing mean abundance per species in each environment, only C. subflavus showed a significant difference (t=-2.89, p=0.02), being more abundant in corridor environments than in fragments. The group pattern ordination analysis (MDS) suggested a difference in the composition of smallmammal communities (presence and absence) from corridor to fragment, with two spatially distinct groups noted (Fig. 3A). While for the abundance, the MDS did not detect any difference between the small mammal communities in these environments (Fig. 3B).
Patterns depicted by the MDS analysis for the small-mammal species in the corridor-fragment system were reinforced by the ANOSIM analysis, with corridors differing from fragments in terms of species composition (ANOSIM, Global R=0.27, p=0.03) but not in terms of abundance (ANOSIM, Global R=0.02, p=0.43).
Discussion
These results demonstrate that smallmammal communities present in the corridors and fragments differ in species richness and composition but are similar with respect to abundance. This difference could be related to the presence of three species (N. squamipes, N. lasiurus and M. incanus) exclusive to fragments. Nectomys squamipes was detected only in two of the fragments (F3 and F5) that have areas boasting watercourses and permanently flooded soil. This species has semiaquatic habits (Bonvicino et al. 2008); hence its presence was restricted to these two fragments. The occurrence of N. lasiurus was restricted to another two fragments (F1 and F2), but both fragments showed low abundance. This species appears in open and forested Cerrado formations and in the transitional Cerrado- Atlantic forest ecotone (Bonvicino et al. 2008), it being the dominant species in cerrado and campo sujo habitats, but not in riparian forests (Alho 1981). According to Castro (2004), these fragments (F1 and F2) have floristic characteristics common to Cerrado areas which explains the presence of N. lasiurus. The other exclusive species was M. incanus, recorded only once in one of the fragments (F3). Considering our data during a year of study, the presence of M. incanus may be a casual recorded of as individual. Its occasional capture is probably related to an isolated event of movements, since typically this species moves across open areas (Passamani & Fernandez 2011b), occupying portions of agroforest crops (Fonseca 1997, Passamani & Ribeiro 2009).
In the corridor-fragment system, only C. subflavus was more frequent and significantly more abundant in the corridors than in fragment sites, a finding that is similar to verified by Rocha et al. (2011) regarding corridors connecting fragments near this area. In addition, the studied corridors harbour a considerable number of species in common with the fragments and similarity in abundance. The presence of small-mammal species in corridor and fragment sites could be related to the fact that corridors effectively play the role of habitats for small-mammal species rather than being mere linking environments for their movement.
Several studies in temperate countries have pointed to the effectiveness of linear vegetation corridors in preserving small mammals’ fauna. Wegner & Merriam (1979) monitored the movements of small mammals in forest fragments and fencerows in Canada and detected movements of three small-mammal species, most of which occurred between fragments and fencerows, and rarely in cultivated fields. Andreassen et al. (1998), in studying the wild rodent Microtus oeconomus, observed that corridor presence increases the chance of individuals incorporating more than one habitat patch into their home range. Bolger et al. (2001), in evaluating the use of remnant shrub strips and straight-line revegetated areas connecting forest remainder in California (USA), found no significant difference in the richness of small-mammal species between corridors and forest remainders. Haddad & Tewksbury (2006) evaluated the effects of vegetation corridors on the maintenance of local and regional small mammal biodiversity and verified that the presence of corridors is much important to the environmental management.
It should be noted that, in terms of structure, hedgerows and fencerows in temperate countries resemble the vegetation corridors evaluated in this study, yet they strongly differ in their floristic composition, comprised typically of a single or a few arboreal species. The corridors of Southern Minas Gerais boast a robust forest structure and high floristic diversity (Castro 2004), which may positively influence food and shelter availability for small-mammal. These facts point to the important role of corridors in attenuating the negative effects of fragmentation on small-mammal fauna on a local and/or regional scale. In fact, Rocha et al. (2011) has shown the high conservation value of this structure in another fragmented landscape of Southern Minas Gerais.
In conclusion, corridors seem to be effective in the exchange of individuals between forest fragments, promoting similarity in species composition within both patches and corridors. This can be demonstrated simply by the fact that more than 70% of species present in fragments also appear in corridors. Owing to its effective structural and functional connectivity, the corridor-fragment system studied in this work should be seen as a potentially usable environment for local species, whether allowing their movement from fragments to corridors or from corridors to fragments, thus helping ensure the maintenance of species in such a landscape.
Acknowledgments
We would like to thank the Coordenação de Aperfeiçoamento de Pessoal de Ensino Superior (CAPES) for its support; the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) for the license to collect specimens; Toby A. Gardner for helping to analyze data; and everyone who helped us during the fieldwork, especially Carlos H, Jacinto, Marina S. Pereira, and Adriana L. Gouveia. This study was funded by the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG).
References
Aars, J. & R.A. Ims. 1999. The effect of habitat corridors on rates of transfer and interbreeding between vole demes. Ecology 80: 1648-1655. [ Links ]
Alho, C.J.R. 1981. Small mammal populations of Brazilian Cerrado: the dependence of abundance and diversity on habitat complexity. Rev. Bras. Biol. 41: 223-230. [ Links ]
Andreassen, H.P., K. Hertzberg & R.A. Ims. 1998. Spaceuse responses to habitat fragmentation and connectivity in the root vole Microtus oeconomus. Ecology 79: 1223-1235. [ Links ]
Bennett, A.F. 1990. Habitat corridors and the conservation of small mammals in a fragment forest environment. Landscape Ecol. 4: 109-122. [ Links ]
Bennett, A.F., J.Q. Radford & A. Haslem. 2006. Properties of land mosaics: implications for nature conservation in agricultural environments. Biol. Conserv. 133: 250-264. [ Links ]
Bolger, D.T., T.A. Scott & J.T. Rotenberry. 2001. Use of corridor-like landscape structures by birds and small mammal species. Biol. Conserv. 102: 213-224. [ Links ]
Bonvicino, C.R., J.A. Oliveira & P.S. D’Andrea. 2008. Guia dos roedores do Brasil, com chaves para gêneros baseadas em caracteres externos. Rio de Janeiro: Centro Pan-Americano de Febre Aftosa - OPAS/ OMS, Río de Janeiro, Brasil. [ Links ]
Bonvicino, C.R., J.A. Oliveira & R. Gentile. 2010. A new species of Calomys (Rodentia: Sigmodontinae) from Eastern Brazil. Zootaxa 2336: 19-25. [ Links ]
Brooks, T.M., C.G. Mittermeier, R.A. Mittermeier, G.A.B. Fonseca, A.B. Rylands, W.R. Konstant, P. Flick, J. Pilgram, S. Olfield, G. Magin & C. Hilton-Taylor. 2002. Habitat loss and extinction in the hotspots of biodiversity. Conserv. Biol. 16: 909-923. [ Links ]
Castro, G.C. 2004. Análise da estrutura, diversidade florística e variações espaciais do componente arbóreo de corredores de vegetação na região do Alto Rio Grande, MG. Master Thesis, Universidade Federal de Lavras, Lavras, Minas Gerais, Brasil. [ Links ]
Ceballos, G. & P.R. Ehrlich. 2006. Global mammal distributions, biodiversity hotspots, and conservation. P. Natl. Acad. Sci. U.S.A. 103: 19374-19379. [ Links ]
Clarke, K.R. & R.N. Gorley. 2001. PRIMER v-5: User Manual/Tutorial. Primer-E, Plymouth, England. [ Links ]
Colwell, R.K. 2008. EstimateS: Statistical Estimation of Species Richness and Shared Species from samples. version 8.0. University of Connecticut, Connecticut, USA. (also available online: http://viceroy.eeb.uconn. edu/estimates). [ Links ]
Costa, P.L., Y.L.R. Leite, S.L. Mendes & D.A. Ditchfield. 2005. Conservação de mamíferos no Brasil. Megadiversidade 1: 103-112. [ Links ]
Cuarón, A.D. 2000. A global perspective on habitat distubance and tropical rainforest mammals. Conserv. Biol. 14: 1574-1579. [ Links ]
Dantas, A.A.A., LG. Carvalho & E. Ferreira. 2007. Classificação e tendências climáticas em Lavras, MG. Cienc. Agrotec. 31: 1862-1866. [ Links ]
Downes, S.J., K.A. Handasyde & M.A. Elgar. 1997. The use of corridors by mammals in fragmented Australian Eucalypt Forests. Conserv. Biol. 11: 718-726. [ Links ]
Fahrig, L. 2002. Effect of habitat fragmentation on the extinction threshold: a synthesis. Ecol. Appl. 12: 346-353. [ Links ]
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 34: 487-515. [ Links ]
Fahrig, L. & G. Merriam. 1994. Conservation of fragmented populations. Conserv. Biol. 8: 50-59. [ Links ]
Fischer, J. & D.B. Lindenmayer. 2007. Landscape modification and habitat fragmentation: a synthesis. Global Ecol. Biogeogr. 16: 265-280. [ Links ]
Fonseca, M.T. 1997. A estrutura da comunidade de pequenos mamíferos em um fragmento de Mata Atlântica e monocultura de Eucalipto: a importância da matriz de habitat. Master Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brasil. [ Links ]
Gascon, C., T.E. Lovejoy, R.O. Bierregaard, J.R. Malcolm, P.C. Stouffer, H.L. vasconcelos, W.F. Laurance, B. Zimmerman, M. Tocher & S. Borges. 1999. Matrix habitat and species richness in tropical forest remnants. Biol. Conserv. 91: 223-229. [ Links ]
Gibbs, J.P. 2001. Demography versus habitat fragmentation as determinants of genetic variation in wild populations. Biol. Conserv. 100: 15-20. [ Links ]
Haddad, N.M., D.R. Bowne, A. Cunningham, B.J. Danielson, D.J. Levey, S. Sargent & T. Spira. 2003. Corridor use by diverse taxa. Ecology 84: 609-615. [ Links ]
Haddad, N.M. & J.J. Tewksbury. 2006. Impacts of corridors on populations and communities, p. 390-415. In K.R. Crooks & M. Sanjayan (eds.). Connectivity Conservation. Cambridge University, Cambridge, England. [ Links ]
Hobbs, R.J. 1992. The role of corridors in conservation: solution or badwagon? Trends Ecol. Evol. 7: 389-392. [ Links ]
Knaapen, J.P., M. Scheffer & B. Harms. 1992. Estimating habitat isolation in landscape planning. Landscape Urban Plan. 23: 1-16. [ Links ]
Lidicker, W.Z.J. 1999. Responses of mammals to habitat edges: an overview. Landscape Ecol. 14: 333-343. [ Links ]
Malcolm, J.R. 1994. Edge effects in central Amazonian forest fragments. Ecology 75: 2438-2445. [ Links ]
Mech, S.G. & J.G. Hallett. 2001. Evaluating the effectiveness of corridors: a genetic approach. Conserv. Biol. 15: 467-474. [ Links ]
Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A.B. Fonseca & J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858. [ Links ]
Oliveira-Filho, A.T., E. vilela, M.L. Gavilanes & D.A. Carvalho. 1994. Comparison of the woody flora and soils of six areas of montane semideciduous forest in Southern Minas Gerais, Brazil. Edinb. J. Bot. 51: 355-389. [ Links ]
Oliveira, J.A. & C.R. Bonvicino. 2006. Ordem Rodentia, p. 347-406. In N.R. Reis, A.L. Peracchi & I.P. Lima (eds.). Mamíferos do Brasil. UEL, Curitiba, Paraná, Brasil. [ Links ]
Orme, C.D.L., R.G. Davies, M. Burgess, F. Eigenbrod, N. Pickup, v.A. Olson, A.J. Webster, T.S. Ding, P.C. Rasmussen, R.S. Ridgely, A.J. Stattersfield, P.M. Bennett, T.M. Blackburn, K.J. Gaston & I.P.F. Owens. 2005. Global hotspots of species richness are not congruent with endemism or threat. Nature 436: 1016-1019. [ Links ]
Pardini, R. 2004. Effects of forest fragmentation on small mammals in an Atlantic Forest landscape. Biodivers. Conserv. 13: 2567-2586. [ Links ]
Pardini, R., S.M. Souza, R. Braga-Neto & J.P. Metzger. 2005. The role of forest structure, fragment size and corridors in maintaining small mammal abundance and diversity in an Atlantic forest landscape. Biol. Conserv. 124: 253-266. [ Links ]
Pardini, R., D. Faria, G.M. Accacio, R.R. Laps, E. Marianno-Neto, M.L.B. Paciencia, M. Dixo & J. Baumgarten. 2009. The challenge of maintaining biodiversity in the Atlantic forest: a multi-taxa con- servation assessment of specialist and generalist species in an agro-forestrymosaic in southern Bahia. Biol. Conserv. 142: 1178-1190. [ Links ]
Pardini, R., A.A. Bueno, T.A. Gardner, P.I. Prado & J.P. Metzger. 2010. Beyond the Fragmentation Threshold Hypothesis: Regime Shifts in Biodiversity Across Fragmented Landscapes. Plos One 5: e13666. [ Links ]
Passamani, M. & D. Ribeiro. 2009. Small mammals in a fragment and adjacent matrix in Southeastern Brazil. Braz. J. Biol. 69: 58-60. [ Links ]
Passamani, M. & F.A.S. Fernandez. 2011a. Abundance and richness of small mammals in fragmented Atlantic Forest of Southeastern Brazil. J. Nat. Hist. 45: 553-565. [ Links ]
Passamani, M. & F.A.S. Fernandez. 2011b. Movements of small mammals among Atlantic Forest fragments in Espírito Santo, Southeastern Brazil. Mammalia 75: 83-86. [ Links ]
Püttker, T., Y. Meyer-Lucht & S. Sommer. 2008. Fragmentation effects on population density of three rodent species in secondary Atlantic Rainforest, Brazil. Stud. Neotrop. Fauna E. 43: 11-18. [ Links ]
Rocha, M.F., M. Passamani & J. Louzada. 2011. A small mammals community in a forest fragment-vegetation corridor-coffee matrix system in the Brazilian Atlantic forest. Plos One 6: e23312. [ Links ]
Rossi, R.v., G.v. Bianconi & W.A. Pedro. 2006. Ordem Didelphimorphia, p. 27-60. In N.R. Reis, A.L. Peracchi & I.P. Lima (eds.). Mamíferos do Brasil. UEL, Curitiba, Paraná, Brasil. [ Links ]
Saunders, D.A., R.J. Hobbs & C.R. Margules. 1991. Biological consequences of ecosystem fragmentation: a review. Conserv. Biol. 5: 18-32. [ Links ]
Statsoft. 2001. Statistica for Windows: computer program manual. Tulsa, Oklahoma, USA. (also available online: http://www.statsoft.com). [ Links ]
Turner, I.M. 1996. Species loss in fragments of tropical rain forest: a review of the evidence. J. Appl. Ecol. 33: 200-209. [ Links ]
Umetsu, F. & R. Pardini. 2007. Small mammals in a mosaic of forest remnants and anthropogenic habitats: evaluating matrix quality in an Atlantic forest landscape. Landscape Ecol. 22: 517- 530. [ Links ]
Vieira, M.V., N. Olifiers, A.C. Delciellos, V.Z. Antunes, L.R. Bernardo, C.E.v. Grelle & R. Cerqueira. 2009. Land use vs. fragment size and isolation as determinants of small mammal composition and richness in Atlantic Forest remnants. Biol. Conserv. 142: 1191-1200. [ Links ]
Viveiros de Castro, E.B. & F.A.S. Fernandez. 2004. Determinants of differential extinction vulnerabilities of small mammals in Atlantic Forest fragments in Brazil. Biol. Conserv. 119: 73-80. [ Links ]
Wegner, J.F. & G. Merriam. 1979. Movements by birds and small mammals between a wood and adjoining farmland habitats. J. Appl. Ecol. 16: 349-357. [ Links ]
Weksler, M., A.R. Percequillo & R.S. voss. 2006. Ten new genera of Oryzomyini rodents (Cricetidae: Simodontinae). Am. Mus. Novit. 3537: 1-29.
[ Links ]
*Correspondencia
Andréa O. Mesquita: Pós-Graduação em Ecologia Aplicada, Universidade Federal de Lavras, Campus Universitário - Lavras, MG - 37200-000, Brazil. andreamesquita.bio@gmail.com
Marcelo Passamani: Universidade Federal de Lavras, Setor de Ecologia, DBI, Campus Universitário - Lavras, MG - 37200-000, Brazil/Pós-Graduação em Ecologia Aplicada, Universidade Federal de Lavras, Campus Universitário - Lavras, MG - 37200-000, Brazil. mpassamani@ufla.br
1. Pós-Graduação em Ecologia Aplicada, Universidade Federal de Lavras, Campus Universitário - Lavras, MG - 37200-000, Brazil; andreamesquita.bio@gmail.com
2. Universidade Federal de Lavras, Setor de Ecologia, DBI, Campus Universitário - Lavras, MG - 37200-000, Brazil; mpassamani@ufla.br
Received 12-VIII-2011. Corrected 10-III-2012. Accepted 12-IV-2012.


{kind=link}
{kind=link}
{kind=link}