










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Infecciones parasitarias del coyote, Canis latrans (Carnivora: Canidae) en un Parque Nacional y una zona agrícola en Costa Rica
*Dirección para correspondencia:
Abstract
As human populations expand into wild habitats with their pets and livestock, the potential spread of disease to wildlife or vice versa increases. Because, wild and domestic canids may pose as reservoirs or disseminators of infectious diseases (including parasites), coyotes (Canis latrans) may also serve as indicators of ecological health. In Costa Rica, little information exists on coyote parasites, making research necessary to identify potential zoonotic interactions. For this reason, a survey of the coyote parasites was performed in a mixed area of protected woodland and agricultural land, surrounding Irazu Volcano National Park (IVNP) in Cartago, Costa Rica. Over a one-year period, 209 fecal samples were collected directly from the ground. Collection took place on a monthly basis in a trail sectioned into three sub-areas named Irazú (closest to the volcano), potato fields (where potatoes were cultivated), and Prusia (a protected sector of IVNP). Sectioning the trail allowed separate collection and analysis of the samples, where 99 were obtained from Irazú, 11 from potato fields and 99 from Prusia. Using direct examination and mechanical concentration 36.84% positive samples containing at least one helminth were found. The presence of parasites was similar for both woodland areas (33.3% in Prusia and 37.4% in Irazú), but differed from the 63.6% observed in the potato fields. Hookworm parasites (probably Ancylostoma caninum), threadworms (possibly Strongyloides sp.), Toxocara canis, Trichuris sp. and Taenia pisiformis were identified, as well as Hymenolepis diminuta, possible spurious parasite resulting from the ingestion of rodents by coyotes. Seasonal details are discussed, concluding that wet and dry seasons affect presence of parasites. Some remarks are made on the importance of these first findings for Costa Rica, especially considering the systematic way in which the collection of samples was carried out. Rev. Biol. Trop. 60 (2): 799-808. Epub 2012 June 01.
Key words: Canis latrans, coyotes, intestinal parasites, Costa Rica, helminths.
Resumen
Conforme las poblaciones humanas se expanden hacia los hábitats silvestres con sus mascotas y ganado, el potencial de transmisión de enfermedades hacia los animales silvestres -y viceversa- aumenta, y hace necesario identificar interacciones zoonóticas potenciales. Los cánidos domésticos y silvestres pueden funcionar como reservorios o diseminadores de enfermedades infecciosas (se incluyen parásitos), por lo que el coyote (Canis latrans) puede también servir como indicador de la salud ecológica. Asimismo, se estudiaron los parásitos de 209 muestras de heces de coyotes en una zona mixta de área silvestre protegida y campo agrícola del Parque Nacional Volcán Irazú (PNVI) en Costa Rica. La recolección fue realizada mensualmente durante un año en tres sub-áreas denominadas: Irazú (la más cercana al volcán), papales (por el cultivo de papas), y Prusia (un sector del PNVI). Entonces, se empleó examen directo y concentración mecánica, se obtuvo 36.84% de muestras positivas por al menos un helminto. La presencia de parásitos fue muy similar para ambos sectores boscosos del PNVI (33.3% en Prusia y 37.4% en Irazú), pero contrastó con el 63.63% observado en los papales. También, se identificaron uncinarias (probablemente Ancylostoma caninum), estrongilidios (posiblemente Strongyloides sp.), Toxocara canis, Trichuris sp. y Taenia pisiformis, así como Hymenolepis diminuta, probablemente un parásito espurio proveniente de roedores ingeridos por los coyotes. Se comenta la importancia de estos primeros hallazgos y se concluye que las estaciones seca y lluviosa influyen en la presencia de los parásitos.
Palabras clave: Canis latrans, coyotes, parásitos intestinales, Costa Rica, helmintos.
El coyote (Canis latrans) es una especie generalista y oportunista (de la Rosa & Nocke 2000) distribuida en una amplia gama de ecosistemas (Vaughan 1983) y adaptable a los recursos alimenticios locales (Vaughan & Rodriguez 1986). En las áreas silvestres protegidas, como el Parque Nacional Volcán Irazú, existe un impacto ambiental importante debido a la pérdida y fragmentación del hábitat, la cacería y la urbanización progresiva. Las características adaptativas del coyote le han permitido instalarse en zonas agrícolas y suburbanas, en donde expande su territorio (Riley et al. 2003) y favorece el contacto con especies domésticas (Trout et al. 2006). El acercamiento entre animales silvestres y domésticos aumenta la probabilidad (Manning 2007) y altera el mecanismo de transmisión de enfermedades (Aguirre 2009). Algunos países cuentan con estudios de enfermedades infecciosas en cánidos de vida libre (Acha & Szyfres 2003), caso contrario del Neotrópico y específicamente de Costa Rica, donde los reportes son escasos, incluso los de endoparásitos. Esta falta de datos es preocupante, si se toma en cuenta el riesgo que pueden representar los parásitos para las poblaciones de esta y otras especies de animales silvestres, domésticos e incluso el ser humano.
Los cánidos silvestres cumplen un rol de centinelas bioacumuladores sin ser reservorios (Barnes 1982, Ver Cauteren et al. 2008). Al existir una relación trófica entre centinelas y las especies de interés, los animales carnívoros u omnívoros pueden funcionar como indicadores de enfermedades, incluso zoonóticas, en las poblaciones locales de los animales afectados (Arjo et al. 2003, Sangster et al. 2007). El muestreo directo en animales de vida libre resulta complicado. Al mismo tiempo, un muestreo realizado en animales en cautiverio no necesariamente refleja la situación real en animales silvestres.
Por otra parte, las necropsias ofrecen información acerca de las parasitosis de los hospederos (Hirsch & Gier 1974, Van Den Bussche et al. 1987), pero la captura y eutanasia de los animales no siempre es indicada. Ante las limitaciones apuntadas, la coprologia parasitaria resulta un método confiable para determinar la helmintofauna que alberga una población de cánidos silvestres (Torres et al. 2001).
Por las razones anteriores y debido a la falta de estudios poblacionales y a la imposibilidad de capturar coyotes en el Parque Nacional Volcán Irazú, se decidio estudiar los parásitos presentes en las heces de C. latrans recolectadas en un sendero que comunica el sector Irazú con Prusia.
Materiales y métodos
Área de estudio y recolecta de las heces de coyotes: El estudio se realizó en el Parque Nacional Volcán Irazú (PNVI) en Costa Rica (9°95'12'' - 9°99'63'' N - 83°87'55'' - 83°83'87'' W), y se siguió un sendero que abarcó los sectores llamados Irazú y Prusia, así como unas fincas cultivadas con papa (Solanum tuberosum). Este trayecto posee distintas alturas, microclimas, dos zonas de vida: bosque pluvial montano y bosque muy húmedo montano, así como un área de páramo (ACCVC 2008).
La recolecta de muestras se inicio en el sector Irazú (9°97'05'' N - 83°85'93'' W) con una distancia aproximada de 1.2km. Debido a las características físicas del área de estudio, el sendero se sale de los límites del parque en un punto (9°96'07'' N - 83°86'10'' W), puesto que atraviesa 1.3km de fincas cultivadas con papa, denominado sector papales. Seguidamente, el transecto se reintroduce en el área del Parque (9°95'67'' N - 83°86'50'' W), punto a partir del que continuúa el sector de Prusia, con un recorrido de alrededor de 2.65km. Este sector presenta un bosque con áreas de vegetación tanto autóctona como exótica, por la introducción de pino (Pinus sp.), ciprés (Cupressus sp.) y eucalipto (Eucalyptus sp.). Existen también senderos transitados por turistas y una carretera interna en mal estado. El recorrido se completa con 5.15km en el area administrativa del sector Prusia (9°95'35'' N - 83°88'13'' W).
La obtencion, de forma mensual, de heces de coyotes se llevo a cabo entre agosto 2008- julio 2009 en compania de un funcionario del PNVI. En la zona, los reportes de avistamientos y aullidos de coyotes por finqueros son comunes y se cuenta con evidencia fotográfica de camaras trampa colocadas en el Parque. Las muestras fueron seleccionadas, con base en la experiencia de los guardaparques, ademas de seguir las caracteristicas comparativas de rastros descritas por Murie (1974) y Aranda (2000).
Aunque, en las fincas aledanas al PNVI existen perros, es raro que ingresen al Parque. Sin embargo, para evitar al maximo la recolección de excretas de otros animales, por ejemplo perros, unicamente se tomo muestras de heces consistentes, mayores a 1cm de diametro y 10cm de largo, con abundantes restos de pelo y/o fragmentos oseos, ademas del olor y la forma caracteristicos de las excretas de coyote debido a su dieta. Asimismo, se omitio la toma de muestras semiliquidas que no tuvieran un volumen considerable, al igual que cualquier muestra que se sospechara que perteneciera a un perro domestico (C. familiaris). Aún, al seguir estas precauciones, cabe la posibilidad de que se recolectara alguna muestra que no fuera de coyote, puesto que no se realizo caracterización molecular de las heces.
Análisis de muestras y pruebas de laboratorio: Las muestras fueron clasificadas según el sitio de muestreo y el tiempo de deyección. Debido a la gran cantidad de excretas viejas encontradas, estas fueron incorporadas al estudio, las cuales se subdividieron en frescas (menos de cuatro días) y viejas (cuatro días o mas). Esta clasificación se baso en características organolepticas como apariencia física, olor, consistencia y la relación de materia fecal con los restos permanentes (pelo y huesos). La prevalencia se calculó como la frecuencia del número de muestras positivas sobre el número de muestras examinadas.
En el Laboratorio de parásitologia de la Universidad de Ciencias Medicas (UCIMED) se llevo a cabo el análisis coproparásitoscopico, el cual consistió en la observacion directa en solución salina al 0.85% y en Lugol de Anthony. El mismo procedimiento se siguió para las muestras concentradas por el método de sedimentación con solución salina al 0.85%.
También, se utilizó estadística no paramétrica para la prueba t de student y la prueba de Mann Whitney en los correspondientes análisis de variancia, y se supone, tanto igualdad como desigualdad de variancia para la prueba de t.
Resultados
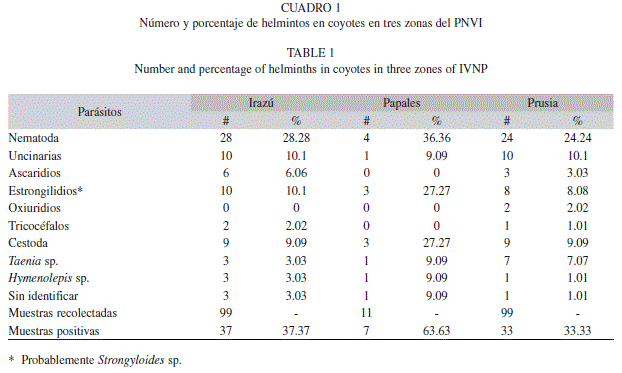
De las 209 muestras obtenidas durante un año, 77 resultaron positivas por algún helminto, con una prevalencia de 36.84% (Cuadro 1). Del total de muestras, 99 fueron recolectadas en el sector Irazú, 99 en Prusia y 11 en los papales. Este sector, presentó la mayor proporción de heces infectadas (63.63%). Seguidamente, el sector Irazú obtuvo 37.37% de muestras positivas para helmintos y Prusia 33.33%, y no existen diferencias estadísticas significativas en el análisis estadístico no paramétrico (p=0.59 para helmintos, 0.83 nemátodos y 0.48 céstodos).
El grupo más representativo de helmintos encontrados fue el de nemátodos (26.8%) y luego los céstodos (10.05%). A lo largo del estudio, las uncinarias y los estrongilidios (10.05% para cada uno) representaron los grupos más importantes de nemátodos. Adicionalmente, otros géneros fueron encontrados en menos del 5%: Toxocara canis (4.31%), Trichuris sp. (1.44%) y oxiuridios (0.96%). Para los céstodos, Taenia pisiformis constituyó 5.26% mientras que Hymenolepis sp. y los céstodos sin identificar conformaron 2.39% (Cuadro 1). Además, en dos ocasiones (0.96%) se observaron huevecillos de acaros cuyos géneros no fueron identificados.
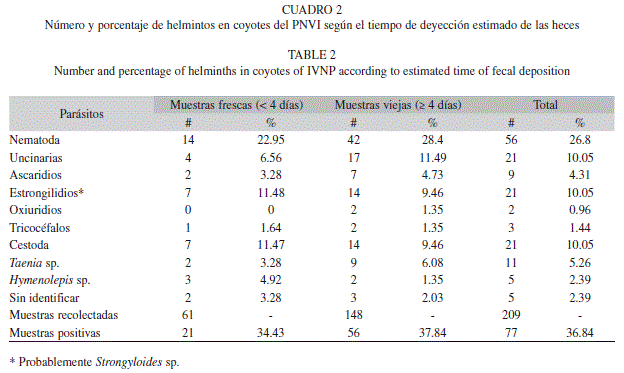
Al separar las muestras según el tiempo de deyección estimado, se observó que los porcentajes de positividad eran similares, aunque, en las muestras viejas resultó ligeramente mayor: 34.43% (21 de 61) para las muestras frescas y 37.84% (56 de 148) para las viejas (Cuadro 2). En general, la presencia de helmintos fue estadísticamente significativa en las muestras viejas (27.7%) en comparación con las frescas (22.95%). De tal manera que con un nivel de significancia del 10% para el promedio de muestras con helmintos (p=0.07 para la prueba no parametrica), se encuentran diferencias entre los dos grupos, pero no para los nemátodos (p=0.14) ni para los céstodos (p=0.26).
Los estrongilidios y Trichuris sp. Resultaron ser mas abundantes en las muestras frescas (11.48 % y 1.64% respectivamente) que en las viejas (9.46% y 1.35%), pero los anquilostomidios y ascaridios (T. canis) fueron mas altos en las muestras viejas (11.49% y 4.73% versus 6.56% y 3.28%, respectivamente). Los oxiuridios se encontraron solamente en las muestras viejas (1.35%). Los céstodos tuvieron una prevalencia similar en ambos grupos (11.47% en frescas y 9.46% en viejas), T. pisiformis fue la más común en las muestras viejas (6.08% vs. 3.28%) e Hymenolepis sp. en las frescas (4.92% vs. 1.35%). Algunos huevecillos de céstodos no identificados fueron observados en ambos grupos y en porcentajes semejantes (3.28% en frescas y 2.03% en viejas).
La mayor cantidad de muestras positivas se presentaron en el mes de agosto (60%) y la
menor cantidad en febrero, abril y diciembre (19.2%, 13% y 15.4%, respectivamente). En general, se observó un patrón ondulatorio, con una mayor prevalencia de helmintos durante la época lluviosa, es decir, de agosto a noviembre 2008, y de mayo a junio 2009 (Fig. 1).
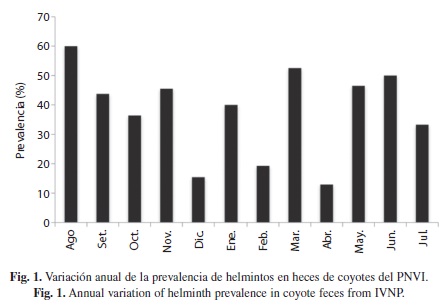
Los nemátodos transmitidos por medio de larvas infectivas, uncinarias y estrongilidios, presentaron un incremento considerable durante la época lluviosa (Fig. 2). Los helmintos más constantes durante todo el año fueron los estrongilidios (11 meses) y las uncinarias (nueve meses), mientras que los demás fueron mas fluctuantes. Los céstodos no tuvieron diferencias estacionales significativas; sin embargo, a diferencia de los nemátodos, se mantuvieron presentes o elevados (como el caso de T. pisiformis) durante los meses mas secos del año (Fig. 3).
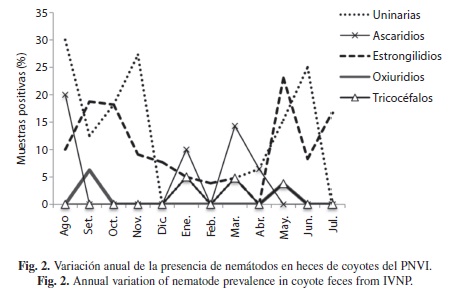
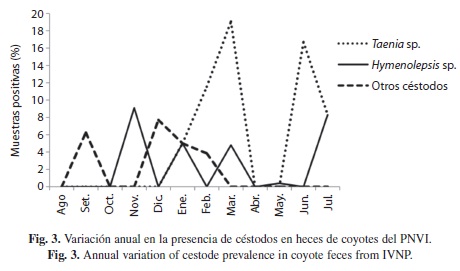
Parásitos encontrados y taxones por muestra: Se encontraron ocho grupos de helmintos (táxones) dentro del área de estudio, de los cuales solamente uno (A. caninum) ha sido reportado previamente en coyotes de Costa Rica (Rodríguez-Ortíz et al. 2004). Del total de helmintos, cuatro fueron comunes en los tres sitios de recolecta, otros seis fueron comunes para las zonas de Irazú y Prusia, mientras que solamente un taxon (oxiuridios) fue exclusivo de Prusia.
De las 209 muestras analizadas, 23% contenían un taxón de helminto, 6.7% contenían dos y el 0.48% cuatro táxones. La mayor proporción de muestras con un único helminto la obtuvo el grupo de excretas frescas con 29.5%, versus 21.62% en viejas. La mayor cantidad de muestras con dos táxones fue observada en las heces viejas (7.43%), mientras que las frescas obtuvieron sólo 1.64%. Además, en las heces viejas se observó la única muestra con cuatro táxones de helmintos (0.67%).
Las muestras de Irazú obtuvieron el mayor porcentaje con un helminto o taxon (29.29%), seguido por el sector papales (27.27%) y Prusia (16.16%). Las muestras con dos táxones de parásitos fueron prevalentes en los papales con 18.2%, seguidos por la región de Prusia (8.1%) e Irazú con 4.04%. Las muestras con cuatro táxones fueron exclusivas de Prusia (1%).
Discusión
Debido a que no se encontró ningún estudio similar de parásitos en Centroamérica, no se pudieron hacer comparaciones directas sino solamente, referencia a los estudios disponibles en cánidos silvestres para Norteamérica y Europa, donde las condiciones ambientales son totalmente diferentes. La prevalencia observada (36.84%) difiere mucho de las obtenidas por Gompper et al. (2003) en coyotes de Nueva York (56%), así como del 72% encontrado en excretas de lobos ibéricos (Domínguez & de la Torre 2002) y del 14% en heces de lobos del Ártico (Marquard-Petersen 1997). Aunque, es común que un porcentaje de la población se encuentre sin parasitar, la incorporación de muestras viejas al análisis podría favorecer esta situación. Sin embargo, no hubieron diferencias importantes entre los resultados obtenidos para muestras frescas y viejas.
Los parásitos encontrados son comunes en los cánidos domésticos y silvestres, con excepción de los oxiuridios observados en dos de las muestras viejas del sector Prusia. Por lo tanto, se puede deducir que su presencia se debió a la contaminación proveniente de las presas de estos animales. En efecto, se han encontrado estos parásitos junto con restos de mamíferos pequeños (conejos y roedores) en estómagos e intestinos gruesos de coyotes (Butler & Grundmann 1954, Thornton et al. 1974) en Estados Unidos.
Entonces se encontraron parásitos cuyo sistema de trasmisión corresponde a ciclo directo (uncinarias, estrongilidios, oxiuridios y Trichuris sp.), indirecto (Taenia e Hymenolepis) o ambos ciclos (T. canis), en una relación de 4:2:1 (directo:indirecto:ambivalente). En relación con este aspecto, Domínguez & de la Torre (2002) establecen hallazgos importantes de parásitos de transmisión directa que pueden relacionarse con factores ambientales, densidades de población de los hospederos y otros cánidos silvestres como las zorras rojas. En ambos sectores del PNVI, los coyotes y las zorras grises (Urocyon cinereoargenteus) comparten el territorio, lo que posibilita la transmisión de estos parásitos entre ellos. Sin embargo, no se incluyeron excretas de zorra gris en el análisis por lo que esta asociación no pudo establecerse.
La diferencia estacional en las prevalencias de uncinarias y estrongilidios a lo largo del año puede relacionarse con el tipo de parásitos, en vista de que se trata de helmintos que utilizan el estadio evolutivo de larva como método de transmisión. Por tanto, aunque los coyotes se encontraran infectados durante todo el año, la forma larvaria de estos parásitos es poco resistente a las condiciones ambientales, y es probable que en los meses secos de febrero y marzo se reduzca la transmisión. Lo contrario sucede en el caso de los helmintos transmitidos por medio de huevecillos, como Toxocara canis, Trichuris sp. y Taenia pisiformis, que continuaron presentes durante estos meses. Esto puede explicarse por la mayor resistencia de estos huevecillos (Hays 1977, Theis et al. 1978, Zunino et al. 2000).
La prevalencia y abundancia parasitaria similares para Irazú y Prusia (seis táxones en común) se puede deber al clima y las características boscosas de la zona, donde las temperaturas permanecen bajas y la precipitación se mantiene a lo largo del año. Esto permite preservar las heces por mayor tiempo, y a los parásitos que contienen. En contraste, lo observado en el área de papales (cuatro táxones de parásitos) es atribuible a la exposición ambiental directa, lo que favorece su rápida degradación (Acha & Szyfres 2003).
La prevalencia de uncinarias obtenida en este estudio es similar (10.05%) a la encontrada en lobos ibéricos (11.1% para U. stenoceophala y 16.6% para A. caninum) por Domínguez & de la Torre (2002). Sin embargo, Gompper et al. (2003) encontraron prevalencias variables de U. stenocephala en heces de coyotes de distintas localidades (1.5% versus 26.1%) en Estados Unidos, probablemente causadas por diferencias en las condiciones ambientales.
La infeccion elevada con uncinarias junto a factores nutricionales y etarios, son una causa importante de mortalidad en zorras y coyotes (Radomski 1989). Ejemplo de lo anterior es A. caninum, el cual representa un regulador natural de las poblaciónes de coyotes de vida libre, y funciona como un importante factor de mortalidad neonatal (Pence et al. 1988, Henke et al. 2002). Estos detalles dan una idea general de la situación poblacional de los coyotes adultos, dado que altas prevalencias suelen correlacionarse con altas densidades de población. Generalmente, al conocer que los animales jóvenes presentan mayores índices de infección (Franson et al. 1978), los resultados podrían ser diferentes si las heces pequeñas se hubieran incluido en el muestreo.
Este factor pudo haber influido en el caso de los ascaridios, ya que el porcentaje encontrado (4.3%) es menor al observado por Gompper et al. (2003), en el que hubo 8.7% positivos para Toxocara canis y Toxascaris leonina. A pesar de esto, se asemeja a la prevalencia obtenida en heces de lobos ibéricos (5.5% para ambas especies) por Domínguez & de la Torre (2002).
La baja prevalencia observada en Trichuris sp. (1.4%) contrasta con otros trabajos en cánidos silvestres, donde se encuentran arriba del 10% (Domínguez & de la Torre 2002, Alarcón-Navarro 2005). Sin embargo, existen datos menores al 1% en algunos estudios (Pence & Meinzer 1979, Marquard-Petersen 1997). Dada la similitud en la trasmisión de ascaridios y Trichuris sp. (Acha & Szyfres 2003) y en la relación estrecha entre los hallazgos coprológicos y por necropsia que se ha encontrado en lobos ibéricos para ambos parásitos (Torres et al. 2001), se esperarían datos similares para los dos organismos en ambos sectores boscosos.
Tanto los cánidos domésticos como silvestres son hospederos del parásito y constituyen sus reservorios (Bowman 2008), por lo que es posible que exista una importante fuente de infección entre coyotes y perros en el área de estudio.
Solamente, se encontró compatibilidad con el hallazgo de estrongilidios en otro estudio realizado en zorro gris suramericano (Alarcón-Navarro 2005). Algunas de las larvas observadas pueden corresponder a Strongyloides, puesto que se conoce la presencia de S. stercoralis en cánidos (Forrester 1992). Dada su importancia médica para el hombre, especialmente para los individuos alcohólicos e inmunosuprimidos (Llagunes et al. 2010), el hallazgo resulta importante desde el punto de vista de la salud pública. La alta prevalencia para estrongilidios puede deberse a las características adaptativas de las larvas a las condiciones ambientales locales (Acha & Szyfres 2003).
Los céstodos se observaron en las tres zonas (Cuadro 2). Taenia pisiformis se concentró mayormente en la zona baja (papales 9% y Prusia 7%) con una prevalencia general de 5.3%. Entonces, se documenta un 11% de prevalencia para T. pisiformis en coyotes (Gompper et al. 2003) y un 5% para Taeniidae en heces de lobos del Ártico (Marquard-Petersen 1997). La forma quística de T. pisiformis se ubica en conejos, probables fuentes de infección para los coyotes.
Por otro lado, Hymenolepis sp., encontrada principalmente en la zona alta (papales 9% e Irazú 3%) con una prevalencia general de 2.4%, era morfológicamente similar a H. diminuta, un parásito que no ha sido reportado parasitando coyotes. Los reportes de Hymenolepis sp. en heces de coyotes de Florida (Manning 2007), H. diminuta en heces de zorros rojos en Eslovaquia (Miterpáková et al. 2009), al igual que H. fasciata en necropsias de zorros grises suramericanos (Alarcón-Navarro 2005), no son conclusivos para determinar que esta especie realmente se encuentra en C. latrans.
La fase larvaria (cisticercoide) de H. diminuta se desarrolla en pulgas, coleópteros, lepidópteros, miriápodos y blatodeos, los cuales pueden ser ingeridos por los coyotes o bien, puede tratarse de parasitismo espurio, causado por la ingestión de roedores hospederos en los que Hymenolepis spp. es frecuente (Miterpáková et al. 2009).
A partir de este primer trabajo sobre las infecciones parasitarias en coyotes, se concluye lo siguiente:
• El estudio permite dar una idea de la fauna parasitaria entérica de los coyotes de vida libre en un área mixta que abarca un Parque Nacional y campos de cultivo en una zona volcánica del país. Aunque, no existen estudios similares previos en la zona que permitan hacer comparaciones, se puede establecer como enzoóticos los agentes encontrados en la región de Irazú.
• La presencia de perros en fincas aledañas al PNVI favorece el contacto directo con las heces de coyotes y viceversa. Esto puede causar una transmisión bidireccional entre perros y coyotes, lo que reflejaría una fauna parasitaria muy similar para ambos cánidos, de tal manera que hace necesario evaluar el estado sanitario de los perros domésticos de la zona.
• Asimismo, debe tomarse en cuenta que al ser la zona turística, la presencia de heces en los senderos del parque figura como una fuente potencial de infección para los visitantes con helmintos zoonóticos como A. caninum, H. diminuta, S. stercoralis, T. canis y T. vulpis.
El muestreo periódico en estos y otros carnivoros se hace necesario, pues ante la evidencia limitada o nula sobre la función ecológica que cumplen los parásitos en los diferentes hábitats, estos controles pueden descubrir variaciones en la prevalencia de estos agentes infecciosos, así como factores asociados a estas variaciones. De esta forma, podrán comprenderse mejor los riesgos reales que representan los parásitos y definir medidas sostenibles para la prevención y el manejo de los problemas que trae consigo la perturbación antropogénica creciente en áreas silvestres (Arjo et al. 2003).
Agradecimientos
Se le agradece al Sistema Nacional de Áreas de Conservación (SINAC), en especial a Mauricio Gamboa Ramírez y a Daniel Núñez Montenegro del Parque Nacional Volcán Irazú (PNVI) por su valiosa colaboración en el estudio, así como a la Cátedra de parásitología y al Departamento de Investigación Básica de la Universidad de Ciencias Médicas (UCIMED). También se agradece a Juan Carlos Vanegas por los análisis estadísticos realizados.
Referencias
ACCVC & Onca Natural. 2008. Plan Maestro-Parque Nacional Volcán Irazú. SINAC-ACCVC, San José, Costa Rica. [ Links ]
Acha, P.N. & B. Szyfres. 2003. Zoonosis y enfermedades transmisibles comunes al hombre y a los animales, p. V-XVIII. Volumen III. parásitosis. Organización Panamericana de la Salud. Washington D.C., EEUU. [ Links ]
Aguirre, A.A. 2009. Wild canids as sentinels of ecológical health: a Conservation Medicine perspective. Parasit. Vectors 2: S7. [ Links ]
Alarcón-Navarro, U.F. 2005. Estudio taxonómico de la fauna parasitaria del tracto gastrointestinal de zorro gris (Pseudalopex griseus, Gray 1837), en la XII región de Magallanes y Antártica chilena. Tesis de Licenciatura, Universidad Austral de Chile, Valdivia, Chile. [ Links ]
Aranda, M. 2000. Huellas y rastros de los mamíferos grandes y medianos de México. Instituto de Ecología, Xalapa, Veracruz, México. [ Links ]
Arjo, W.M., B. Gese, C. Bromley, A. Kozlowski & E.S. Williams. 2003. Serologic survey for diseases in freeranging coyotes (Canis latrans) from two ecológically distinct áreas of Utah. J. Wildl. Dis. 39: 449-455. [ Links ]
Barnes, A.M. 1982. Surveillance and control of bubonic plague in the United States. Symp. Zool. Soc. Lond. 50: 237-270. [ Links ]
Bowman, D.D. 2008. Parasite 101 - A clinical overview of whipworms. Vet. Forum Agosto: 48-70. [ Links ]
Butler, J.M. & A.W. Grundmann. 1954. The intestinal helminths of the coyote Canis latrans Say in Utah. J. parásitol. 40: 1-4. [ Links ]
De la Rosa, C.L. & C.C. Nocke. 2000. A guide to the carnivores of Central America. University of Texas, Austin, Texas, USA. [ Links ]
Domínguez, G. & J.A. de la Torre. 2002. Aportaciones al conocimiento de los endoparásitos del lobo ibérico (Canis lupus signatus, Cabrera 1907) en el Norte de Burgos. Galemys 14: 49-58. [ Links ]
Franson, J.C., R.D. Jorgenson, E.K. Boggess & J.H. Greve. 1978. Gastrointestinal parasitism of Iowa coyotes in relation to age. J. parásitol. 64: 303-305. [ Links ]
Forrester, D.J. 1992. Parasites and diseases of wild mammals in Florida. University Press of Florida, Gainesville, Florida, USA. [ Links ]
Gompper, M.E., R.M. Goodman, R.W. Kays, J.C. Ray, C.V. Fiorello & S.E. Wade. 2003. A survey of the parasites of coyotes (Canis latrans) in New York based on fecal analysis. J. Wildl. Dis. 39: 712-717. [ Links ]
Hays, B.D. 1977. Potential for parasitic disease transmission with land application of sewage plant effluents and sludges. Water Res. 11: 583-595. [ Links ]
Henke, S.E., D.B. Pence & F.C. Bryant. 2002. Effect of short-term coyote removal on populations of coyote helminths. J. Wildl. Dis. 38: 54-67. [ Links ]
Hirsch, R.P. & H.T. Gier. 1974. Multiple-species infections of intestinal helminths in Kansas coyotes. J. parásitol. 60: 650-653. [ Links ]
Llagunes, J., E. Mateo, J.J. Peña, P. Carmona & J. de Andrés. 2010. Hiperinfección por Strongyloides stercoralis. Med. Intensiva 34: 353-356. [ Links ]
Manning, D.L. 2007. A comparative ecológical study between coyotes (Canis latrans) in a protected and urban hábitat: a closer look at enteric parasites and diet between Florida coyotes. Tesis de Maestria, University of South Florida, Florida, EEUU. [ Links ]
Marquard-Petersen, U. 1997. Endoparasites of arctic wolves in Greenland. Arctic 50: 349-354. [ Links ]
Miterpáková, M., Z. Hurníková, D. Antolová & P. Dubinský. 2009. Endoparasites of red fox (Vulpes vulpes) in the Slovak Republic with the emphasis on zoonotic species Echinococcus multilocularis and Trichinella spp. Helmintologia 46: 73-79. [ Links ]
Murie, O.J. 1974. Animal tracks. Houghton Mifflin, Nueva York, Nueva York, EEUU. [ Links ]
Pence, D.B. & W.P. Meinzer. 1979. Helminth parasitism in the coyote, Canis latrans, from the Rolling Plains of Texas. Int. J. parásitol. 9: 339-344. [ Links ]
Pence, D.B., F.F. Knowlton & L.A. Windberg. 1988. Transmission of Ancylostoma caninum and Alaria marcianae in coyotes (Canis latrans). J. Wildl. Dis. 24: 560-563. [ Links ]
Radomski, A.A. 1989. Host-parasite relationships of helminths in a coyote population from Southern Texas with particular reference to the dog hookworm. Tesis de Maestria, Texas Tech University, Lubbock, Texas, EEUU. [ Links ]
Riley, S.P.D., R.M. Sauvajot, T.K. Fuller, E.C. York, D.A. Kamradt, C. Bromley & R.K. Wayne. 2003. Effects of urbanization and hábitat fragmentation on bobcats
and coyotes in Southern California. Conservat. Biol. 17: 566-576. [ Links ]
Rodríguez-Ortíz, B., L. García-Prieto & G. Pérez-Ponce de León. 2004. Checklist of the helminth parasites of vertebrates in Costa Rica. Rev. Biol. Trop. 52: 313-354. [ Links ]
Sangster, C., D. Bergeson, C. Lutze-Wallace, V. Crichton & G. Wobeser. 2007. Feasibility of using coyotes (Canis latrans) as sentinels for bovine mycobacteriosis (Mycobacterium bovis) infection in wild cervids in and around Riding Mountain National Park, Manitoba, Canada. J. Wildl. Dis. 43: 432-438. [ Links ]
Theis, J.H., V. Volton & D.R. Storm. 1978. Helminth ova in soil and sludges from twelve U.S. urban áreas. J. Water Pollut. Control Fed. 50: 2485-2493. [ Links ]
Thornton, J.E., R.R. Bell & M.J. Reardon. 1974. Internal parasites of coyotes in Southern Texas. J. Wildl. Dis. 10: 232-236. [ Links ]
Torres, J., M.J. Perez, J.M. Segovia & J. Miquel. 2001. Utilidad de la coprologia parasitaria en la deteccion de helmintos parásitos en los cánidos silvestres ibericos.
Galemys 13: 75-83. [ Links ]
Trout, J.M., M. Santin & R. Fayer. 2006. Giardia and Cryptosporidium species and genotypes in coyotes (Canis latrans). J. Zoo Wildl. Med. 37: 141-144. [ Links ]
Van Den Bussche, R.A., M.L. Kennedy & W.E. Wilhelm. 1987. Helminth parasites of the coyote (Canis latrans) in Tennessee. J. parásitol. 73: 327-332. [ Links ]
Vaughan, C. 1983. Coyote range expansion in Costa Rica and Panama. Brenesia 21: 27-32. [ Links ]
Vaughan, C. & M. Rodriguez. 1986. Comparacion de los habitos alimentarios del coyote (Canis latrans) en dos localidades en Costa Rica. Vida Silvestre Neotropical 1: 6-11. [ Links ]
VerCauteren, K.C., T.C. Atwood, T.J. DeLiberto, H.J. Smith, J.S. Stevenson, B.V. Thomsen, T. Gidlewski & J. Payeur. 2008. Sentinel-based surveillance of coyotes to detect bovine tuberculosis, Michigan. Emerg. Infect. Dis. 14: 1862-1869. [ Links ]
Zunino, M.G., M.V. De Francesco, J.A. Kuruc, N. Schweigmann, M.C. Wisnivesky-Colli & O. Jensen. 2000. Contaminacion por helmintos en espacios públicos de la provincia de Chubut, Argentina. Bol. Chil. parásitol. 55: 78-83. [ Links ]
*Correspondencia a: Carmen Niehaus: Escuela de Medicina y Cirugía Veterinaria San Francisco de Asís, Universidad Veritas, carretera a Las Nubes, frente a los tanques de AyA, San Rafael de Coronado, San José, Costa Rica. Apartado 294-2200; carmen.niehaus@gmail.com
Idalia Valerio & Misael Chinchilla: Departamento de Investigación y Cátedra de parásitología, Universidad de Ciencias Medicas, San José, Costa Rica; valerioci@ucimed.com, chinchillacm@ucimed.com
Kinndle Blanco: Escuela de Medicina Veterinaria, Universidad Nacional, Heredia, Costa Rica; kblanco@una.ac.cr
1. Escuela de Medicina y Cirugía Veterinaria San Francisco de Asís, Universidad Veritas, carretera a Las Nubes, frente a los tanques de AyA, San Rafael de Coronado, San José, Costa Rica. Apartado 294-2200; carmen.niehaus@gmail.com
2. Departamento de Investigacion y Catedra de Parasitologia, Universidad de Ciencias Medicas, San José, Costa Rica; valerioci@ucimed.com, chinchillacm@ucimed.com
3. Escuela de Medicina Veterinaria, Universidad Nacional, Heredia, Costa Rica; kblanco@una.ac.cr
Recibido 22-III-2011. Corregido 25-VIII-2011. Aceptado 27-IX-2011.


{kind=link}
{kind=link}