










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Estructura poblacional de Polythore gigantea (Odonata: Polythoridae) en sistemas lóticos con diferentes estados de conservación en Antioquia, Colombia
*Dirección para correspondencia
Abstract
Population structure of Polythore gigantea (Odonata: Polythoridae) in lotic systems with different conservation states in Antioquia-Colombia. The knowledge about population structure and dynamics of some neotropical species, especially those living in lotic systems is still barely studied. This study had the aim to assess if the conservation status of some lotic systems, is related to some demographic variables of P. gigantea, so this may be used as a model for ecological monitoring. For this, we evaluated the population structure of P. gigantea three times per month (almost one sampling event every eight days) in four streams of the state of Antioquia, Colombia, from March-June 2009. The specimens were collected using entomological nets along a transect of 200m in the littoral zone of each stream. The insects were marked on the wings and the population size was estimated with the mark-recapture method. Our results showed that the largest population size was recorded for the stream “La Catedral” with aprox 299 individuals, followed by the stream “La Doctora” with 218 individuals. Nevertheless, no significant differences in population size among the evaluated streams were found; and no statistical relationships were found between vegetation variables and the population size of P. gigantea. However, taking into account the limited dispersal capacity of P. gigantea, its survival in the studied streams was considered to be at risk, due to the continuous modification of large riparian forest areas, which cause the increase of forest patches, with different levels of interconnection, and hinder long-term permanence of populations.
Key words: Polythore gigantea, Odonata, mark-recapture, population size, sex ratio, life expectancy.
Resumen
Evaluar si el estado de conservación de algunos sistemas lóticos, se relaciona con algunas variables demográficas de P. gigantea puede ser un modelo útil para el monitoreo ecológico. Se evaluó la estructura poblacional de P. gigantea en cuatro sistemas lóticos del Departamento de Antioquia, Colombia. Para cada quebrada, se emplearon tres eventos mensuales, repartidos en un evento cada ocho días, es decir, un total de 12 eventos para los cuatro sistemas lóticos, de marzo - junio de 2009. La metodología empleada fue marcaje y recaptura a lo largo de un transecto de 200m en la zona litoral de cada quebrada. El mayor tamaño poblacional promedio estimado por medio de Jolly, fue para la quebrada La Catedral con 299.4 individuos, seguida de la quebrada la Doctora con 218.3 individuos; sin embargo, no se encontraron diferencias significativas en el tamaño poblacional entre quebradas. No se encontraron relaciones estadísticas entre algunas variables estructurales de la vegetación con el tamaño poblacional de la especie en las cuatro quebradas. No obstante, teniendo en cuenta la limitada capacidad de dispersión de P. gigantea, esta constituye una especie con alto riesgo supervivencia en las quebradas evaluadas, a causa de la permanente modificación de extensas áreas de bosque por parte de los habitantes, que ocasionan la formación de numerosos fragmentos boscosos con diferente grado de interconexión y dificultan la permanencia a largo plazo de las poblaciones.
Palabras clave: Polythore gigantea, Odonata, marcaje- recaptura, tamaño poblacional, proporción sexual, esperanza de vida.
Las comunidades bióticas tanto de ecosistemas terrestres como acuáticos, son muy sensibles a las alteraciones del medio, sean éstas de origen natural o antrópico. En ecosistemas acuáticos; la cantidad de materia orgánica, el estado trófico del agua, la productividad, la sedimentación, el uso de fertilizantes y la contaminación química, entre otros procesos, determinan la composición, abundancia y diversidad de las comunidades acuáticas asociadas (Hawking & New 2002). La alteración de cualquiera de estos procesos, produce cambios en las características físicas y químicas del agua, y en consecuencia, cambios drásticos en las características ecológicas de las mismas. En ecosistemas acuáticos, lóticos en particular, la deforestación, la destrucción del hábitat y el grado de intervención sobre las comunidades vegetales ribereñas, alteran las condiciones microclimáticas específicas de los cuerpos de agua, y por lo tanto, conllevan también, cambios en la composición, abundancia relativa y diversidad de las especies de este tipo de comunidades. Esta alteración y grado de intervención de la vegetación ribereña en ecosistemas acuáticos lóticos, pueden tener efectos posteriores drásticos sobre todo el ecosistema (Thorp & Covich 2001).
En el trópico, el escaso conocimiento de las características demográficas y poblacionales de las especies de ecosistemas acuáticos lóticos, ha obstaculizado el establecimiento de prioridades y la concentración de esfuerzos en acciones prácticas para su conservación (Dalzochio et al. 2011). En tal sentido, en los últimos años, el uso de las comunidades de insectos y en particular de algunas especies de odonatos, como herramienta para evaluar la salud de los ecosistemas ha ido en aumento (Kalkman et al. 2008). Una de las ventajas de este grupo, es que los estados inmaduros de los odonatos pasan la mayor parte de su ciclo de vida en el agua, y que tanto larvas como adultos son muy sensibles a las alteraciones del medio; asimismo, el estudio de sus poblaciones y la estabilidad de las mismas, permiten evaluar también, la estabilidad del ambiente (Von Ellenrieder 2000, Osborn 2005). Otras características como la facilidad de captura, la funcionalidad dentro de los ecosistemas, la sensibilidad a los cambios ambientales, la fidelidad ecológica y sus poblaciones relativamente estables, hacen de algunas especies de odonatos, un grupo de mucha utilidad en el monitoreo de las alteraciones y degradación los ecosistemas ribereños de ríos y quebradas.
Sin embargo, pese a la importancia de las poblaciones de odonatos como indicadores de alteración de ecosistemas y su utilidad en el monitoreo ambiental, el conocimiento sobre la dinámica y estructura poblacional de las especies neotropicales en ecosistemas acuáticos lóticos es aún muy incipiente (Sánchez & Realpe 2010). Actualmente, la mayoría de estudios de los odonatos y su papel como indicadores, se han desarrollado en la zona templada y principalmente con especies en ecosistemas lénticos (Corbet 1999). Si los estudios de campo son intensivos, los datos no sólo permitirán determinar el papel de las especies en su ambiente, sino que también, podrían permitir la elaboración de modelos para el análisis de poblaciones localizadas de la misma especie, bajo condiciones ambientales diferentes. Asimismo, los estudios comparativos entre dos o más poblaciones son especialmente valiosos porque el resultado puede ilustrar las diferencias y la flexibilidad en los patrones de comportamiento de una especie en diferentes ambientes (Garrison & Hafernik 1981).
Polythore gigantea (Selys 1853) es una especie del orden Odonata con distribución neotropical que al igual que muchas especies de la familia Polythoridae, habita zonas ribereñas de ecosistemas lóticos, caracterizados por la abundancia y diversidad de especies vegetales, que crean ambientes con condiciones microclimáticas específicas y aportan gran cantidad de hojarasca, que se acumula en el agua y origina sustratos adecuados para la oviposición; desarrollo y crecimiento de las larvas en este tipo de hábitats. Gracias a que algunas especies de odonatos, responden a alteraciones y destrucción de este tipo de habitas, han permitido considerarlas como un grupo de insectos con alto potencial como indicadores del estado de salud de los ecosistemas acuáticos (Samways 1993, Samway & Caldwell 1996).
El presente estudio, pretende determinar si características ambientales, físicas y estructurales tanto de las quebradas, como de los bosques de ribera, guardan relación con parámetros poblacionales como tamaño poblacional, proporción de sexos, porcentaje de supervivencia y esperanza de vida de P. gigantea, estimados a partir del método de captura-marcaje y recaptura en ecosistemas acuáticos lóticos tropicales.
Materiales y métodos
Área de estudio: A través de visitas previas y una vez confirmada la presencia de la especie, se seleccionaron sitios de muestreo en las quebradas “La Doctora” y “La Romera” del municipio de Sabaneta; y el de “La Miel” y “La Catedral” en el municipio de Envigado, pertenecientes al área metropolitana del Valle de Aburrá. Cada quebrada, en diferente estado de conservación, fue caracterizada según la composición, el porcentaje de cobertura, el DAP y el área basal de la vegetación. Los eventos de captura-recaptura para todas las quebradas se realizaron en el periodo de lluvia, entre los meses de marzo-junio 2009.
La quebrada “La Doctora”, se encuentra a 1 759msnm (06°07’ N - 75°36’ W), tiene un ancho promedio de 3.6m y está caracterizada por pasturas con dedicación ganadera, predominio de bosque natural intervenido en la parte alta, y reforestación con plantaciones de Pinus patula (Schl et Cham) y la ribera rodeada de helechos. Por su parte, “La Romera” se encuentra a 1 930msnm (06°08’ N - 75°36’ W), y un ancho promedio 4.4m, con predominio de bambú (Bambusa sp.) en la margen izquierda y un bosque natural de aproximadamente ocho años de regeneración en la margen derecha, abundancia de helechos y musgos en su parte litoral. La quebrada “La Miel”, ubicada a 1 987msnm (06°08’ N - 75° 34’ W), y ancho promedio 7.6m, tiene predominio de zonas de bosque secundario y áreas desforestadas sometidas a procesos de tala y quema, con dedicación agrícola. Finalmente, la quebrada “La Catedral”, ubicada a 1 720msnm (06°10’ N - 75°35’ W), con un ancho promedio de 3m, tiene predominio de bosques naturales en proceso avanzado de más de 10 años de regeneración, rodeada de una gran masa de bosques de helechos y musgos en la zona litoral de la parte alta y la parte baja con suelos de vocación agrícola.
Tamaño poblacional, supervivencia, esperanza de vida y porcentaje de recaptura: Para el estudio de las poblaciones se estableció un transecto de 200m de longitud, paralelo al litoral de cada una de las quebradas. Se recolectaron adultos de P. gigantea, usando el método de captura manual con red entomológica; los muestreos se efectuaron en un período continuo de ocho horas. Cada evento de muestreo se realizó durante dos días consecutivos, y consistía en capturar, marcar y recapturar el mayor número de ejemplares posibles. Para cada sistema, se emplearon cuatro eventos mensuales, repartidos en un evento cada ocho días. En los cuatro sistemas lóticos seleccionados, el primer evento fue de captura y marcaje únicamente; por lo tanto, se realizó un total 12 eventos de captura y recaptura, tres por cada sistema.
Proporción de sexos y estructura de edades: El marcaje se hizo sobre las alas, y se utilizó marcadores de punta delgada con tinta indeleble de color rojo. A cada ejemplar capturado, se le asignó un número consecutivo y sobre una planilla de registros, se anotaron los datos correspondientes al nombre de la quebrada, fecha de captura, hora, sexo y edad. Se consideraron tres edades: tenerales (individuos recién emergidos), jóvenes y adultos maduros, los patrones de coloración alar y corporal se usaron como criterio de separación. Se tuvo la precaución de no lastimar los individuos al momento de marcar y que la marca no interfiriera con su comportamiento.
Categorización del estado de conservación de las quebradas y su relación con el tamaño poblacional: La caracterización y evaluación del estado de conservación de las quebradas, se determinó usando la metodología de Barbour et al. (1999), que consiste en asignar valores entre 0-20 a variables cualitativas físicas y estructurales de las quebradas. Las variables que se tuvieron en cuenta para el presente trabajo fueron: cobertura vegetal, disponibilidad de sustrato epifaunal, presencia de rocas en el fondo de la quebrada, velocidad de la corriente, profundidad de la quebrada, depósito de sedimentos, estado del flujo de canal, alteración de canal por factores antrópicos, frecuencia de curvas, probabilidad de erosión de la zona litoral, cobertura del bosque de ribera y altura de la vegetación que protege la zona ribereña. Una vez asignado el valor a cada una de las variables, se suman todos los valores y se dividen por el número de variables evaluadas. En cada quebrada se realizaron cuatro observaciones y el valor de cada observación fue promediado.
Se trazaron al azar cuatro transectos de 25x4m, dispuestos de dos en cada uno de las márgenes de la quebrada. En cada uno se midieron cinco variables: cobertura del dosel, número árboles con un diámetro a la altura del pecho (DAP) mayor a 2cm, altura de la vegetación, distribución de las clases diamétricas y área basal. Para la primera variable se usó un densiómetro que mide la cantidad de luz que penetra el dosel del bosque, de acuerdo con este criterio, se definieron tres categorías: 1. Bosques abiertos, cuando el dosel de los árboles obstruye la penetración de luz entre el 10-39%; 2. Bosques moderadamente cerrados, cuando la obstrucción es del 40-69% y 3. Bosques cerrados, cuando la obstrucción es del 70-100%. El análisis de la distribución de clases diamétricas, permite evaluar el estado ecológico y de conservación del bosque, si la proporción de árboles con diámetros pequeños es mayor, indica un bosque joven en estado reciente de recuperación y si por el contrario, la proporción de árboles con diámetros grandes es mayor, indican un bosque ya maduro o de más años de recuperación. El área basal, es una medida directa de la densidad, definida como la suma por unidad de superficie de los troncos de los árboles en su DAP; cuanta mayor área basal, mayor es la densidad e indica un bosque maduro en mejor estado de conservación.
El tamaño poblacional de adultos se determinó con los estimadores de Jolly-Seber y Manly-Parr, a partir de los cuales, se estimaron también la supervivencia y la esperanza de vida (Schwarz et al. 1999). Los parámetros correspondientes a la estructura de edad y la proporción de sexos de la población, se registraron en el momento de la captura y, usando tablas de contingencia, se buscaron relaciones entre las variables y los eventos de captura. Para evaluar si existía alguna relación entre las variables de estructura de edad y proporción de sexos, con los eventos de captura se realizaron tablas de contingencia.
Con un análisis de varianza de una vía (ANOVA), se compararon los tamaños poblacionales estimados con cada uno de los métodos, de igual manera, se compararon la probabilidad de supervivencia entre eventos de muestreo y entre quebradas. Estos análisis se hicieron una vez comprobado el ajuste a la normalidad de los datos con la prueba de Shapiro Wilk usando el programa STATGRAGHICS (Statpoint 2006). Por otro lado, para determinar si existía algún efecto de las quebradas sobre los porcentajes de recaptura y probabilidad de supervivencia, se utilizó un Análisis de Varianza de dos vías.
El promedio de supervivencia diaria y la esperanza de vida en días, se calcularon como lo describen Garrison & Hafernik (1981). La tasa de supervivencia diaria se convirtió en Esperanza de Vida Diaria por el método de Cook et al. 1967 (Garrison & Hafernik 1981, Córdoba 1994).
Por último, para evaluar el grado de asociación entre algunas variables físicas y estructurales de la vegetación y la variación en el tamaño poblacional de P. gigantea, se hizo un análisis de componentes principales (ACP) y se evaluó el coeficiente de correlación de Pearson, con el software Statgraphics Centurion XV.
Resultados
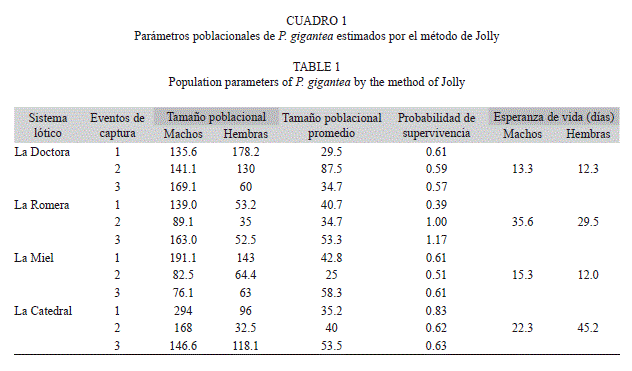
Tamaño poblacional, supervivencia, esperanza de vida y porcentaje de recaptura: El mayor tamaño poblacional promedio estimado por medio de Jolly fue para la quebrada La Catedral con 299.4 individuos, seguida de la quebrada la Doctora con 218.3 individuos, mientras que las quebradas que presentaron los menores tamaños poblacionales fueron La Miel y La Romera con 190.2 y 179.3 individuos, respectivamente; no obstante, al comparar la estimación del tamaño poblacional arrojada por el método de Jolly para cada una de las quebradas, se encontró que no existen diferencias estadísticas significativas (F=3.12, p=0.08, n=12) (Cuadro 1).
Al analizar el tamaño poblacional estimado por Jolly para cada sexo, se observa que en los machos este valor fue siempre superior en todas las quebradas (Cuadro 1). Por otro lado, al comparar el tamaño poblacional estimado para cada sexo en las cuatro quebradas, se observó que no existen diferencias significativas, machos (n=3, F=1.53, p=0.28) y hembras (n=3, F=1.53, p=0.27).
En cuanto a las probabilidades de supervivencia diaria obtenidas por el método de Jolly, se observa que en algunos eventos de captura en la quebrada La Romera, la probabilidad supera el valor de uno (Cuadro 1), lo que indica que hubo valores atípicos en el muestreo. Sin embargo, comparando los valores de supervivencia para las cuatro quebradas, no se encontraron diferencias significativas (n=3, F=0.59, p=0.63).
Los valores de esperanza de vida obtenidos por medio del método Jolly, muestran que en los machos este valor es mayor. Existen diferencias estadísticas entre la esperanza de vida registrada para las cuatro quebradas (F=64.57, p=<0.001), de igual forma se puede observar que en la quebrada La Romera, la esperanza de vida de los individuos (independiente del sexo) es superior a la reportada para los demás ecosistemas lóticos (Cuadro 1), ésto es quizás consecuencia del valor tan alto de la estima de supervivencia diaria.
El promedio de esperanza de vida encontrada para P. gigantea en el presente trabajo, fue de 20.8 días, sin embargo, la esperanza de vida de los machos fue levemente superior al de las hembras con un promedio de 21.55 días.
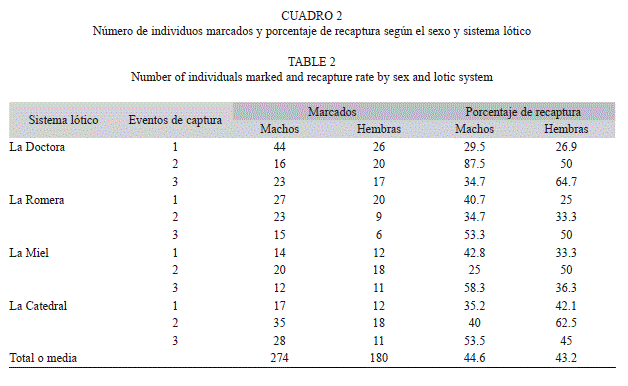
El mayor número de capturas se registró en la quebrada La Doctora con 179 individuos y el menor con 111 en la quebrada La Miel. En general, para las cuatro quebradas las capturas superaron los 100 individuos mientras que las recapturas fluctuaron entre 35-63 individuos, correspondientes a las quebradas La Miel y La Doctora, respectivamente. Al realizar una comparación del porcentaje de recaptura de cada sexo en todos los sistemas lóticos, se evidencia que no existen diferencias estadísticamente significativas (n=6, F=5.25, p=0.08), al igual que al comparar dicho porcentaje entre los sistemas lóticos (n=6, F=0.51, p=0.67); sin embargo, la quebrada “La Doctora” presentó mayor variación en los porcentajes (Cuadro 2).
Proporción de sexos y estructura de edades: La proporción de sexos se mantuvo constante en los cuatro sistemas lóticos, en donde el número de machos siempre fue mayor que el número de hembras. Tal resultado fue más marcado en las quebradas “La Doctora” y “La Catedral” donde se registraron 110 machos/69 hembras y 105 machos/73 hembras, respectivamente. Los resultados muestran que la proporción de machos y hembras no varía significativamente entre las cuatro quebradas (X2=2.24, p=0.52).
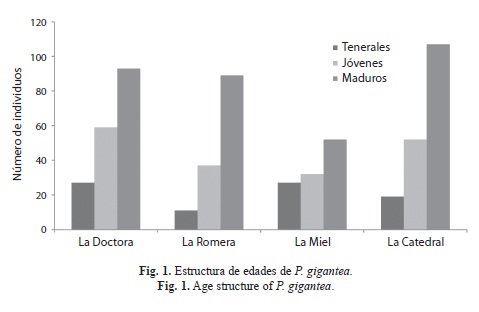
Al comparar el número de individuos registrados en cada categoría de edad, para las cuatro quebradas, se observaron diferencias significativas (X2=19.2, p=0.003). La estructura de edad, siempre fue diferente en cada uno de los sistemas lóticos. En la quebrada “La Miel” a medida que aumenta el número de individuos registrados en la categoría de teneral, el número de adultos maduros decrece, mientras que en la quebrada “La Catedral” este comportamiento se invierte (Fig. 1).
Categorización del estado de conservación de las quebradas y su relación con el tamaño poblacional: Los resultados del análisis del estado de conservación de las quebradas, utilizando la metodología propuestas por Barbour et al (1999), agrupan a los ecosistemas lóticos en dos grupos, la quebrada La Miel y La Catedral se categorizan en estado subóptimo, mientras que La Romera y La Doctora en estado óptimo.
Las comparaciones de las variables estructurales entre los cuatro sistemas lóticos, no presentaron diferencias significativas para las clases diamétricas (n=4, F=1.31, p=0.26), ni entre las alturas (n=4, F=0.22, p=0.87); mientras que para la variable área basal en m2 sí se encontraron diferencias marginalmente significativas (n=4, F=3.84, p=0.07).
Por otro lado, el análisis de las distribuciones diamétricas nos permitió inferir el estado demográfico de las masas de bosque y sus posibles problemas de conservación. Las cuatro quebradas presentan el mismo patrón, en donde la mayoría de los individuos se encuentran agrupados en la clase diamétrica 1-3cm de DAP (38.9% La Miel, 51% La Romera, 46.9% La Doctora y 34.3%), por lo tanto, presentan una distribución de tamaños asimétrica positiva; es decir, con una mayor proporción de las clases de tamaño pequeño (que se supone corresponden a los árboles más jóvenes).
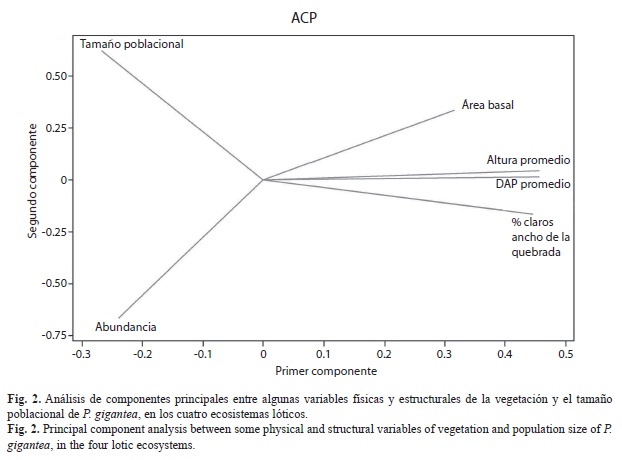
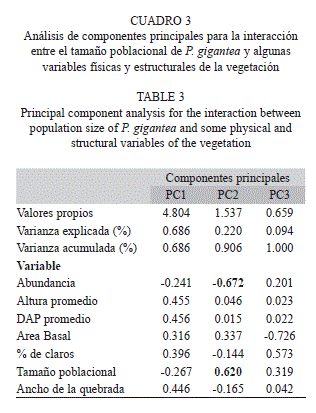
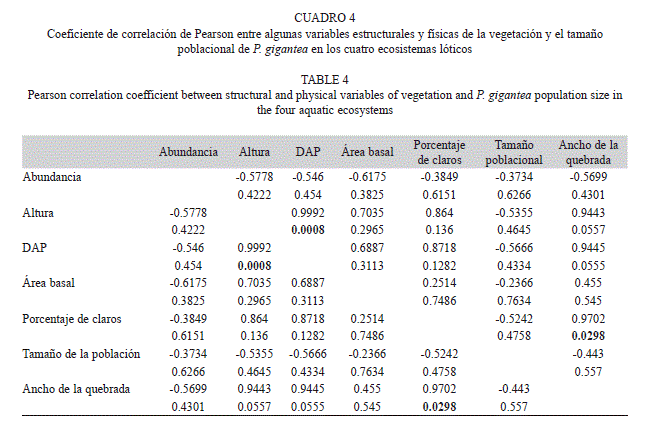
El análisis de componentes principales entre el tamaño poblacional de P. gigantea y algunas variables físicas y estructurales de la vegetación, con solo dos componentes alcanzó un valor apreciable de la varianza acumulada de 90%; sin embargo, del análisis gráfico se puede inferir que no hubo relaciones interactivas de ninguna variable con el tamaño poblacional de la especie (Fig. 2). En el segundo componente, quedaron incluidas las variables de abundancia y el tamaño poblacional (Cuadro 3). Para corroborar el planteamiento anterior se realizó un análisis del coeficiente de correlación de Pearson y permitió evidenciar la inexistencia de dichas relaciones. No obstante, se observa una relación positiva entre el DAP y la altura de la vegetación (r=0.99, p=0.001) y entre el ancho de la quebrada y el porcentaje de claros (r=0.97, p=0.029) (Cuadro 4).
Discusión
Una de las ventajas del marcaje de animales silvestres, es que su longevidad puede ser medida por múltiples eventos de recaptura. No obstante, según Cordero & Stoks (2008) en los estudios de marcaje y recaptura se tiende a subestimar la longevidad real de los adultos por las siguientes razones: la fecha de marcaje no suele ser la fecha de emergencia, es poco probable que el último avistamiento sea la fecha de la muerte y por último, los animales pueden emigrar y por lo tanto en gran parte de sus vidas son imposibles de atrapar; aún con estas limitaciones, el marcaje es considerada la mejor manera de estimar los parámetros demográficos más relevantes en odonatos adultos (Anholt et al. 2001). Posiblemente durante el estado adulto, las hembras sufren mayores tasas de mortalidad y dispersión, así lo evidencia la menor longevidad y porcentaje de recapturas registradas en el presente trabajo; Cordero 1988, lo reporta también para poblaciones de Lestes viridis (Vander Linden 1825) en España. Al comparar la Esperanza de vida de los machos de diferentes especies, algunas con distribución neotropical como: Hetaerina cruentata (Rambur 1842) (Calopterygidae), Palaemnema desiderata (Selys 1886) y P. paulitoyaca (Calvert 1931) (Platystictidae), Polythore procera (Polythoridae), se concluye que H. cruentata y P. procera son las representantes neotropicales de mayor longevidad con 44.9 y 43.6 días, respectivamente (Sánchez & Realpe 2010).
Debido a que en la mayoría de especies de odonatos, los machos son marcadamente territoriales y las hembras visitan esporádicamente los cuerpos de agua, se espera, que tanto las capturas como las recapturas de machos, sean mayores que las de las hembras, tal como se ha corroborado por los resultados del presente trabajo. Autores como Corbet (1999), han señalado que la conducta territorial en libélulas, es característica particular de los machos, y en general éstos establecen sus territorios en perchas cerca de los sitios de apareamiento. En el caso de P. gigantea, los machos se observaron la mayor parte del tiempo posados en la vegetación adyacente a la zona ribereña de estos ecosistemas lóticos. De manera similar, Garrison 1978, Garrison & Hafernik 1981, Córdoba 1993, Angelibert & Giani 2003, lo han reportado también para otras especies de odonatos y Sánchez & Realpe (2010), en particular, lo reportan para Polythore procera (Selys 1869).
La dispersión es un fenómeno ecológico fundamental en la biología de una población, tiene un efecto potencialmente profundo en la dinámica de las poblaciones, el nivel de flujo génico y los procesos de adaptación local, especialmente cuando la población ocupa un hábitat irregular (Torralba & Ocharan 2006). Sistemas lénticos rodeados de paisaje terrestre son un ejemplo de distribución desigual de las condiciones físicas y pueden constituir “islas” para algunas especies de odonatos (Angelibert & Giani 2003). La sensibilidad a las condiciones climáticas, el tamaño y comportamiento de P. gigantea, posiblemente estarían limitando su capacidad y distancia de dispersión, debido a que en ningún periodo de recolecta se observaron individuos desplazándose de un fragmento de bosque a otro por muy cercanos que estuviesen (Fincke 2007, Sánchez & Realpe 2010). Por lo tanto, y de acuerdo con lo observado, se puede considerar que esta especie mantiene poblaciones muy locales y asociadas a pequeños relictos de bosque en mediano o buen estado de conservación.
La proporción de sexos de P. gigantea, en los cuatro sistemas fue similar, siempre los machos en mayor proporción que las hembras. Este resultado puede ser explicado si se tiene en cuenta que los muestreos se realizaron en cercanías a la zona litoral de los sistemas lóticos y las hembras de muchas especies de odonatos maduran los huevos lejos del agua y sólo se acercan para aparearse y realizar la puesta (Corbet 1999).
En general, se asume que la mayoría de las hembras consigue aparearse, dado que la proporción de sexos en las cercanías de las masas de agua está muy sesgada hacia los machos (Cordero 1994). Algunos autores han señalado que este sesgo debe ser considerado un artefacto debido al comportamiento más críptico, patrones de coloración de las hembras y uso diferencial del hábitat. De este modo, la probabilidad de recapturar hembras es típicamente menor a la de los machos (Garrison & Hafernik 1981, Stoks 2001). Por otro lado, Cordero & Stoks 2008, plantean otras hipótesis que tratan de esclarecer el comportamiento de la proporción de sexos en odonatos, entre ellas se destacan, que puede haber un sesgo hacia los machos durante la emergencia, que los machos y las hembras pueden no emerger de manera sincrónica, las hembras tienen un periodo más largo de maduración, las hembras inmaduras tienen mayores tasas de mortalidad y por último, que las hembras maduras tienen menores probabilidades de supervivencia. De acuerdo con los resultados, esta última hipótesis no se cumple para P. gigantea dado que, en todos los sistemas lóticos evaluados, los valores de la probabilidad de supervivencia entre sexos, no mostraron diferencias estadísticamente significativas. Incluso en la quebrada La Catedral la probabilidad de supervivencia de las hembras fue levemente superior. El mayor costo energético de los machos en la defensa del territorio y consecución de las hembras para aparear, puede ser un factor determinante para la disminución de la probabilidad de supervivencia en relación con la de las hembras (Marden & Waage 1990).
Aunque el número de individuos de la categoría adulto maduro fue dominante en todas las quebradas, en “La Miel” en particular, la cantidad de tenerales fue alta si se compara con el resto de las quebradas; a su vez, el número de adultos maduros también fue menor. Dicho resultado, se debió quizás a que la especie en esta quebrada, se encontraba en un posible período de emergencia, lo que explicaría la mayor proporción de tenerales. La dominancia de adultos maduros en la mayoría de los eventos de captura, encontrada en el presente trabajo, debe estar altamente relacionada con la captura de hembras tenerales y jóvenes en mayor proporción, sumado también a la incapacidad de los machos tenerales y jóvenes de establecer territorios con fines reproductivos porque aún no han alcanzado la madurez sexual. Otro factor que quizás puede influir en los resultados, es que en las hembras de odonatos la madurez sexual ocurre más tardíamente que en los machos (Anholt 1991), debido en gran parte a la producción masiva de huevos (Corbet 1999).
De acuerdo con los resultados, las diferencias en el número de individuos registrados en cada categoría de edad, también pudieron estar afectadas por los patrones de distribución espacial de la especie. Kadoya et al. (2004) observaron que los individuos jóvenes y tenerales de algunos odonatos, tienden a ubicarse en zonas con una densa vegetación como medida de protección contra depredadores. Por lo tanto, la captura de estos ejemplares es menos frecuente.
Las diferencias estadísticas registradas en la variable área basal, podrían ser un indicador de que en las quebradas La Romera y La Miel los individuos vegetales registrados presentan una mayor capacidad para llenar el área superficial o espacio disponible en el bosque (Enquist & Niklas 2001), lo que a su vez refleja que el grado de regeneración natural es más rápido en dichos ecosistemas. Por lo tanto, si se diseña y ejecuta rápidamente un programa de recuperación de los bosques ribereños para mejorar el estado actual de conservación de la zona litoral de estas quebradas, se podría garantizar el incremento y mantenimiento de las poblaciones de P. gigantea.
Nuestros resultados sugieren que el estado actual de intervención de las quebradas La Catedral y La Miel, que se categorizaron en estado subóptimo mediante la metodología de Barbour et al. (1999), no fueron lo suficientemente fuertes para afectar de forma considerable el tamaño poblacional de P. gigantea. De hecho estas dos quebradas presentaron los valores más altos de tamaño poblacional según el método de Jolly. Posiblemente, la acción de algunas variables físico-químicas o la disponibilidad y cobertura de microhábitas de oviposición, determinan de manera más efectiva la presencia y abundancia de esta especie en los ecosistemas evaluados.
No se encontraron relaciones estadísticas entre algunas variables estructurales de la vegetación con el tamaño poblacional de la especie en las cuatro quebradas; este resultado posiblemente pueda explicarse al tener en cuenta que varias características in situ que aumentan la heterogeneidad en la estructura vegetal no fueron incluidas, entre ellas: hierbas, cultivos, bromelias, volumen de hojarasca y material en descomposición. Yahner (1988) ha propuesto que una mayor heterogeneidad de la estructura vegetal, en este caso favorecida por la abundancia de especies vegetales generalistas que han colonizado porciones del bosque perturbadas por el hombre, puede aumentar la diversidad faunística de las áreas. Sin embargo, el mismo Yahner (1988) y Saunders et al. (1991), consideran que la deforestación puede crear parches de bosque aislados que estarían ocasionando un impacto, y afectando los patrones de dispersión de algunos animales. Teniendo en cuenta la limitada capacidad de dispersión de muchas especies de odonatos y particularmente P. gigantea, esta constituye una especie con alto riesgo de supervivencia en zonas como la del presente estudio, donde la permanente modificación de extensas áreas de bosque por parte los habitantes, han dado como resultado la formación de numerosos parches boscosos con diferente grado de interconexión que finalmente dificultan la permanencia a largo plazo de las poblaciones.
Agradecimientos
A Mariela Lobo y Cornelio Bota, por la ayuda en la fase de campo. A Melissa Sánchez por la colaboración en la fase de análisis de datos. A Adolfo Cordero Rivera de la Universidad de Vigo-España y Alejandro Córdoba Aguilar de la Universidad Autónoma de México (UNAM), por sus valiosas correcciones y sugerencias, a la Universidad Nacional de Colombia sede Medellín y su Museo Entomológico Francisco Luís Gallego.
Referencias
Angelibert, S. & N. Giani. 2003. Dispersal characteristics of three odonate species in a patchy habitat. Ecography 26: 13-20. [ Links ]
Anholt, B.R. 1991. Measuring selection on a population of damselflies with a manipulated phenotype. Evolution 45: 1091-1106. [ Links ]
Anholt, B.R., C. Vorburger & P. Knaus. 2001. Mark recapture estimates of daily survival rates of two damselflies (Coenagrion puella and Ischnura elegans). Can. J. Zool. 79: 895-899. [ Links ]
Barbour, M.T., J. Gerritsen, B.D. Snyder & J.B. Stribling. 1999. Rapid bioassessment protocols for use in streams and wadeable rivers: Periphyton, benthic macroinvertebrates, and fish. U.S Environmental Protection Agency, Office of Water, Washington, D.C., USA. [ Links ]
Cook, L.M., P.P. Brower & H.J. Croze. 1967. The accuracy of a population estimation from multiple recapture data. J. Anim. Ecol. 36: 57-60. [ Links ]
Corbet, P.S. 1999. Dragonflies. Behaviour and ecology of Odonata. Cornell University, Ithaca, New York, USA. [ Links ]
Cordero, A. 1988. Estudio ecológico de una población de Lestes viridis Vander Linden, 1825 (Zygoptera, Lestidae). Limnetica 4: 1-8. [ Links ]
Cordero, A. 1994. Reproductive allocation in differentsized adults Ischnura graellsii (Rambur) (Zygoptera: Coenagrionidae) Odonatologica 23: 271-276. [ Links ]
Cordero, A. & R. Stoks. 2008. Mark–recapture studies and demography. In Dragonflies: Model Organisms for Ecological and Evolutionary Research. Oxfordshire, England. [ Links ]
Córdoba, A. 1993. Population structure in Ischnura denticollis (Burmeister) (Zygoptera: Coenagrionidae). Odonatologica 22: 455-464. [ Links ]
Córdoba, A. 1994. Adult survival and movement in males of the damselfly Hetaerina cruentata (Odonata: Calopterygidae). Fla. Entom. 6: 256-264. [ Links ]
Dalzochio, M.S., J. Martins-Costa & M.A. Uchôa. 2011. Diversity of Odonata (Insecta) in lotic systems from Serra da Bodoquena Mato Grosso do Sul State, Brazil. Rev. Bras. Entomol. 55: 88-94. [ Links ]
Enquist, B.J. & K.J. Niklas. 2001. Invariant scaling relations across tree-dominated communities. Nature 410: 655-660. [ Links ]
Fincke, O.M. 2007. Use of forest and tree species, and dispersal by giant damselflies (Pseudostigmatidae): their prospects in fragmented forests. Odonatologica 14: 121-133. [ Links ]
Garrison, R.W. 1978. A mark-recapture study of imaginal Enallagma cyathigerum (Charpentier) and Argia vivida Hagen (Zygoptera: Coenagrionidae). Odonatologica 7: 223-236. [ Links ]
Garrison, R.W. & J.E. Hafernik Jr. 1981. Population structure of the rare damselfly, Ischnura gemina (Kennedy) (Odonata: Coenagrionidae). Oecologia 48: 377-384. [ Links ]
Hawking, J.H. & T.R. New. 2002. Interpreting dragonfly diversity to aid in conservation assessment: lessons from the Odonata assemblage at Middle Creek, north-eastern Victoria, Australia. J. Insect Conserv. 6: 171-178. [ Links ]
Kadoya, T., S. Suda & I. Washitani. 2004. Dragonfly species richness on man-made ponds: effects of pond size and pond age on newly established assemblages. Ecol. Res. 19: 461467. [ Links ]
Kalkman, V.J., V. Clausnitzer, B. Dijkstra & D.R. Paulson. 2008. Global diversity of dragonflies (Odonata) in freshwater. Hydrobiologia 595: 351-363. [ Links ]
Marden, J.H. & J.K. Waage. 1990. Damselfly territorial contests are zygopteran populations. Odonatologica 4: 353-374. [ Links ]
Osborn, R. 2005. Odonata as indicators of habitat quality at lakes in Louisiana, United States. Odonatologica 34: 259-270. [ Links ]
Samways, M.J. 1993. Dragonflies (Odonata) in taxic overlays and biodiversity conservation, p. 111-123. In K.L. Gaston, T.R. New & M.J. Samways (eds.). Perspectives in insect conservation. Intercept, Andover, United Kingdom. [ Links ]
Samways, M.J., P.M. Caldwell & R. Osborn. 1996. Spatial patterns of dragonflies (Odonata) as indicators for design a conservation pond. Odonatologica 25: 155-166. [ Links ]
Sanchez, M. & M. Realpe. 2010. Population structure of Polythore procera at a Colombian stream (Odonata: Polythoridae). IJO 13: 27-37. [ Links ]
Saunders, D.A., R.J. Hobbs & C.R. Margules. 1991. Biological consequences of ecosystem fragmentation: a review. Cons. Biol. 5: 18-32. [ Links ]
Schwarz, C.J. & G.A. Seber. 1999. Estimating Animal Abundance: Review III. Stadistical Sciences 14: 427-456. [ Links ]
Stoks, R. 2001. Male-biased sex ratios in mature damselfly populations: real or artefact? Ecol. Entomol. 326: 181-187. [ Links ]
Thorp, J.H. & A.P. Covich. 2001. Ecology and classification of North American freshwater invertebrates. Academic, San Diego, California, USA. [ Links ]
Torralba, A. & F.J. Ocharan. 2006. Dispersión y proporción sexual en la emergencia en una población de Sympecma fusca (Odonata, Lestidae) en Huesca (NE de España) Bol. R. Soc. Esp. Hist. Nat. Secc. Biol. 101: 29-36. [ Links ]
Von Ellenrieder, N. 2000. Species composition and temporal variation of odonate assemblages in the subtropical-pampasic ecotone. Odonatologica 29: 17-30. [ Links ]
Yahner, H.R. 1988. Changes in wildlife communities near edges. Cons. Biol. 2: 333-339. [ Links ]
*Correspondencia:
Mariano Altamiranda-S: Universidad Nacional de Colombia sede Medellín, Grupo de investigación en ecología y sistemática de insectos (GIESI), Calle 59A No 63-20, Medellín Colombia. maltamiranda2@gmail.com
Oscar Ortega-M: Universidad Nacional de Colombia sede Medellín, Grupo de investigación en ecología y sistemática de insectos (GIESI), Calle 59A No 63-20, Medellín Colombia. oeortegam@gmail.com
1. Universidad Nacional de Colombia sede Medellín, Grupo de investigación en ecología y sistemática de insectos (GIESI), Calle 59A No 63-20, Medellín Colombia; maltamiranda2@gmail.com, oeortegam@gmail.com
Recibido 06-IX-2011. Corregido 20-I-2012. Aceptado 29-II-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}