










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Mycorrhizal association in gametophytes and sporophytes of the fern Pteris vittata (Pteridaceae) with Glomus intraradices
Alicia E. Martinez1*,3*, Viviana Chiocchio2*,3, Lo Tai Em1, María A. Rodriguez1,3 & Alicia M. Godeas1,3
*Dirección para correspondencia:
Abstract
Ferns, which are usually colonizing different environments and their roots frequently present mycorrhization, have two adult stages in their life cycle, the sporophytic and the gametophytic phase. This paper describes the experimental mycorrhizal association between Pteris vittata leptosporangiate fern and a strain of Glomus intraradices during the life cycle of the fern, from spore germination to the development of a mature sporophyte. The aim of this study was to compare the colonization pattern of in vitro cultures of G. intraradices along the fern life cycle with those found in nature. For this, mature spores were obtained from fertile P. vittata fronds growing in walls of Buenos Aires city, Argentina. Roots were stained and observed under the light microscope for arbuscular mycorrhizal colonization. Approximately, 75 fern spores were cultured in each pot filled with a sterile substrate and G. intraradices (BAFC N° 51.331) as inoculum on the surface. After germination took place, samples were taken every 15 days until the fern cycle was completed. In order to determine colonization dynamics each sample was observed under optical and confocal microscope after staining. Gametophyte was classified as Adiantum type. Male and female gametangia were limited to the lower face, mycorrhizal colonization started when they were differentiated and took place through the rhizoids. Spores and vesicles were not found in this cycle stage. Paris-type mycorrhizal colonization was established in the midrib and in the embrionary foot. It was colonized by external mycelium. When the first root was developed soil inoculum colonized de novo this structure and Arum-type colonization was observed. This study proves that the type of colonization is determined by the structure of the host, not by the fungus. Both the gametophyte and embryo foot have determined growth and Paris-type colonization, while, sporophyte roots have undetermined growth and Arum-type colonization. The structures found in vitro cultures were highly similar to those found under natural conditions. Rev. Biol. Trop. 60 (2): 857-865. Epub 2012 June 01.
Key words: Pteris vittata, Glomus intraradices, Pteridophyta-gametophyte sporophyte, Arum colonization, Paris colonization.
Resumen
Los helechos presentan dos etapas en su ciclo de vida, una fase esporofítica y una gametofítica. Estos por lo general pueden colonizar diferentes ambientes y frecuentemente presentan raíces micorrizadas. Este estudio describe la asociación experimental entre Pteris vittata, un helecho leptosporangiado y una cepa de Glomus intraradices durante el ciclo de vida del helecho, desde la germinación de las esporas hasta el desarrollo del esporofito maduro. El objetivo de este estudio fue comparar los patrones de colonización de G. intraradices a lo largo de todo el ciclo de vida del helecho con los tipos encontrados en la naturaleza. Las esporas maduras fueron obtenidas de frondes fértiles de P. vittata que crecen sobre las paredes de la ciudad de Buenos Aires, Argentina. Las raíces se tiñeron y fueron observadas bajo microscopio óptico para el estudio de la colonización micorrízica. Aproximadamente 75 esporas de helecho se cultivaron en macetas con un sustrato estéril y con un inóculo de G. intraradices (N° 51.331 BAFC) en la superficie. Después de la germinación, se tomaron muestras cada 15 días hasta que se completó el ciclo de vida del helecho. Con el fin de determinar la dinámica de la colonización, cada muestra se observó con el microscopio óptico y el microscopio de confocal luego de la tinción correspondiente. El gametofito fue clasificado como del tipo “Adiantum”. Los gametangios femeninos y masculinos se desarrollaron en la cara inferior del mismo. La micorrización comenzó cuando los gametangios estaban ya diferenciados y la colonización se produjo a través de los rizoides. Las esporas y las vesículas no se encontraron en esta fase del ciclo. La micorrizacion tipo Paris se observó sobre la línea de la nervadura central. El pie del esporofito fue colonizado por el micelio externo. Cuando la raíz se desarrolló, se colonizó “de novo”, y se observó una colonización de tipo Arum. Este estudio demuestra que el tipo de colonización está determinado por la estructura del helecho y no por el hongo. Tanto el gametofito como el pie del embrión tienen crecimiento definido y colonización tipo Paris, mientras que las raíces del esporofito presentan un crecimiento indeterminado y una colonización tipo Arum. Las estructuras que se encontraron bajo cultivo coinciden con las que se encontraron en condiciones naturales.
Palabras clave: Pteris vittata, Glomus intraradices, Pteridophyta-gametofito, esporofito, colonización tipo Arum, colonización tipo Paris.
Over 90% of terrestrial plant groups have some type of symbiosis with soil fungi, and/or some mycorrhizal form. During the Devonic period, the first plants with roots appeared on land, the ferns belonging to Pteridophyta, Filicales groups, still exist today. They are widely distributed, particularly in tropical environments, and many of them have roots colonized by arbuscular mycorrhizal fungi (AM) (Brundrett 2002). Fern species with fine roots and long absorbent hairs sometimes limit mycorrhizal colonization. This facultative association is considered to be a feature of more evolved ferns (Fernandez et al. 2010). Cairney (2000) and Brundrett (2002) suggested that mycorrhizal symbiosis probably enabled plants to colonize land, conferring advantages such as increased fitness and resistance to drought or pathogenic microorganisms under certain conditions (Smith & Read 1997, Daniell et al. 1999, Read 1999).
Preliminary data indicated that the presence of AM fungi at both the sporophyte and the gametophyte stage, stimulates fern growth (Turnau et al. 2005). P. vittata sporophytes grown in controlled conditions showed an increase in fresh and dry weight shoots when grown in contaminated soil (Trotta et al. 2006, Leung et al. 2006). In natural conditions (Zhiwei 2000) from 12 different species of Pteris (P. vittata included) found sporophytic mycorrhization only in P. setulosocostulata.
P. vittata is a widely distributed fern in Buenos Aires city, often growing on the substrate that fills hollows in damp walls. Under these stressful conditions, it was observed that the sporophyte was always colonized by arbuscular mycorrhizal fungi.
P. vittata has a chlorophyllous gametophyte of limited growth, the most vulnerable phase of the life cycle (Zhang et al. 2008). The appearance of AM fungi Glomus intraradices at the gametophyte phase may significantly shorten the period when the small plants are especially susceptible to drought, allowing them to adapt better to the environment (Boullard 1957, 1979, Pirozynski & Malloch 1975).
There are two main morphological types of arbuscular mycorrhizas (AM), the Arum-type and the Paris-type and a continuum between them. They are responsible of the P plant nutrition (van Aarle et al. 2005). In the Arum-type, fungi form intercellular hyphae between the cortical cells and intracellular arbuscules within them. The Paris-type is characterized by extensive intracellular hyphal coils and arbusculate coils in the root cortex. In the Paris-type the intercellular phase of colonization is almost absent. With very few exceptions, members of single plant species formed only one type of colonization. It is often accepted that AM morphology is controlled by plant identity (Smith & Smith 1997). The aim of this study is to determine the relationship between the life cycle of Pteris vittata and the colonization of the arbuscular mycorrhizal fungi G. intraradices.
Materials and methods
Collection of plant material: Approximately 100 specimens of P. vittata L. at different stages of sporophyte development were collected from hollows filled of saline substrate on damp walls of Buenos Aires city, and analysed for root colonization. Sampled sporophytes ranged in height from few millimetres up to 6cm. A number of 20 samples were attached to the gametophyte.
Cultivation of plant material: Spores were obtained from fertile P. vittata fronds and kept in dry, covered containers until they were used. Thirty 50mL pots were filled with a sterile mixture of perlite:peat:soil 5:2:1V/V, and 500mg of G. intraradices inoculum placed on the surface of each pot, containing approximately 75 fern spores on top.
The mycorrhizal strain used in this experiment is a pure culture identified as G. intraradices, originated from a pastureland from Buenos Aires province in Argentina, grown in association with white clover (Trifolium repens). Spores have been preserved as herbarium material (BAFC N° 51.331). The cultura was replicated, and part of it kept in our collection under the name Strain GB1 (Banco de Glomeromycota In vitro (BGIV) http://www.bgiv.com.ar/strains/glomus-intraradices/gb1).
Pots were watered to field capacity and kept in a humid chamber (relative humidity 100%) at 27-30°C and a regime of 16 hours light/8 hours darkness. As from germination, samples were taken every 15 days until the fern cycle was complete, in order to determine colonization dynamics.
Microscopy analyses: The cultured and field material was cleared and stained using the method of Phillips & Hayman (1970) as follows: fresh roots were heated in a 10% KOH solution at 90ºC for 15 minutes, washed in tap water and immersed in 20 vol (H2O2) for 10 minutes until bleached. Then, they were rinsed in tap water to remove H2O2, acidified in 0.1N HCl, and stained with 0.05% Trypan Blue solution for 20 minutes at 90ºC. Dye excess was removed in clear 85% lactic acid. Root segments were mounted on slides in 85% Lactic acid. Observation and microphotography were assessed using a Nikon Opthot-2 microscope fitted with a digital Coolpix 950 camera.
For a more accurate observation of root colonization, confocal microscopy was used. Root samples were fixed at least 12 hours in 50% ethanol. Roots were cleared by heating them in 5% KOH (w/v) at 90ºC, roots were washed with tap water, and then acidified with 0.1N HCl for 5-10 minutes. Roots were stained with 0.01% acid fuchsin (w/v) in solution of acid-glycerin-water (875mL lactic acid, 63mL glycerin, 63mL water) for one hour at 55ºC. Dye excess was removed in 100% glycerin. Observations and microphotography were conducted using an Olympus FV300 confocal scanning HeNe green laser microscope and with excitation at 543nm (Argon laser). The lenses used included Zeiss UplanFI 20x/0.5 and UplanApo 40x/1.0 water immersion objectives. Images were captured and processed using Photoshop v 5.5 software Adobe Systems, San Jose, CA, USA (Peterson et al. 2004).
Results
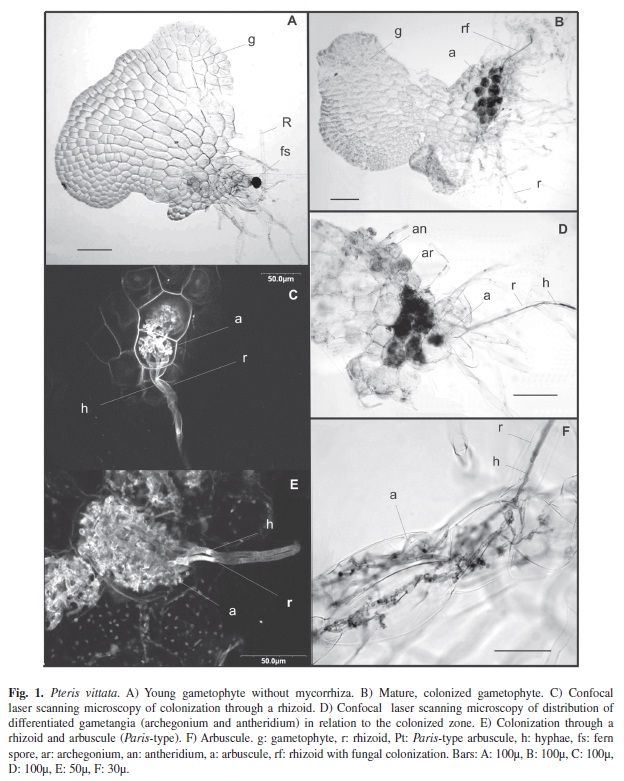
Experimental life cycle of P. vittata showed that spores germinated after eight days and they gave rise to a few cell filaments, and a rhizoid able to differentiate. Gametophyte development (Fig. 1A) was of Adiantum-type (Nayar & Kaur 1969, Martinez 2010), characterized by a meristematic apical cell that divided laterally giving rise to new cells, which formed a notch. The apical cell divided transversally into further meristematic cells, which began to divide rapidly on another plane, producing a central zone made up of several cell layers with rhizoids and two wings. Male and female gametangia were limited to the lower face and to the ventral surface below the notch, mainly between the rhizoids (Fig. 1D). About 75% of the gametophytes examined were colonized by G. intraradices, of sporophytes arising from embryo development.
The first stages of prothallus development lack mycorrhizal colonization even when rhizoids have developed (Fig. 1A). Gametophyte colonization only began when gametangia were differentiated (Fig. 1D). Gametangial differentiation was separated in the time (protandric gametophyte). Colonization took place through the rhizoids (Fig. 1B-F), on which an appresorium formed and penetrated the cell. On reaching the basal cells of the midrib, the hypha grew forming coils and spread through cells (Paris-type), taking up ¾ of the midrib (Fig. 1B, D). The arbuscules, being very small, grew from these coils (Fig. 1C, E, F). There was little, if any, intercellular growth. Arbuscules were ephemeral, and in most cells we observed amorphous, blue-stained material. Spores and vesicles were not found in this phase of the cycle. Gametophyte wings and growth ápex were uncolonized, probably because the wings were unstratified.
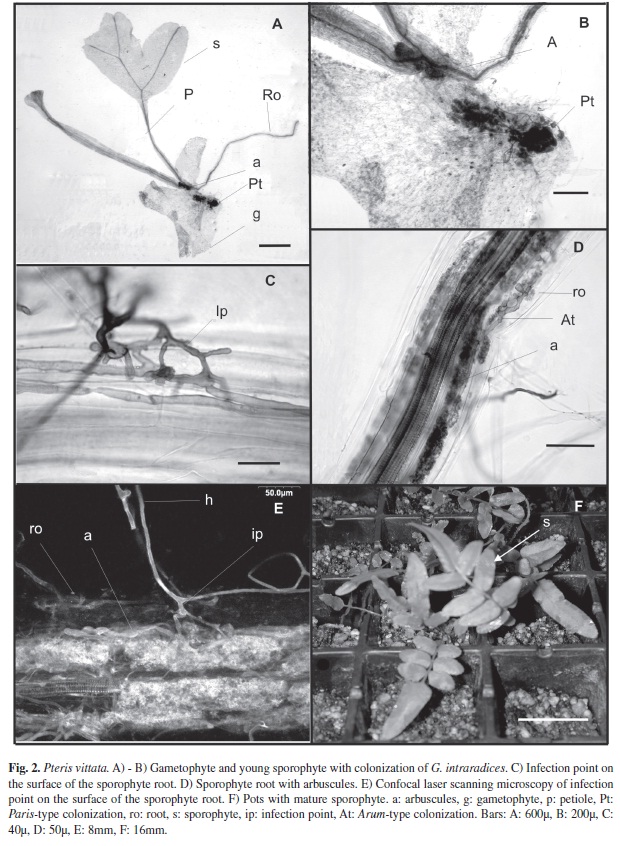
When the embryo developed, the foot that penetrated the gametophyte also showed Paris-type colonization (Fig. 2A). This colonization did not originate from the colonized gametophyte cells, but rather from external mycelium (Fig. 2B), as zones remain separated by uncolonized cells, the foot had determinate growth and enabled the young sporophyte to obtain nutrients from the gametophyte (Fig. 2B).
Right from the start, there was Arum-type colonization of the first sporophyte root (Fig. 2D). Penetration points, intercellular arbuscules and intercellular hyphae could be seen (Fig. 2C-E). There was no colonization through the gametophyte. The sporophyte was colonized by inoculum in the medium (Fig. 1F).
The substrate accompanying the samples was analyzed, resulting in a salt imbalanced substrate with the following characteristics: pH 7.6 (1:2.5 H2O), C.E. dS/m 3.59, Ct(W. Black) 24.98g/kg, Nt (Kjeldahl) 1.92g/kg, P (K y B) 9.3mg/kg, CIC (Ac. NH4 pH7 μDest.) 18.4cmolc/kg, Ca2+ (Ac. NH4 pH7 A.A) 30.5cmolc/kg, Mg2+ (Ac NH4 pH7 A.A) 1.7cmolc/kg, Na+ (Ac NH4 pH7 E/A.A) 4.2 cmolc/kg, K+ (Ac. NH4 pH7 E/A.A) 1.7 cmolc/kg.
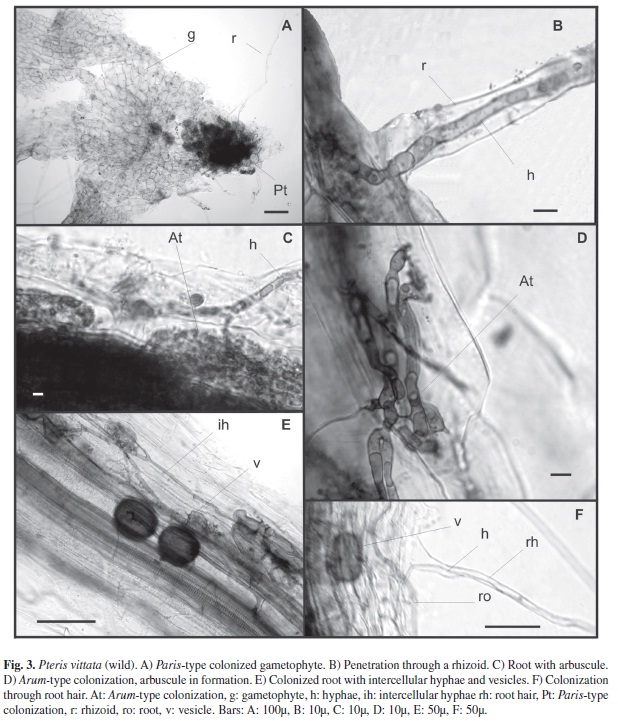
Most gametophytes of P. vittata that developed in hollows in damp walls were mycothallic (Fig. 3A). Fern gametophytes and roots of sporophytes were strongly colonized (Fig. 3A, E). Thin fungal mycelium with extraradical hyphae of 0.8-1.3μm, hyphae developing within the gametophyte often forming complex coils (Paris-type arbuscules) filling the gametophyte cells (Fig. 3A), strongly stained in trypan blue, were visible either in the basal part of the rhizoids or within cells of the Adiantum-type gametophyte, where the rhizoids were initiated. The mycelium was also observed within the elongated part of the rhizoids (Fig. 3B). The fungus was spreading from one cell to another without the development of the intercelular phase (Fig. 3A). The colonization Arum-type of sporophyte roots was observed by the mycelium originating from the extraradical hyphal net (Fig. 3D-F). Vesicles of 3-7μm diameter were found in root cortical cells. (Fig. 3E-F).
From the field material collected, the samples presenting gametophyte (n=20), showed a Paris-type colonization occurring through the rhizoids in contact with the inoculum (Fig. 3A, B). A large amount of extra-radical mycelium can be seen penetrating root hairs in the sporophytes, forming intercellular and intracelular hyphae, arbuscules and vesicles (Fig. 3C-F) similar to the ones found in the sporophytes cultured with G. intraradices.
Discussion
Mycorrhizal dependence of a plant species is one of its constitutive features, allowing plant classification as facultative or obligately mycotrophic. Facultative mycotrophs are those that can grow without mycorrhizae in relatively fertile natural soils. Previous studies, found a low mycorrhization level in Pteridophyte sporophytes, particularly in P. vittata roots (Zhiwei 2000). On the contrary our results showed a high level of fern roots colonization, supporting the previous hypothesis that the amount of available nutrients in the substrate regulates this process (Janos 1993, 2007, Brundrett 2002), leading to the notion that symbiosis may be necessary when P. vittata is growing under highly stressful conditions (wild P. vittata) or under low experimental soil fertility (cultured P. vittata) (Hajiboland et al. 2010). This symbiotic relationship between plant and fungi allowed completing its life cycle in different substrates, enabling the estimation of the colonization at all stages of the fern life cycle. To our knowledge, this is the first time the experimental AMF development in fern sporophytes and gametophytes under culture is reported.
Both gametophytes and sporophytes can form symbioses with the same Glomeromycota fungi that are common symbionts of phanerogamic plants. Previous studies showed that gametophytes of different genera as: Pteridium, Histiopteris, Todea, Cyathea, Asplenium, Blechnum and Schizaea pusilla grown in natural environments were colonized by arbuscular mycorrhiza when they were found in places where nutrients were unavailable (Cooper 1976, Swatzell et al. 1996). Although, as it was shown in this study, the development of the symbiosis between the gametophytes and the Glomalean fungi is not obligatory, we observed that the sporophyte was usually colonized by Glomus intraradices, leading to the notion that sporophytes could be colonized by other mycorrhizal fungus.
Our observation showed Paris-type colonization in cells of the gametophyte and sporophyte foot, while the mycelium never colonized the sporophyte through the junction between the gametophyte and the sporophyte, in line with Turnau (2005) and Reyes-Jaramillo et al. (2008) reports. Previous studies found hyphal coils in the gametophyte, near the penetration point considering this structure as an adaptation of the fungus to the limited growth of the gametophyte and the embryo foot (Schmid & Oberwinkler 1995). Gametophyte colonization is restricted to the basal zone of the midrib. Fungus colonization always occurs through the rhizoids after male gametangia differentiation. Previous experiments have shown that the gametophytes synthesize antheridiogen (Döpp 1950), which is a gibberellin-like compound (Wynne et al. 1998, Banks 1999). This change in the phytohormonal balance (Shaul-Keinan et al. 2002), enabling the hypha to penetrate the rhizoid and colonize the gametophyte, suggests a biochemical regulation of the mycorrhizal colonization.
Reyes-Jaramillo et al. (2008) found sporophyte root does not become colonized from the gametophyte; this finding is in agreement with our observations. In P. vittata the embryo foot established a close association with the gametophyte from which it takes nutrients for its growth, but was not colonized by the intracelular mycelium growing in the gametophyte. A plant tissue with several layers of uncolonized cells separated structures: embryo foot/gametophyte. Sporophyte colonization took place after development of root hairs. There were intercellular hyphae in the subepidermic tissue, parallel to the root surface, which formed intracellular branches that gave rise to arbuscules (Arum-type). This type of colonization has also been found in Gleichenia bifida (Schmid & Oberwinkler 1995). We report the simultaneous development of colonization units of Paris-type and Arum-type mycorrhizae with the same fungus in different stages of the life cycle concluding that the morphology of arbuscular mycorrhizae, (Arum versus Paris-types), is solely under the control of P. vittata genome during its life cycle.
Acknowledments
We wish to thank to UBA, CONICET and ANPCYT for financial support. The authors acknowledge especially to Mónica Ponce from Instituto Darwinion, Mónica Palacios-Rios from INECOL (Instituto de Ecología, A.C.) Xalapa, Veracruz, México, Romina Giacometti from CONICET and Roberto Fernandez IFYBIME (Instituto de Fisiología y Biología Molecular) for his technical assistance in confocal microscopy.
References
Banks, J.A. 1999. Gametophyte development in ferns. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 50: 163-186. [ Links ]
Boullard, B. 1957. La mycotropie chez les Pteridophytes. Sa frequence, ses caracterès, sa signification. Le Botaniste 41: 5-185. [ Links ]
Boullard, B. 1979. Considérations sur les symbiosis fongiques chez les Pteridophytes. Syllogeus 19: 1-59. [ Links ]
Brundrett, M.C. 2002. Coevolution of roots and mycorrhizas of land plants. New Phytol. 154: 275-304. [ Links ]
Cairney, J.W.G. 2000. Evolution of mycorrhiza systems. Naturwissenschaften 87: 467-475. [ Links ]
Cooper, K.M. 1976. A field survey of VA mycorrhizas in New Zealand ferns. N.Z.J. Bot. 14: 169-181. [ Links ]
Daniell, T.J., A. Hodge, J.P.W. Young & A. Fitter. 1999. How many fungi does it take to change a plant community? Trends Plant Sci. 4: 81-82. [ Links ]
Döpp, W. 1950. Eine die Anheridienbildung bei farnen fördernde Substanz in den Prothallien von Pteridium aquilinum L., Kuhn. Ber. Dtsch. Bot. Ges. 63: 139-47. [ Links ]
Fernandez, N., S. Fontenla & M. Messuti. 2010. Mycorrhizal status of obligate and facultative epiphytic ferns in a Valdivian Temperate Forest of Patagonia, Argentina. Am. Fern J. 100: 16-26. [ Links ]
Hajiboland, R., N. Aliasgharzadeh, S.F. Laiegh & C. Poschenrieder. 2010. Colonization with arbuscular mycorrhizal fungi in proves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 331: 313-327. [ Links ]
Janos, D.P. 1993. Vesicular-arbuscular mycorrhizae of epiphytes. Mycorrhiza 4: 1-4. [ Links ]
Janos, D.P. 2007. Plant responsiveness to mycorrhizae differs from dependence upon mycorrhizae. Mycorrhiza 17: 75-91. [ Links ]
Leung, H.M., Z.H. Ye & M.H. Wong. 2006. Interactions of mycorrhizal fungi with Pteris vittata (As hyperaccumulator) in As-contaminated soils. Environ. Pollut. 139: 1-8. [ Links ]
Martinez, O.G. 2010. Gametófitos y esporófitos jóvenes de cuatro especies de helechos del género Pteris (Pteridaceae) naturalizadas en América. Rev. Biol. Trop. 58: 89-102. [ Links ]
Nayar, B.K. & S. Kaur. 1969. Types of prothallial development in homosporous ferns. Phytomorphology 19: 179-188. [ Links ]
Peterson, R.L., H.B. Massicotte & L.H. Melville. 2004. Mycorrhizas: Anatomy and Cell Biology. NRC Research, Ottawa, Canada. [ Links ]
Phillips, J.M. & D.S. Hayman. 1970. Improved procedures for clearing roots and staining parasitic and VA mycorrhizal fungi for rapid assesment of infection. Trans. Br. Mycol. Soc. 5: 158-161. [ Links ]
Pirozynski, K.A. & D.W. Malloch. 1975. The origin of land plants: a matter of mycotrophism. Biosystems 6: 153-164. [ Links ]
Read, D.J. 1999. The ecophysiology of mycorrhizal symbiosis with special reference to impacts upon plant fitness, p. 133-152. In J.D. Scholes & M.G. Barker (eds.). Physiological Plant Ecology. MC, Blackwell Science, London. [ Links ]
Reyes-Jaramillo, I., S. Camargo-Ricalde & M. Aquiahuatl-Ramos. 2008. Mycorrhizal-like interaction between gametophytes and young sporophytes of the fern Dryopteris muenchii (Filicales) and its fungal endophyte. Rev. Biol. Trop. 56: 1101-1107. [ Links ]
Schmid, E. & F. Oberwinkler. 1995. A light-microscopic and electron-microscopic study on a vesicular arbuscular host-fungus interaction in gametophytes and young sporophytes of the Gleicheniaceae (Filicales). New Phytol. 129: 319-324. [ Links ]
Shaul-Keinan, O., V. Gadkar, I. Ginzberg, J. Grúnzweig, I. Chet, Y. Elad, S. Wininger, E. Belausov, Y. Eshed, N. Atzmon, Y. Ben-Tal & Y. Kapulnik. 2002. Hormone concentrations in tobacco roots change during arbuscular mycorrhizal colonization with Glomus intraradices. New Phytol. 154: 501-507. [ Links ]
Smith, S.E. ; D.J. Read. 1997. Mycorrhizal symbiosis. Academic, London, United Kingdom. [ Links ]
Smith, F.A. ; S.E. Smith. 1997. Structural diversity in (versicular arbuscular mycorrhizal symbioses. New Phytol. 137: 373-388. [ Links ]
Swatzell, L.J., M.J. Powell & J.Z. Kiss. 1996. The relationship of endophytic fungi to the gametophyte of the fern Schizaea pusilla. Int. J. Plant Sci. 157:53-62. [ Links ]
Turnau, K., T. Anielska & A. Jurkiewiez. 2005. Mycothallic/mycorrhizal symbiosis of chlorophyllous gametophytes and sporophytes of a fern, Pellaea viridis (Forsk.) Prantl (Pellaeaceae, Pteridales). Mycorrhiza 15: 121-128. [ Links ]
Trotta, A., P. Falaschi, L. Cornara, V. Minganti, A. Fusconi, G. Drava & G. Berta. 2006. Arbuscular mycorrhizae increase the arsenic translocation factor in the As hyperaccumulating fern Pteris vittata L. Chemosphere 65: 74-81. [ Links ]
van Aarle, I.M., T.R. Cavagnaro, S.E. Smith, F.A. Smith & S. Dickson. 2005. Metabolic activity of Glomus intraradices in Arum- and Paris-type arbuscular mycorrhizal colonization. New Phytol. 166: 611-618. [ Links ]
Wynne, G.M., L.N. Mander, N. Goto, H. Yamane & T. Omori. 1998. Gibberellin A117 methyl ester, a new antheridiogen from Lygodium circinatum. Phytochem. 49: 1837-1840. [ Links ]
Zhi-wei, Z. 2000. The arbuscular mycorrhizas of pteridophytes in Yunnan southwest China: evolutionary interpretations. Mycorrhiza 10: 145-149. [ Links ]
Zhang, K.M., I. Shi, X.C. Zhang, C.D. Jiang & W.L. Tim-Chum. 2008. Gametophyte morphology and development of six Chinese species of Pteris (Pteridaceae). Am. Fern J. 98: 33-41. [ Links ]
*Correspondencia a: Alicia E. Martinez, Lo Tai Em, María A. Rodriguez & Alicia M. Godeas: Departamento de Biodiversidad y Biología Experimental. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. Av. Int. Güiraldes s/N. Pabellón II. Ciudad Universitaria. 1428, Buenos Aires, Argentina; martae@bg.fcen.uba.ar, tai@bg.fcen.uba.ar, arodrig@bg.fcen.uba.ar, godeas@bg.fcen.uba.ar
Viviana Chiocchio: Cátedra de Microbiología Agrícola y Ambiental. Facultad de Agronomía. Universidad de Buenos Aires - Av. San Martín 4453. 1417. Buenos Aires. Argentina; chiocchi@agro.uba.ar
Alicia E. Martinez, Viviana Chiocchio, María A. Rodriguez & Alicia M. Godeas: INBA – CONICET
1. Departamento de Biodiversidad y Biología Experimental. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. Av. Int. Güiraldes s/N. Pabellón II. Ciudad Universitaria. 1428, Buenos Aires, Argentina; martae@bg.fcen.uba.ar, tai@bg.fcen.uba.ar, arodrig@bg.fcen.uba.ar, godeas@bg.fcen.uba.ar
2. Cátedra de Microbiología Agrícola y Ambiental. Facultad de Agronomía. Universidad de Buenos Aires - Av. San Martín 4453. 1417. Buenos Aires. Argentina; chiocchi@agro.uba.ar
3. INBA – CONICET
Received 09-V-2011. Corrected 10-XI-2011. Accepted 12-XII-2011.


{kind=link}
{kind=link}
{kind=link}