










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Coadaptación entre los ácaros (Arachnida: Klinckowstroemiidae) y coleópteros Passalidae (Insecta: Coleoptera)
*Dirección para correspondencia:
Abstract
Mites of the family Klinckowstroemiidae establish an association with beetles of the family Passalidae known as phoresy. In order to obtain information about this association, we analyzed the relationship between mites of the family Klinckowstroemiidae and beetles of the family Passalidae, as adult mites have been exclusively collected from host beetles. We examined 1 150 beetles collected in seven states of the Mexican Republic, and found 19 species of klinckowstroemiid mites associated with 168 passalids, that belong to 28 different species in 15 genera. Host specificity between species of both groups does not exist, as one species of passalid beetle can have several different symbionts; conversely, a given mite species can associate with passalid beetles of different species and even of different genera. This way, Odontotaenius zodiacus has been found associated with mites of seven species of the genus Klinckowstroemia. Besides, Klinckowstroemia valdezi is associated with five species of passalids. Furthermore, two and even three different species of mites have been found on one host beetle (synhospitality). The lack of congruence between the phylogenies of the mites and that of the beetles indicates that a process of co-adaptation by colonization is going on, because the association is due to the resources that passalid beetles can offer to the mites, like transportation, food and refuge. Since these resources are not host-specific, the klinckowstroemiid mites can climb onto virtually any species of passalid beetles occurring on the same habitat. Rev. Biol. Trop. 60 (2): 599-609. Epub 2012 June 01.
Key words: Klinckowstroemia, co-adaptation, symbiosis, phoresy, synhospitality, Passalidae.
Resumen
Realizamos un análisis de la relación que existe entre los ácaros Klinckowstroemiidae y los coleópteros Passalidae, puesto que los ácaros adultos se recolectaron exclusivamente sobre pasálidos (huéspedes). Asimismo, examinamos 1 150 coleópteros recolectados en siete estados de la República Mexicana, y encontramos 19 especies de ácaros klinckowstroémidos asociados con 168 de ellos, pertenecientes a 28 diferentes especies de 15 géneros distintos. Estos táxones establecen una relación simbiótica llamada foresia. No existe una especificidad entre las especies de ambos grupos, debido a que una especie de pasálido puede tener varios forontes, y una especie de ácaro puede asociarse a pasálidos de diferentes géneros y especies. Un ejemplo es Odontotaenius zodiacus que presentó a siete especies de Klinckowstroemia asociadas, y por otro lado, Klinckowstroemia valdezi se encuentra asociada a cinco especies de pasálidos. Además, dos o hasta tres especies de ácaros se encontraron sobre un coleóptero huésped (synhospitality). La falta de congruencias entre la filogenia de los ácaros con la de los pasálidos nos indica que se está produciendo un proceso de coadaptación por colonización, y que ésta asociación se debe a los recursos que le brindan los pasálidos a los ácaros: transporte, refugio y alimento. Como estos recursos no son específicos para algún huésped los klinckowstroémidos se suben a cualquier especie de pasálido.
Palabras clave: Klinckowstroemia, coadaptación, simbiosis, foresia.
Los organismos se relacionan entre sí, de tal manera que estableciendo diferentes tipos de simbiosis, la cual es una relación inter específica y prolongada, en la que dos especies viven juntas en una asociación estrecha. Esta asociación ecológica implica transferencia de energía o algún beneficio adaptativo (Emmel 1975). En esta se involucran dos organismos: el huésped, que es el organismo de mayor tamaño y el simbionte de menor talla (Smith & Douglas 1987).
Los ácaros de la familia Klinckowstroemiidae establecen con los coleópteros de la familia Passalidae, una asociación conocida como foresia. Esta se define como una asociación no parásita entre un huésped (individuo de mayor tamaño) que transporta al foronte (individuo de menor tamaño). Esta asociación incluye: 1) la foresia pasiva, cuando el foronte se coloca debajo o dentro de alguna estructura o cavidad del huésped y no invierte mayor energía al ser transportado; y 2) la foresia activa, cuando el foronte se sostiene o sujeta de alguna estructura del cuerpo del huésped y por consecuencia si invierte energía (Athias-Binche 1994, Vachon 1940).
En algunos casos, la relación entre poblaciones de dos especies llega a ser tan estrecha que cada una de ellas ejerce una fuerza selectiva sobre la otra, por lo que en cada especie se originan respuestas adaptativas a esta interacción ecológica, conocida como coevolución (Ehrlich & Raven 1964). La evolución de las asociaciones ecológicas involucra una combinación de especiación y adaptación, que según Brooks & McLennan (1991) la coevolución involucra tanto el grado de adaptación filogenética mutua (coespeciación) como el grado de modificación mutua (coadaptación).
La coespeciación nos describe los patrones de asociación geográfica y/o ecológica entre los clados. Esta se puede desarrollar a través de dos procesos: 1) por descendencia, en donde los ancestros se asociaron entre sí en el pasado y las especies heredaron dicha asociación, y 2) por colonización, en donde una de las especies se originó en otro huésped y subsecuentemente se relacionaron por transferencia o dispersión.
La coadaptación se manifiesta en el grado en que las especies que coevolucionan se afectan o se han afectado genética o ecológicamente. Existen tres modelos para explicar la coadaptación: 1) la coespeciación alopátrida, 2) la colonización y 3) la exclusión (Morrone 2001). La coespeciación alopátrida constituye la hipótesis nula para los estudios de coadaptación, en donde se predice que habrá congruencia entre las filogenias de los huéspedes y los asociados; la asociación entre dichos linajes se debe a la presencia de algún recurso en los huéspedes que resulta de utilidad para los asociados. La colonización se basa en la idea de que los huéspedes representan recursos que los asociados han rastreado en el curso del tiempo evolutivo. En este modelo, ya sea porque el recurso (huésped) estaba ampliamente distribuido antes de la diversificación del asociado o porque fue colonizado en eventos independientes (convergentemente), no habrá congruencia entre los patrones filogenéticos de huéspedes y asociados. Finalmente, la exclusión consiste en que la asociación se mantiene por respuestas adaptativas recíprocas, en que algún carácter del huésped lo protege de los efectos del asociado.
Actualmente, la familia Klinckowstroemiidae contiene cuatro géneros y 36 especies (Villegas-Guzmán 2010); se conocen solamente los adultos y estos se han recolectado exclusivamente asociados con coleópteros de la familia Passalidae. Los pasálidos tienen hábitos subsociales y es común encontrar una pareja, macho y hembra, los cuales cohabitan con su prole (larvas, pupas y adultos tenerales), en una red de galerías dentro de un tronco en descomposición.
El objetivo de este trabajo es determinar si existe algún tipo de proceso coadaptativo o coevolutivo que explique la relación simbiótica entre los ácaros Klinckowstroemiidae y los coleópteros Passalidae.
Materiales y métodos
Se recolectaron pasálidos dentro y debajo de troncos en descomposición, todas las recolectas fueron diurnas; se realizaron entre las 10:00 y 18:00 horas, en siete estados de la República Mexicana: Hidalgo (22 y 23 septiembre 2006), Oaxaca (22 noviembre 1996, 13 noviembre 2005, 16, 15, 24 y 28 junio 2006, 20 y 22 julio 2007), Chiapas (10 agosto 2005, 4 octubre 2005, 8 mayo 2006, 9 agosto 2006, 19 mayo 2007, 7 junio 2008) , Veracruz (11 octubre 1982, 14 y 15 noviembre 1987, 23 y 30 septiembre 2006, 23 marzo 2007), Yucatán (9 julio 2007), Guerrero (16 junio2007) y Quintana Roo (7 julio 2007). Los coleópteros fueron colocados individualmente (un pasálido por frasco) en frascos con alcohol etílico al 80%. En los primeros muestreos, los pasálidos de una misma localidad fueron puestos en un solo frasco; sin embargo, notamos que muchos se desprendían de su huésped, lo cual imposibilitó en un principio establecer con precisión la relación simbionte huésped, con el fin de dilucidar las asociaciones específicas. Además, los pasálidos de un tronco fueron marcados como pertenecientes a un grupo familiar y se manejaron separados de otros grupos familiares de la misma localidad, para establecer patrones de asociación ácaro-pasálido no solamente a nivel individual, sino a nivel de un grupo familiar o grupo social. Para obtener la prevalencia y la intensidad promedio se siguió a Bush et al. (1997). No se realizó un método de muestreo específico, sino que se buscaron troncos en descomposición, que es donde habitan los pasálidos, en diferentes localidades. Chiapas: Ejido Benito Juárez (15°05.966’ N - 92°08.600’ W), 1 687m. Hidalgo: 2km carretera Federal crucero Tianguistengo (20°40.413’ N - 98°40.255’ W), 2 080m. 2km Mojonera-Tizapan, Zacualtipan (20°38.107’ N- 98°35.981’ W), 2 019m. 2km E Acaxochitlán (20°08.500’ N - 98°10.740’ W), 2 214m. Ejido Atepixco, Zacualtipan (20°35.466’ N-98°36.358’ W), 1 972m. Oaxaca: 8km NW San Miguel Coatlán (16°15.064’ N - 96°41.629’W), 2 170m. 101km carretera federal Puerto Escondido-Oaxaca (16°28.156’ N - 97°01.674’W), 1 906m. Ixtlán de Juárez (17°31.722’ N - 96°30.678’ W), 2 742m. Cerro Pelón, Carretera Tuxtepec-Oaxaca (17°35.75’ N - 96°28.41’W), 2 240 m. 8km SE San Miguel Talea de Castro (17°19.620’ N - 96°17.403’ W), 2082m. 7km E San Pedro-San Pablo Ayutla (17°01.037’ N - 96°01.915’ W), 2 459m. 10km NE Mirador Santiago Comaltepec (17°35.293’N - 96°27.162’ W), 2 188m. Camino a El Porvenir (17°18.160’ N - 96°14.423’ W), 1 870m. Veracruz: 1.5km N Viborillas, Huayacocotla (20°31.245’ N - 98°29.293’ W), 2328m. 11.9km N Huayacocotla (20°32.116’ N - 98°28.783’ W). 14.6km N Huayacocotla (20°32.116’ N - 98°28.783’ W). Dos Amates, Catemaco (18º29.35’ N - 94º56.36’ W), 120m. Coyame Bastonal, Catemaco (18°23.25’ N - 94°28.783’ W), 830-970m. Soledad Atzompa (18°44.712’ N - 97°40.967’ W), 2 125m. Tlacolulan (19°38.508’ N - 96°59.650’ W), 1 876m.
Los ejemplares fueron transportados a la Colección Nacional de Ácaros (CNAC) del Instituto de Biología de la UNAM, donde los ácaros fueron removidos con pinzas de punta fina bajo el microscopio estereoscópico. Los ácaros fueron almacenados en frascos con alcohol al 80%. Algunos de ellos fueron aclarados con lacto-fenol y montados con líquido de Hoyer para su posterior identificación. Los pasálidos fueron depositados en la Colección Entomológica del Instituto de Ecología, Xalapa (IEXA), donde otros ejemplares depositados fueron revisados para buscar klinckowstroémidos y otros ácaros. Los ácaros están depositados en la CNAC. Los coleópteros y ácaros fueron determinados hasta nivel específico con ayuda de bibliografía especializada.
Resultados
Se revisaron un total 1 150 pasálidos, de los cuales solo 758 presentaron ácaros. Los ejemplares fueron recolectados de 167 localidades, provenientes de siete estados de la República Mexicana: 65 de Hidalgo, 51 de Oaxaca, 27 de Chiapas, 19 de Veracruz, 2 de Yucatán, 2 de Guerrero y 1 de Quintana Roo. Dichos huéspedes pertenecen a 53 especies de 15 géneros de Passalidae. Del total de huéspedes analizados solamente se encontraron klinckowstroémidos adultos en 168 pasálidos (14.6%), pertenecientes a 28 especies de 15 géneros.
La especie con la mayor cantidad de ejemplares con ácaros fue Odontotaenius zodiacus (Truqui) con una prevalencia del 75% y una intensidad promedio de 9.02, seguida por Proculejus hirtus (Truqui) con 73% y 5.88, respectivamente (Cuadro 1). Estos valores dependen del número de coleópteros con ácaros, debido a la observación que se hizo en los pasálidos con pocos ejemplares revisados [Pseudacanthus sp. y Proculus goryi (Melly)] presentaron los mayores valores de prevalencia e intensidad promedio, puesto que todos los ejemplares revisados tenían ácaros en abundante cantidad.
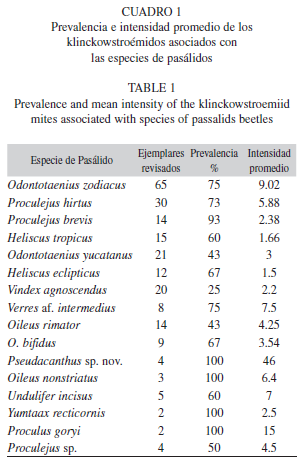
En Odontotaenius zodiacus se encontraron siete especies de klinckowstroémidos, mientras que en Proculejus brevis y Heliscus eclipticus se encontraron cuatro y tres especies, respectivamente. Verres af. intermedius, Proculejus hirtus, Oileus nonstriatus, O. rimator, O. bifidus y Pseudacanthus sp., presentaron dos especies de klinckowstroémidos cada una. En los 168 pasálidos fueron encontrados 1 200 ácaros (621♀♀ y 579 ♂♂) de Klinckowstroemia Baker & Wharton (Villegas-Guzmán 2010) con un promedio de 7.2 ácaros por pasálido. La abundancia de ácaros forontes varía de uno a 41 por huésped. En siete huéspedes se concentraron más del 62% de las especies de klinckowstroémidos encontradas: Odontotaenius zodiacus (41%), Proculejus hirtus (8.5%), Verres af. intermedius (3.7%), Oileus bifidus (3.4%), Proculus goryi (2.5%), Odontotaenius yucatanus (1.7%) y Proculejus brevis (1.6%). El número de forontes encontrado en estas especies, tiene relación con el número de huéspedes revisado, O. zodiacus y P. hirtus fueron las especies con el mayor número de ejemplares revisado (Cuadro 1).
La mayoría de los ácaros se encontraron en el alcohol (80.4%) donde fueron colocados los pasálidos, sin embargo, algunos coleópteros aún conservaban ácaros adheridos en diferentes partes del cuerpo (19.6%): en el área de los húmeros, en la región ventral del cuerpo cercana a las coxas II y III, en la base del pronoto y en el submentón (Cuadro 2); así como en el detritus que mantiene el pasálido entre sus mandíbulas. Debido a que la mayoría de los ácaros se encontraron en el alcohol, es muy probable que estos estaban realizando foresia activa, es decir, se sujetaban con sus patas a las sedas o alguna estructura del cuerpo del coleóptero y por lo que al ser fijados con sus huéspedes, se desprendieron fácilmente de los sitios de anclaje.
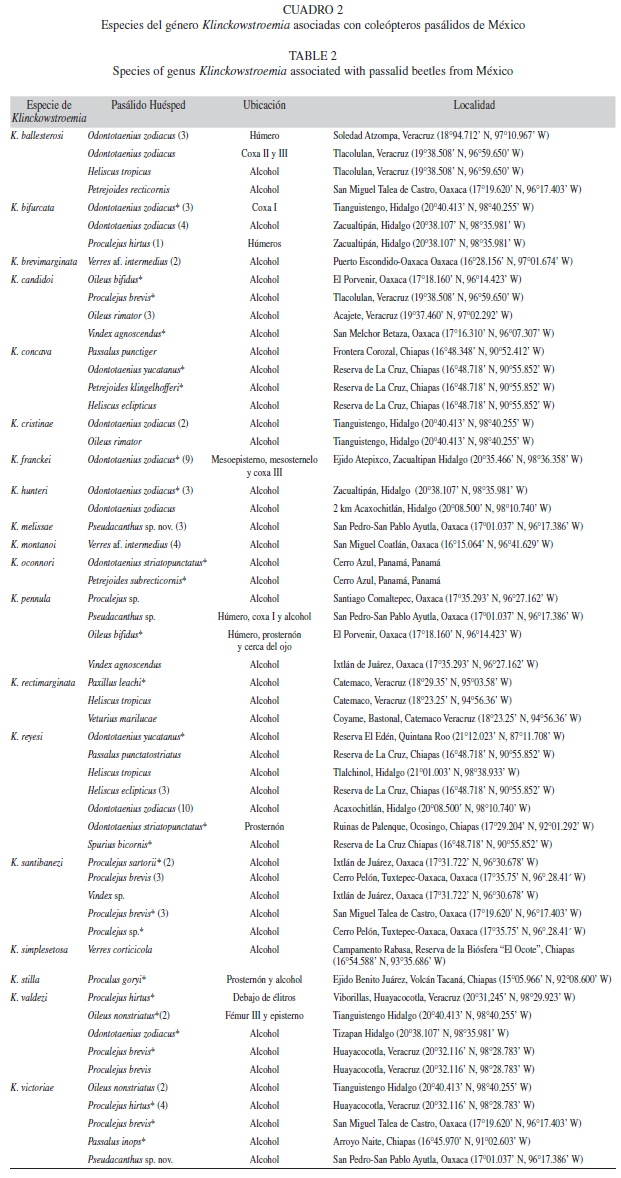
En total se identificaron 19 especies de Klinckowstroemia, de las cuales 14 resultaron ser nuevas para la ciencia (Villegas-Guzmán et al. 2009, Villegas-Guzmán et al. 2011) y cinco son especies previamente descritas. En 18 de los 168 pasálidos revisados, se encontraron dos especies de Klinckowstroemia por hospedero, esta asociación es conocida como synhospitality, que consiste cuando dos o más especies relacionadas se encuentran juntas en el mismo huésped (Eichler 1966). El término synhospitality se ha usado previamente en ácaros demodícidos parásitos asociados a mamíferos (Nutting 1979) y en acáridos foréticos asociados con abejas (Ochoa & Oconnor 2000).
Las especies de Klinckowstroemia están organizadas en dos grupos basados en el tamaño: el de las especies grandes y el de las especies pequeñas. En los casos de synhospitality, generalmente se encontraron juntas una especie pequeña (650-870μm) y una grande (900-1 350μm). En seis de estos, la especie de menor tamaño fue Klinckowstroemia victoriae Rosario & Hunter. En un otro se encontraron Klinckowstroemia bifurcata Villegas-Guzmán et al., K. hunteri Villegas-Guzmán et al. y K. valdezi Villegas-Guzmán et al., asociados con el mismo ejemplar de Odontotaenius zodiacus, las dos primeras son de tamaño similar (grande), mientras que la última es de tamaño pequeño (Cuadro 3). Otro ejemplo de esta relación es el grande K. candidoi Rosario & Hunter y el pequeño K. pennula Villegas-Guzmán et al., los cuales fueron encontrados asociados con Oileus bifidus. Solamente en un caso se encontraron ácaros del mismo tamaño juntos, ambos pequeños, K. pennula y K. melissae Villegas-Guzmán et al., asociados a un ejemplar de Pseudacanthus sp.
En diez ocasiones se recolectaron pasálidos de un grupo familiar. En una misma localidad se encontraron dos familias de P. brevis, una con dos ejemplares y la otra con cuatro. En el primer caso, los dos ejemplares tenían sobre ellos a K. santibanezi; en la otra familia tres de los cuatro ejemplares estaban asociados a la misma especie de ácaro. En otra familia, se encontraron seis ejemplares de O. bifidus (Cuadro 3), cinco de los cuales presentaron K. candidoi (especie grande) y K. pennula (especie pequeña). Estas observaciones nos indican lo siguiente: los pasálidos que viven juntos, i. e. un grupo familiar, comparten las mismas especies de ácaros sobre ellos, o, que los ácaros que ocupan una red de galerías aprovechan a los pasálidos que las construyeron para dispersarse a un hábitat nuevo.
En algunas ocasiones las familias de pasálidos no presentaron ácaros Klinckowstroemiidae, que son de nuestro particular interés. Sin embargo, sí presentaron y compartieron ácaros de Megisthaniidae, Euzerconiidae, Uropodidae, Diarthrophallidae y Diplogyniidae, los cuales han sido previamente registrados sobre varias especies de pasálidos (Hunter 1993, Villegas-Guzmán et al. 2008). Tal es el caso de la familia compuesta por dos ejemplares de V. agnoscendus donde ambos compartían los ácaros de las familias Diplogyniidae y Diarthrophallidae.
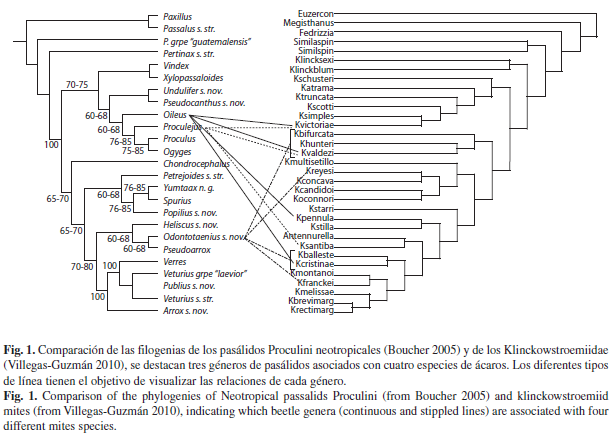
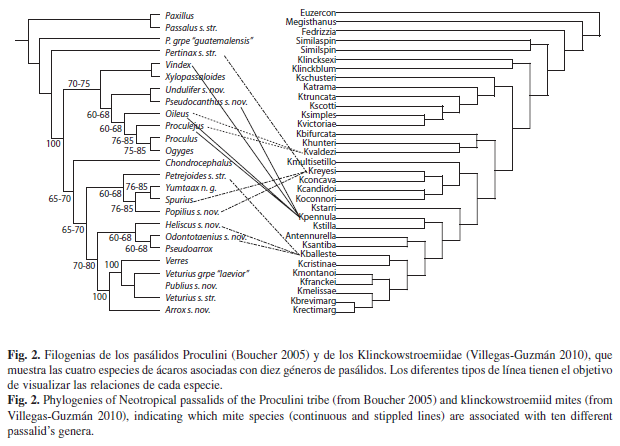
Al comparar la filogenia de los géneros neotropicales de Proculini y de Passalini (Boucher 2005) con la filogenia de la familia Klinckowstroemiidae (Villegas-Guzmán 2010), encontramos que no hay congruencia entre la filogenia de los huéspedes (coleópteros) y la de los simbiontes (ácaros). Algunos pasálidos pueden presentar diferentes especies de klinckowstroémidos asociados. Un ejemplo es Odontotaenius con siete especies de ácaros asociadas las cuales no pertenecen a un clado monofilético, y otro es Oileus que presenta cinco especies de ácaros asociadas y estas tampoco forman un clado monofilético (Fig. 1). Por otra parte, una especie de klinckowstroémido puede tener diferentes huéspedes: K. ballesterosi y K. valdezi tienen como huéspedes a tres géneros de pasálidos, que no pertenecen a un clado monofilético (Fig. 2), mientras que K. pennula se encuentra asociada con cuatro géneros de pasálidos que tampoco están cercanamente relacionados entre sí, así como K. reyesi, el ácaro con el mayor número de huéspedes, con cinco géneros (Fig. 2).
Discusión
Los adultos de las especies de Klinckowstroemia habitan en las madrigueras de los pasálidos, los estados inmaduros se desconocen, donde encuentran las condiciones adecuadas para su desarrollo, además emplean al huésped como su medio de dispersión, con el que establecen una relación forética. Ahora bien, se encontró que no existe especificidad entre una especie de ácaro con alguna especie de pasálido, sino que una especie de Klinckowstroemia puede encontrarse en diferentes especies de huésped: ejemplo de ello son K. valdezi, Klinckowstroemia ballesterosi Villegas-Guzmán et al., Klinckowstroemia santibanezi Villegas-Guzmán et al., y K. pennula, cada una de ellas encontrada en cuatro especies de huésped, mientras que Klinckowstroemia rectimarginata Villegas-Guzmán et al., y Klinckowstroemia reyesi Rosario y Hunter están asociadas con tres y siete especies de pasálidos respectivamente (Cuadro 2), mientras que Klinckowstroemia concava Rosario y Hunter, y K. victoriae, se han encontrado asociadas con cinco.
También ocurre que en una especie de huésped pueden encontrarse varias especies de klinckowstroémidos, en Odontotaenius zodiacus se hallaron a siete especies (Cuadro 2); mientras que en Proculejus brevis y Heliscus eclipticus se descubrieron cuatro y tres especies de ácaro, respectivamente. Las especies de Klinckowstroemia se encontraron asociadas con 28 especies de pasálidos, la mayoría de ellos perteneciente a Proculini (24 especies) y el resto a Passalini (4 especies). Seis de estas especies de pasálidos habían sido previamente referidas como huéspedes del género Klinckowstroemia, y el resto (22 especies) representan nuevos registros de huéspedes; O. rimator registrado previamente como huésped de Klinckowstroemia grabowskii Chernoff & Pope, es un nuevo huésped para Klinckowstroemia scotti Rosario & Hunter y Klinckowstroemia candidoi Rosario & Hunter.
Por lo anterior, se concluye que no existe especificidad entre las especies de ácaros Klinckowstroemiidae y los coleópteros Passalidae. Los ácaros establecen la relación simbiótica con los pasálidos por los recursos generalizados que estos le brindan: transporte, alojamiento y posiblemente comida, y no específicamente por el huésped. Sin embargo, la relación simbiótica está restringida a nivel de estas dos familias de artrópodos, los adultos de las especies conocidas de Klinckowstroemiidae solamente están asociados con coleópteros adultos de Passalidae. Además, se desconocen los estados inmaduros de estos ácaros, pero no se descarta que dependan de alguna forma del contenido que los pasálidos generan dentro de sus galerías (madrigueras), lo que vendría a reforzar la relación forética establecida entre estos dos grupos de organismos.
Debido a que los pasálidos tienen hábitos subsociales, se puede encontrar en su nido a una pareja de adultos con toda su descendencia. Lo cual ocasiona que los adultos integrantes del grupo familiar pueden compartir sus ácaros asociados. Esto no es válido para los integrantes de un grupo familiar, únicamente, sino también para los pasálidos habitantes del tronco, puesto que se conoce que un mismo tronco puede estar habitado por cuatro o cinco especies de pasálidos (Reyes-Castillo & Halffter 1984). Por ejemplo, en un tronco se encontraron tres ejemplares, dos ejemplares de Pseudacanthus sp., dónde ambos coleópteros presentaron a K. pennula y K. melissae (ambas pequeñas) (Cuadro 3); mientras que el otro pasálido, Vindex agnoscendus, probablemente habitaba en una galería cercana, pero independiente no presentó klinckowstroémidos. Sin embargo, los tres pasálidos presentaron debajo de los élitros y en las alas membranosas a ácaros de la familia Diarthrophallidae. Esto indica que probablemente, por el hecho de habitar el mismo tronco, los ácaros pueden trasladarse de una galería a otra y de esta forma pasan de una especie de pasálido a otra, por lo que una especie de Klinckowstroemia puede encontrarse en diferentes especies de pasálidos.
Consideramos que el cuidado parental que realizan los pasálidos es crucial para la interacción con los klinckowstroémidos y otros ácaros, ya que es la forma mediante la cual los ácaros pasan de un tronco en descomposición a otro, cuando los pasálidos adultos jóvenes abandonan las galerías paternales y van a formar un nuevo grupo familiar. El macho y la hembra que establecen un nuevo grupo familiar pueden aportar ácaros foréticos diferentes a la nueva red de galerías donde sus larvas se desarrollan; las cuales a su vez, al madurar transportan otra “mezcla” de ácaros que contaminan sus propias galerías y su prole.
El hecho de que las filogenias ácaro y pasálido no guarden congruencia indica que no se está llevando a cabo un proceso coevolutivo sino uno de coadaptación por colonización. Esto es, que la asociación entre los ácaros y los coleópteros se debe a los recursos que le brindan los pasálidos a los klinckowstroémidos (transporte, refugio y posiblemente alimento), razón por la cual éstos pueden subirse indistintamente a cualquier especie de pasálido, porque todas tienen el mismo comportamiento subsocial, lo cual propicia que diferentes ácaros se desarrollen en sus madrigueras (Hunter 1993, Villegas-Guzmán et al. 2008). Los ácaros están realizando la foresia activa sobre los pasálidos, esto es evidente al encontrar solamente adultos; de acuerdo con Athias-Binche (1994) son éstos, en especial las hembras, los que establecen la relación forética con sus huéspedes. Sin embargo, en el caso de los klinckowstroémidos adultos recolectados, la proporción sexual es cercana a uno (621♀♀ y 579 ♂♂).
La distribución de las especies de Klinckowstroemia se encuentran en un intervalo altitudinal entre 1 680-2 750ms, con excepción de K. rectimarginata y K. oconnori Villegas-Guzmán et al., esta última de Panamá, que se encuentran en altitudes menores de los 1 000m. Los klinckowstroémidos mexicanos tienen un patrón de distribución mesoamericano de montaña, lo que coincide con la distribución de sus especies huésped, que en su mayoría pertenecen a Proculini, grupo predominantemente montano a nivel genérico (Halffter, 2006). La mayoría de las especies de la tribu habitan el bosque mesófilo de montaña, hábitat típico donde se presenta un conjunto significativo de especies endémicas de pasálidos, propias en varios de los sistemas montañosos mexicanos (Reyes-Castillo et al. 2006). Consideramos que se deben realizar estudios que profundicen en el conocimiento de las relaciones entre pasálidos y ácaros, así como realizar la búsqueda de los estados inmaduros de los ácaros Klinckowstroemia, que nos ayuden a determinar su ciclo de vida.
Agradecimientos
Los autores agradecen a Jesús Ballesteros, Héctor Montaño, Carlos Santibañez y Alejandro Valdez por la recolección de los coleópteros. Este estudio fue apoyado por el CONACYT y el Posgrado en Ciencias Biológicas de la UNAM. Las salidas al campo fueron apoyadas por el proyecto NSF BIO-DEB 0413453 Lorenzo Prendini y por el Instituto Bioclon, México, D. F. Se recolectó material bajo el amparo de la Licencia de Colector Científico (FAUT-0175) expedida por la SEMARNAT a O. F. Francke. A tres revisores anónimos que con sus acertados comentarios enriquecieron este trabajo.
Referencias
Athias-Binche, F. 1994. La phorésie chez les acariens. Aspects adaptatifs et Evolutifs. Castillet, Perpignan, Paris, Francia. [ Links ]
Boucher, S. 2005. Évolution et phylogénie des coléoptères Passalidae (Scarabaeoidea). Ann. Soc. Entomol. Fr. (n. s.) 41: 239-604. [ Links ]
Brooks, D.R. & D.A. McLeennan. 1991. Phylogeny, ecology, and behavior. A research program in comparative biology. The University Chicago, Chicago, USA. [ Links ]
Bush, A.O., K.D. Latterty, J.M. Lotz & A.W. Shostak. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 83: 575-583. [ Links ]
Emmel, T.C. 1975. Ecología y biología de poblaciones. Interamericana McGraw-Hill, México, D.F. [ Links ]
Eichler, W. 1966. Two new evolutionary terms for speciation in parasitic animals. Syst. Zool. 15: 216-218. [ Links ]
Ehrlich, P.R. & P.H. Raven. 1964. Butterflies and plants: a study in coevolution. Evolution 18: 586-608. [ Links ]
Halffter, G. 2006. Biogeografía de la entomofauna de montaña de México y América Central, p. 1-21. In J.J. Morrone & J. Llorente Bousquets (eds.). Componentes Bióticos principales de la Entomofauna Mexicana. Las Prensas de Ciencias, UNAM, México, D.F. [ Links ]
Hunter, P.E. 1993. Mites associated with New World passalid beetles (Coleoptera: Passalidae). Acta Zool. Mex. (n.s.) 58: 1-37. [ Links ]
Morrone, J.J. 2001. Sistemática, biogeografía, evolución. Los patrones de biodiversidad en tiempo-espacio. Las Prensas de Ciencias, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Nutting, W.B. 1979. Synhospitaly and speciation in the Demodicidae (Trombidiformes), 267-272. In E. Piffl (ed.). Proceedings of the 4th International Congress of Acarology. Akadémia Kiadó, Budapest. [ Links ]
Ochoa, R. & B.M. O’Connor. 2000. Revision of the genus Horstiella (Acari: Acaridae): mites associated with Neotropical Epicharis bees (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 93: 713-737. [ Links ]
Reyes-Castillo, P. & G. Halffter. 1984. Estructura social de los Passalidae (Coleoptera: Lamelicornia). Folia Entomol. Mex. 61: 49-72. [ Links ]
Reyes-Castillo, P., C.V. Rojas-Gómez & H. Vázquez. 2006. Patrones de distribución de la familia Passalidae (Coleoptera), p. 237-270. In J.J. Morrone & J. Llorente Bousquets (eds.). Componentes Bióticos principales de la Entomofauna Mexicana. Las Prensas de Ciencias, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Smith, D.C. & A.E. Douglas. 1987. The biology of Symbiosis. Edward Arnold, Londres, Inglaterra. [ Links ]
Vachon, M. 1940. Remarques sur la phorésie des pseudoscorpions. Ann. Soc. Entomol. Fr. 109: 1-18. [ Links ]
Villegas-Guzmán, G.A. 2010. Revisión taxonómica y análisis filogenético del género Klinckowstroemia Baker y Wharton (Acarina: Mesostigmata: Trigynaspida: Klinckowstroemiidae). Tesis de Doctorado, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Villegas-Guzmán, G.A., T.M. Pérez & P. Reyes-Castillo. 2008. Ácaros asociados al coleóptero Passalus cognatus (Coleoptera: Passalidae) de los Tuxtlas, Veracruz, México. Rev. Biol. Trop. 56: 1261-1268. [ Links ]
Villegas-Guzman, G.A., T.M. Pérez & P. Reyes-Castillo. 2009. New species of the genus Klinckowstroemia Baker & Wharton from México (Acari: Mesostigamata: Klinckowstroemiidae). Zootaxa 2248: 1-46. [ Links ]
Villegas-Guzmán, G.A., P. Reyes-Castillo & T.M. Pérez. 2011. New species of Klinckowstroemia Baker & Wharton, 1952 (Acari: Klinckowstroemiidae), associated with passalid beetles from México and Panama. Zootaxa 2732: 1-17. [ Links ]
*Correspondencia a: Gabriel A. Villegas-Guzmán &Tila M. Pérez: Colección Nacional de Ácaros, Departamento Zoología, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado Postal 70-153, Ciudad Universitaria, 04510 México, D.F.; gabrvill@yahoo.com, tilam@ibiologia.unam.mx
Oscar F. Francke: Colección Nacional de Arácnidos, Departamento Zoología, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado Postal 70-153, Ciudad Universitaria, 04510 México, D.F.; offb@ibiologia.unam.mx
Pedro Reyes-Castillo: Instituto de Ecología, A.C. Apartado Postal 63, 91000 Xalapa, Veracruz, México; pedro.reyes@inecol.edu.mx
Gabriel A. Villegas-Guzmán: Laboratorio de Entomología, Escuela Nacional de Ciencias Biológicas, IPN, Departamento de Zoología, Prolongación de Carpio y Plan de Ayala s/n, Col. Santo Tomás, C.P. 11340, México D.F.
1. Colección Nacional de Ácaros, Departamento Zoología, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado Postal 70-153, Ciudad Universitaria, 04510 México, D.F.; gabrvill@yahoo.com, tilam@ibiologia.unam.mx
2. Colección Nacional de Arácnidos, Departamento Zoología, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado Postal 70-153, Ciudad Universitaria, 04510 México, D.F.; offb@ibiologia.unam.mx
3. Instituto de Ecología, A.C. Apartado Postal 63, 91000 Xalapa, Veracruz, México; pedro.reyes@inecol.edu.mx
Correspondencia. Dirección actual: Laboratorio de Entomología, Escuela Nacional de Ciencias Biológicas, IPN, Departamento de Zoología, Prolongación de Carpio y Plan de Ayala s/n, Col. Santo Tomás, C.P. 11340, México D.F.
Recibido 04-IV-2011. Corregido 09-IX-2011. Aceptado 04-X-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}