










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Variabilidad genética de poblaciones en cautiverio de Crocodylus moreletii (Crocodylia: Crocodylidae) mediante el uso de marcadores microsatelitales
*Dirección para correspondencia
Abstract
Genetic variability in captive populations of Crocodylus moreletii (Crocodylia: Crocodylidae) using microsatellites markers. Crocodylus moreletii, an extinction threatened species, represents an emblem for tropical ecosystems in Mexico. Surprisingly, there is a lack of information about their genetic constitution, which should be evaluated for a proper management ex situ and for making decisions on the release of crocodiles into natural habitats. The aim of this study was to characterize and compare the genetic variability of four populations of C. moreletii (two wild versus two born ex situ). Through PCR were amplified seven microsatellite polymorphic loci, however a heterozygote deficit, diminished by the presence of null alleles, was found in the populations (average HO=0.02). The AMOVA indicated that the highest proportion of genetic variability is within populations, and a limited genetic differentiation among populations (average FST=0.03), probably due to high inbreeding index (average FIS=0.97). When comparing the genetic variability between and within other crocodilian species, we found that in C. moreletii is well below those reported. We concluded that the limited genetic variability in ex situ born populations is probably due to a founder effect derived from the social structure of their progenitors, and by the bottleneck effect, inferred by the limited effective population size, that historically
characterizes their natural distribution in wild populations.
Key words: endogamous crocodile, homozygous crocodile, Unit of Management for Conservation of the Wild
Resumen
Crocodylus moreletii representa un emblema para los ecosistemas tropicales de México pero actualmente está amenazada por extinción. Sorprendentemente, hay una falta de información de su constitución genética, que debe ser evaluada para un manejo apropiado ex situ y para toma de decisiones en la liberación de cocodrilos a su hábitat natural. El objetivo del estudio fue caracterizar y comparar la variabilidad genética de cuatro grupos poblacionales de C. moreletii (dos silvestres y dos nacidas ex situ). Mediante PCR se amplificaron siete loci de microsatélites polimórficos, sin embargo se encontró déficit de heterocigotos en las poblaciones (promedio HO=0.02) mermado por la presencia de alelos nulos. El AMOVA indicó que la mayor proporción de variabilidad genética se encuentra dentro de las poblaciones y una limitada diferenciación genética entre poblaciones (promedio FST =0.03), probablemente debida al alto índice de endogamia (promedio FIS=0.97). Al comparar la variabilidad genética inter e intra especies de cocodrilianos, encontramos que en C. moreletii está muy por debajo de los reportados. Se concluye que la limitada variabilidad genética de las poblaciones nacidas ex situ probablemente se debe al efecto fundador derivado de la estructura social de sus progenitores, y de las poblaciones silvestres, por el efecto cuello de botella, inferido por el limitado tamaño efectivo de población que presentó históricamente en su distribución natural.
Palabras clave: cocodrilos endogámicos, cocodrilos homocigóticos, Unidad de Manejo para la Conservación de la Vida Silvestre, Conservación ex situ, Cocodrilo de Pantano, México.
Life, ex situ conservation, Morelet’s crocodile, Mexico. Los efectos de las actividades antropogénicas han impactado de manera negativa a la mayoría de los cocodrilianos (lagartos, caimanes y cocodrilos). Esto se ha reflejado en cambios en la estructura y dinámica poblacional, e incluso en la extinción de algunas de sus poblaciones (Ross 1998). Consecuentemente, doce especies de cocodrilianos están incluidas en alguna categoría de amenaza en la lista roja de la Unión Internacional para la Conservación de la Naturaleza (IUCN 2010). Ante tal situación, se han seguido dos estrategias directas de conservación. Por un lado, su protección se regula legalmente por los decretos de veda y a través de prácticas de mejoramiento y protección de sus hábitats declarándolos como zonas protegidas. Y por el otro, se ha optado por la conservación de estas especies a partir de su aprovechamiento ex situ (Luxmoore 1992). Esta última estrategia es con la finalidad a posteriori de recuperar poblaciones silvestres a través de la liberación de cocodrilos (Huchzermeyer 2003). Una de las prioridades en estos sistemas de crianza es la conservación de la variabilidad genética, ya que es necesario que las crías nacidas ex situ puedan resistir y adaptarse a las presiones ambientales del hábitat en el que son liberadas (FitzSimmons et al. 2002).
Actualmente, se han utilizado los marcadores moleculares para caracterizar el estado de la variabilidad genética de los cocodrilianos con fines de conservación (Glenn 2002, Forstner & Forstner 2002). Por ejemplo, AFLPs (Amplified Fragment Length Polymorphisms) en Alligator sinensis (Yang et al. 2006) y RAPD (Random Amplification of Polymorphic DNA) en A. sinensis (Wu et al. 2002), Caiman latirostris (Amavet et al. 2007) y Crocodylus acutus (Porras et al. 2008). Además se han empleado los marcadores de microsatélites (STR: Short Tandem Repeats; Tautz et al. 1986), y se ha determinado que éstos han detectado mejor el polimorfismo en lagartos (Davis et al. 2000), caimanes (Zucoloto et al. 2002, Zucoloto et al. 2006) y cocodrilos (FitzSimmons et al. 2000).
Las acciones enfocadas a la conservación mediante el aprovechamiento regulado de los cocodrilianos ex situ se realiza principalmente en sistemas de producción, en cautiverio y/o en granjas comerciales (Huchzermeyer 2003). Sin embargo, una de las problemáticas concurrentes en estos esquemas de aprovechamiento es la alteración de su variabilidad genética. En ciertos casos se ha demostrado mediante marcadores moleculares que poblaciones de A. sinensis en cautiverio presentan bajos niveles de variabilidad genética (Wu et al. 2002, Wang et al. 2006), al igual que poblaciones de C. niloticus manejadas en granjas comerciales (Flint et al. 2000, Bishop et al. 2009). En este contexto, la información sobre la variabilidad genética de cocodrilos bajo los diferentes esquemas de aprovechamiento ex situ sería complementaria para tomar decisiones que lleven a liberar individuos genéticamente viables en su hábitat natural (Yang et al. 2006), y de esa forma, recuperar las poblaciones donde históricamente se distribuían e implementar estrategias que preserven la variabilidad genética en las poblaciones en cautiverio (FitzSimmons et al. 2002).
Por muchos años, algunas poblaciones silvestres del Cocodrilo de Pantano (Crocodylus moreletii) distribuidas en las tierras bajas del Caribe Mexicano, fueron diezmadas por la caza ilegal y la pérdida de su hábitat (Álvarez del Toro & Sigler 2001), lo que llevó a catalogarla en amenazada por riesgo de extinción y dependiente de conservación (IUCN 2010). Ante ello, autoridades gubernamentales de México legislaron para su protección especial (DOF 2001), lo que ha favorecido su recuperación poblacional en vida libre (Domínguez-Laso 2006). Esto permitió sugerir su transferencia del Apéndice I al II de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestre para su utilización sostenible mediante rancheo en México (CITES 2008). Asimismo, el aprovechamiento reglamentado de C. moreletii ex situ es mediante esquemas de producción denominados Unidad de Manejo para la Conservación de la Vida Silvestre (UMA) (INE 2000), donde uno de los objetivo es la conservación y liberación de individuos a su hábitat natural (Casas 2002).
En C. moreletii aún no se logrado la liberación de individuos al medio. Hasta ahora, no existen estudios que analicen la variabilidad genética de las poblaciones cautivas en las UMAs establecidas en México, incluyendo la UMA donde se realizó este estudio. Esta información es importante para valorar el acervo genético de los cocodrilos que se pretendan liberar y que debe ser similar al que se encuentra en las poblaciones naturales. Dicha información también se puede utilizar para la planeación de acciones que garanticen mantener y/o incrementar la variabilidad genética de las poblaciones. Por estas razones, el objetivo del presente estudio fue caracterizar y comparar la variabilidad genética de cuatro grupos poblacionales de C. moreletii, dos que fueron extraídos de su hábitat silvestre con dos que nacieron ex situ, y que se encuentran actualmente bajo las mismas condiciones de cautiverio en una UMA establecida en Veracruz, México.
Materiales y métodos
Características de las poblaciones estudiadas: En la UMA Cacahuatal (INE/CITES/DFYFS-CRIN-0069-SIN/99), que se encuentra ubicada en la localidad José Ingenieros, municipio de La Antigua, Veracruz, México (19°22’19” N - 96°22’19” W), se mantienen reclutadas cuatro poblaciones de C. moreletii de diferente procedencia, cuyos individuos están marcados con placas interdigitales con el número clave de la población de origen. La primera y segunda población están conformadas por individuos capturados en su hábitat silvestre en los municipios de Tezonapa y Gutiérrez Zamora, ambas del estado de Veracruz. La tercera y cuarta población están constituidas por individuos F1 para explotación comercial, que nacieron y se criaron en UMAs establecidas en el municipio de Puerto Vallarta, Jalisco y Villa Juárez, Sinaloa, y que posteriormente fueron enviados a la UMA Cacahuatal. Los progenitores de estos últimos grupos poblacionales fueron extraídos de su distribución natural (en los estados del Golfo de México y la Península de Yucatán), para establecer los núcleos reproductivos en las UMAs de Puerto Vallarta, Jalisco y Villa Juárez, Sinaloa; sin embargo, se desconocen las localidades de origen. Los nombres de las poblaciones de C. moreletii se asignaron acorde a su origen de nacencia o área de procedencia.
Material biológico: Del total poblacional manejado en la UMA Cacahuatal (N=800), se eligieron y capturaron a diez ejemplares adultos de cada población (n=40; 5%). A cada uno se le extrajeron aproximadamente 1.5mL de sangre del seno post-craneal (justo por detrás de la cabeza), con un equipo Vacutainer® con EDTA como anticoagulante, siguiendo las indicaciones para el tratamiento de las muestras hemáticas descrito en McVay et al. (2008).
Aislamiento y purificación de ADN y amplificación de microsatélites: Antes de aislar el ADN de las muestras sanguíneas, todas se digirieron con proteinasa K a una concentración final de 0.5mg/mL. El ADN de 18 muestras se aisló y purificó con DNeasy Tissue Kit (Qiagen) siguiendo las instrucciones del fabricante. De las restantes 22 muestras, el ADN se aisló a partir de 0.6mL de buffer de extracción CTAB 2% acorde con el protocolo descrito por González & Vovides (2002), y se purificó con columnas de Sephadex.
El ADN de los ejemplares fue caracterizado genotípicamente con siete loci de microsatélites (Cj16, Cj18, Cj20, Cj35, Cj119, Cj128 y Cj131) desarrollados para el género Crocodylus (FitzSimmons et al. 2000), mediante reacción en cadena de la polimerasa (PCR). La amplificación se llevó a cabo en un volumen final de 20μL con las siguientes condiciones: 10mM de buffer Tris-HCl, 50mM KCl, 1.5mM MgCl2 y 0.1% Triton X-100, 1.0mM de cada oligonucleótido, 250μM de cada dNTP, 10-15ng de ADN y 2U de polimerasa Taq (Apex Gene choice). El termociclador (Labnet International Inc.) se programó según los parámetros propuestos para estos loci por Dever & Densmore (2001).
Electroforesis en gel de acrilamida, visualización y registro de alelos: Los productos de PCR se corrieron en geles de acrilamida al 6%, preparados y teñidos siguiendo el procedimiento descrito por Benbouza et al. (2006). La electroforesis se llevó a cabo por 3h a 1 680V, 60mA y 80W en una cámara vertical para electroforesis (Thermo Scientific Owl P10DS Dual Gel System) con buffer TBE 1X (10mM Trizma, 8.9mM ácido bórico y 2mM Na2 EDTA). Previo a la electroforesis, el ADN se desnaturalizó 2min a 92ºC. Entre cada diez muestras, se colocó un marcador de peso molecular (ladder) de 100pb (Promega). Después de la electroforesis los geles se tiñeron con nitrato de plata. La imagen del gel se capturó con una cámara Kodak Digital Science®. Para conocer el tamaño de los alelos (en pares de bases; pb), se utilizó el software 1D Image Analysis Software (versión 3.0), tomando como referencia el tamaño de los fragmentos del ladder.
En Excel® se determinó el tamaño mínimo y máximo de los alelos en pb para cada locus. Con el complemento MICROSATELLITE TOOLKIT para Excel® se calculó el contenido de información polimórfica (PIC) por loci. Se usó el software POPGENE (v1.32, Yeh et al. 1999), para calcular el número de alelos observados (na) y número efectivo de alelos (ne) (Kimura & Crow 1964), heterocigosidad observada (HO) y esperada (HE) y los estadísticos F (FIS y FST) (Nei 1987). Las frecuencias alélicas se obtuvieron con el software FSTAT (v2.9.3, Goudet 2001) y con MICROCHECKER (v2.2.3, Oosterhout et al. 2004) se calcularon las frecuencias de los alelos nulos. Con el software ARLEQUIN (v3.0, Excoffier et al. 2005), se determinó el número de loci polimórficos, porcentaje de polimorfismo y la diversidad alélica total por población (Nei 1987). Con este mismo software se aplicó la prueba exacta de Fisher para el desequilibrio de ligamiento entre todos los posibles pares de loci (Slatkin & Excoffier 1996), y para confirmar la presencia de alelos nulos, se aplicó la prueba de equilibrio de Hardy-Weinberg. Con este programa también se evaluó la estructura genética de las poblaciones aplicando un análisis de varianza molecular (AMOVA) locus por locus (basado en 1 023 permutaciones aleatorias), el cual analiza las frecuencias alélicas por locus, infiriendo jerárquicamente la partición de la variabilidad genética a nivel intra e inter poblacional; además determina los locus que detectan la mayor proporción de variabilidad genética. El AMOVA se aplicó contrastando población por población, y se obtuvo el FST con sus respectivas pruebas de significancia (α<0.05), con los cuales se infirió la estructuración genética de las poblaciones (Excoffier et al. 1992). Los componentes de variabilidad genética de las poblaciones bajo estudio se determinaron asumiendo equilibro Hardy-Weinberg.
Resultados
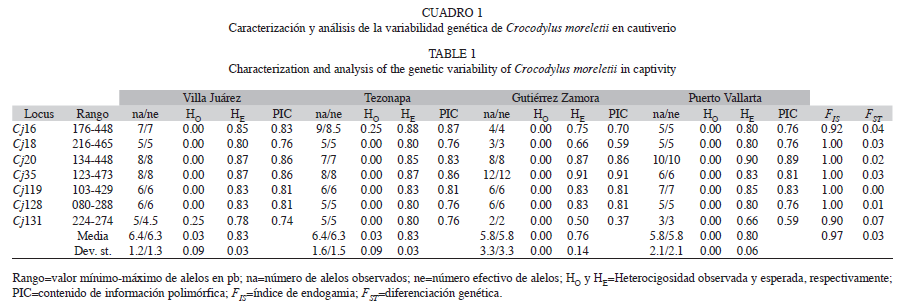
El porcentaje de polimorfismo de los siete loci de microsatélites fue del 100%. Los valores del PIC fueron superiores a 0.5, lo que indica que son altamente variables, con excepción del locus Cj131 en la población de Gutiérrez Zamora que obtuvo un valor de 0.37 (Cuadro 1). El tamaño mínimo y máximo en pb de alelos se presentó en los loci Cj128 (80pb) y Cj35 (473pb), respectivamente. La población de Gutiérrez Zamora fue la que presentó el menor (2 alelos en el locus Cj131) y mayor número de alelos (12 alelos en el locus Cj35) (Cuadro 1). Las poblaciones de Villa Juárez, Sin., y Tezonapa, Ver., presentaron el mismo valor promedio de alelos (6.4), que resultó superior a las otras dos poblaciones que tuvieron un promedio de 5.8 alelos (Cuadro 1). En cada grupo poblacional, se obtuvieron valores similares en el número observado de alelos y número efectivo de alelos (Cuadro 1), lo cual indica un posible efecto fundador. Respecto al análisis de la diversidad alélica total, las poblaciones de Villa Juárez, Sin., y Tezonapa, Ver., presentaron un total de 84 y 82 alelos, respectivamente, mientras que las poblaciones de Gutiérrez Zamora, Ver., y Puerto Vallarta, Jal., presentaron una diversidad alélica total de 78 alelos cada una. La población de Puerto Vallarta, Jal., presentó las frecuencias alélicas más bajas (locus Cj35) y las más altas (locus Cj18) (Fig. 1). Todos los loci presentaron alelos nulos, los cuales tuvieron una frecuencia alta (Fig. 1).
La heterocigosis observada (HO) fue muy baja con respecto a la esperada (HE), indicando un déficit de heterocigotos en todas las poblaciones (Cuadro 1). Sólo se detectaron heterocigotos en las poblaciones de Villa Juárez, Sin., y Tezonapa, Ver., en los loci Cj131 y Cj16 (Cuadro 1). La prueba de desequilibrio de ligamiento genético resultó no significativa para ninguno de los loci (valores de Chi-cuadrado, p=1.000 en la mayoría de los casos). Esta prueba se realizó para determinar si la frecuencia de un locus puede influir en el otro. En este sentido, de las 84 comparaciones para todos los pares de loci (21 comparaciones por pares en cada población), indicaron que la frecuencia entre loci no se encuentran ligadas. Según la prueba de equilibrio de Hardy-Weinberg, todos los valores de HO fueron significativamente menores (p<0.05) respecto a los valores de la HE para todos los loci, lo que confirma la presencia de alelos nulos, resultado que conlleva a obtener altos niveles de endogamia en cada locus, con un valor promedio del FIS de 0.9, contribuyendo simultáneamente a una reducida diferenciación interpoblacional (FST=0.03 promedio) (Cuadro 1).
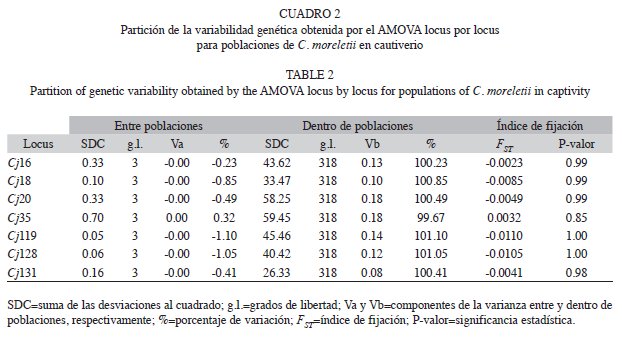
Los resultados del AMOVA locus por locus indicaron que la mayor variabilidad genética ocurre a nivel intrapoblacional (valores iguales al 100% en la mayoría de los loci). Es decir, la fracción de la variabilidad genética es explicada dentro de cada población y sólo el locus Cj35 detectó que existe un 0.32% de variabilidad genética entre poblaciones (Cuadro 2). Un dato interesante que arrojó el AMOVA es la obtención de valores negativos para los componentes de varianza y el porcentaje de variabilidad genética entre las poblaciones (exceptuando el locus Cj35), lo que indica la ausencia de una estructuración genética entre las poblaciones (Cuadro 2). Los valores del FST que se obtuvieron a partir del AMOVA fueron negativos y no significativos (p>0.05), lo que confirma la limitada diferenciación genética entre las subpoblaciones (Cuadro 2).
Discusión
Se encontró que las estimaciones del tamaño mínimo-máximo de alelos así como el número de alelos por locus fueron superiores a lo reportado por Dever & Densmore (2001), Dever et al. (2002), y McVay et al. (2008), para poblaciones de la misma especie en vida silvestre distribuidas en Belice. Estas discrepancias pueden ser debidas al alto polimorfismo que proveen las regiones de microsatélites, particularmente en vertebrados, ya que son regiones conservadas con altas tasas de mutación (Moore et al. 1991). En este contexto, las evaluaciones de la variabilidad genética en Crocodylus (Fitz- Simmons et al. 2000), han demostrado que el polimorfismo se debe a inserciones/deleciones de nucleótidos en las regiones microsatélites (Tauz 1989, Strassmann et al. 1996), generando altas tasas de mutación y alelos múltiples (Queller et al. 1993). La diversidad y las frecuencias alélicas fueron similares a lo reportado para C. rhombifer y C. acutus, posiblemente por la presencia de alelos nulos (Weaver et al. 2008).
En todos los loci, el valor de la HO fue menor a la HE, lo que indica un marcado déficit de heterocigocidad; de hecho, la mayoría de los valores de la HO son iguales a cero, lo que pone en manifiesto la inexistencia de heterocigosidad en todas las poblaciones. Esta baja variabilidad genética puede deberse a la presencia de alelos nulos, los cuales se presentan con cierta frecuencia en microsatélites y se ha sugerido que éstos pudieran mermar los valores de heterocigocidad (Brookfield 1996), por tanto los individuos son catalogados como homocigotos (Aranguren-Méndez et al. 2005). Por un lado, esto pudo surgir basándose en que los individuos de las poblaciones de Puerto Vallarta, Jal., y Villa Juárez, Sin., (que nacieron ex situ y son generación F1), pueden descender de un solo macho dominante, pero también es probable que durante los eventos reproductivos ocurridos en cautiverio en las UMAs de procedencia, el cruzamiento se haya dado entre cocodrilos emparentados; esto también sucede en granjas de C. niloticus, donde los apareamientos entre los individuos reproductivos no es al azar, contribuyendo a disminuir sus niveles de variabilidad genética (Jurgens et al. 1994). En el caso de las poblaciones de procedencia silvestre (Gutiérrez Zamora y Tezonapa, Ver.), que tampoco presentaron variabilidad genética, puede ser reflejo del estado natural de la variabilidad genética de las poblaciones donde proceden. Esto puede atribuirse a la inestabilidad en la estructura demográfica que presentaron las poblaciones de C. moreletii distribuidas en México (Casas 1995), lo que pudo haber conducido a mantener un limitado tamaño efectivo de población (efecto cuello de botella) o un pequeño grupo de individuos reproductivos (efecto fundador) (Bishop et al. 2009). Esto último se confirma cuando se contrastan los valores del número de alelos observados con el número de alelos efectivos, ya que esto pone de manifiesto que estas poblaciones presentan una reducción de variabilidad genética por un posible efecto fundador; sin embargo, se requieren hacer estudios con ejemplares de varias generaciones para corroborar esta hipótesis.
Al comparar los valores de heterocigosidad observada (promedio HO=0.02) para las cuatro poblaciones bajo estudio encontramos que son inferiores a los reportados para otras poblaciones de C. moreletii distribuidas en Belice estudiadas por Dever & Densmore (2001) y Dever et al. (2002) (promedio HO=0.49 en ambos estudios), lo que pone en manifiesto una reducción en los niveles de variabilidad genética, atribuidos principalmente por depresión endogámica en las poblaciones extraídas del medio silvestre. A nivel interespecífico, nuestros resultados son similares a lo reportado para C. niloticus en cautiverio (promedio HO=0.00) a partir de un estudio de proteínas que muestra la ausencia de variabilidad genética debido al efecto fundador ocurrido en cautiverio (Flint et al. 2000). Es preocupante que las poblaciones de procedencia silvestre también presenten una limitada variabilidad genética. En cocodrilos, los niveles de variabilidad genética están determinados por su entorno (Gartside et al. 1977), como respuesta a los factores ambientales (Adams et al. 1980). En este sentido, las concurrentes presiones al hábitat y los consecuentes impactos sobre la dinámica poblacional de C. moreletii distribuidas en el territorio mexicano, puede ser una de las causas de la disminución de su variabilidad genética. C. moreletii es una especie sujeta a protección especial en México y amenazada a nivel internacional, por lo que se debe generar un control más estricto para conservar su variabilidad genética. La suma de esfuerzos en la conservación de sus hábitats donde la prioridad sea la conservación de los corredores biológicos para fomentar la migración entre las poblaciones de estos cocodrilos, puede ser una de las acciones para preservar, y en su caso incrementar la variabilidad genética de esta especie emblemática de los ecosistemas tropicales de México.
Tanto los eventos reproductivos ocurridos en cautiverio como los cambios demográficos por el que transcurrieron las poblaciones en vida silvestre reflejan el déficit de heterocigotos, lo cual quiere decir que las poblaciones bajo estudio son endogámicas, consistente con lo determinado por el FIS, que está influenciado por tener alelos comunes entre los individuos, que generalmente implica que son idénticos por descendencia (Raybould et al. 2002). La endogamia no siempre representa una desventaja para las especies de cocodrilianos, sino que puede considerarse como una adaptación óptima al responder a las presiones del medio (Gartside et al. 1977). Sin embargo, se considera que la endogamia y la limitada variabilidad genética está jugando un papel determinante al relacionar estos parámetros genéticos con las diferenciaciones en ciertos caracteres morfológicos y las bajas tasas de crecimiento corporal que se han observado en estos grupos poblacionales (Serna-Lagunes et al. 2010). Esto representa una problemática en la UMA Cacahuatal ya que impacta de manera negativa en los ingresos económicos debido a la obtención de pieles de menor calidad y consecuentemente mantener a los individuos por un mayor tiempo hasta alcanzar la talla comercial (Serna-Lagunes 2010).
El AMOVA indicó que la mayor proporción de la variabilidad genética se presentó dentro de cada población. Por tanto, gran parte de la variación genética fue aportada por la divergencia de los alelos dentro de cada subpoblación y no se detectó una estructuración genética entre las poblaciones. El presentar alelos idénticos se debe a que las poblaciones de C. moreletii fueron extraídas del rango de distribución natural, donde históricamente tenían una distribución más o menos continúa (Álvarez del Toro & Sigler 2001), y posiblemente hubo flujo genético, y los alelos desde entonces, así se han mantenido, y tienden a obtener una baja diferenciación entre poblaciones formando un grupo homogéneo (Weir 1996).
Una de las implicaciones inmediatas para el manejo genético de C. moreletii es continuar realizando estudios de caracterización genética de poblaciones silvestres y de las que se encuentran en las diferentes UMAs, con la finalidad de identificar individuos genéticamente diferentes y con mayor variabilidad genética para seleccionarlos y generar un nuevo núcleo reproductivo en la UMA Cacahuatal; estos cocodrilos pueden ser transportados para que puedan copular con las hembras (también caracterizadas genéticamente) de otras UMAs. Inclusive también se puede pensar en inseminación artificial, aunque esta técnica reproductiva aún se encuentra en proceso de desarrolloimplementación- validación para cocodrilos. Dado que los problemas genéticos derivados de endogamia minimizan los procesos básicos de adaptación local de las especies, por ahora no puede recomendarse un programa de liberación del núcleo de individuos de la UMA Cacahuatal a su hábitat silvestre. La liberación solo se podrá realizar hasta que se tenga una proporción similar de variabilidad genética como la que se ha reportado para otras poblaciones como las de Belice (HO=0.49), o tomar como referencia los niveles de variabilidad genética que se reporten en estudios posteriores con poblaciones de C. moreletii distribuidas en México pero que se encuentren en un rango de HO entre 0.3 a 0.5. Como en la mayoría de las UMAs se obtiene píe de cría extraídas de poblaciones silvestres, se puede sugerir que previo a la formación de sus núcleos reproductivos se realicen estudios siguiendo los métodos utilizados en este trabajo y solo considerar aquellos individuos genéticamente diferentes para la reproducción, lo que podría llevar a la disminución de la homocigosis en sus descendientes. Estudios futuros deben proporcionar información sobre el tamaño efectivo de población y el tamaño mínimo viable que se requiere para conservar la variabilidad genética de poblaciones de C. moreletii en cautiverio con la finalidad de prevenir el detrimento genético por endogamia. Esto podría preservar la variabilidad genética de los cocodrilos durante ciertos periodos de tiempo generacional y así tener bases más sólidas para generar un plan de liberación de cocodrilos a su hábitat para garantizar su adaptación y sobrevivencia.
Se puede concluir que la limitada variabilidad genética de las poblaciones de C. moreletii que nacieron en las UMAs (Villa Juárez, Sin. y Puerto Vallarta, Jal.), se debió a un inadecuado manejo reproductivo en estos sistemas de crianza, permitiendo el apareamiento de un limitado grupo de reproductores o de un solo macho dominante. En el caso de las poblaciones extraídas de su hábitat silvestre (Tezonapa y Gutiérrez Zamora, Veracruz), su limitada variabilidad genética se presentó por el reducido tamaño efectivo de la población, inferido principalmente por la variación en la dinámica demográfica que, históricamente han tenido las poblaciones. A nivel inter e intraespecífico, los parámetros de variabilidad genética fueron menores a los reportados para la misma especie y para otros cocodrilos, por lo que se deben considerar medidas específicas que conserven y/o incrementen la variabilidad genética en las poblaciones bajo estudio. La mayor proporción de la variabilidad genética se presentó a nivel intrapoblacional y no presentaron diferenciación ni subdivisión genética, posiblemente por el alto flujo genético que históricamente presentó C. moreletii en su distribución natural y la fijación de alelos se mantuvo desde entonces. Finalmente, el número de loci analizados reflejó el estado actual de la variabilidad genética del grupo poblacional en estudio; por tanto se puede ampliar el análisis aumentando el número de individuos caracterizados junto con la inclusión de otras poblaciones, a fin de llegar a conclusiones más precisas para generar un plan de recuperación de las poblaciones de C. moreletii en las localidades donde fueron extintas.
Agradecimientos
A la Secretaria de Medio Ambiente y Recursos Naturales (SEMARNAT) por brindar el permiso especial de colecta SGPA/ DGVS/07875. A la UMA Cacahuatal por los apoyos brindados para realizar el presente estudio. A la Línea Prioritaria de Investigación 11 y al Campus Veracruz, filiales del Colegio de Postgraduados, así como al Laboratorio de Sistemática Molecular del Instituto de Ecología, A. C. por sus respectivos financiamientos.
Referencias
Adams, S.E., M.H. Smith & R. Baccus. 1980. Biochemical variation in the American alligator (Alligator mississipiensis). Herpetologica 36: 289-296. [ Links ]
Álvarez del Toro, M. & L. Sigler. 2001. Los Crocodylia de México. Instituto Mexicano de Recursos Naturales y Procuraduría Federal de Protección al Ambiente. Distrito Federal, México. [ Links ]
Amavet, P.S., L.E. Rosso, M.R. Markariani & A. Larriera. 2007. Analysis of the population structure of Broad- Snouted Caiman (Caiman latirostris) in Santa Fe, Argentina, using the RAPD technique. J. Herpetol. 41: 294-300. [ Links ]
Arangueren-Méndez, J.A., R. Román-Bravo, W. Isea, Y. Villasmil & J. Jordana. 2005. Los microsatélites (STR’s), marcadores moleculares de ADN por excelencia para programas de conservación: una revisión. Arch. Latinoam. Prod. Anim. 13: 1-6. [ Links ]
Benbouza, H., J. Jean-Marie, B. Jean-Pierre & M. Guy. 2006. Optimization of a reliable, fast, cheap and sensitive silver staining method to detect SSR markers in polyacrylamide gels. Biotechnol., Agron., Soc. Environ. 10: 77-81. [ Links ]
Bishop, M.J., A.J. Leslie, S.L. Bourquin & C.O’Ryan. 2009. Reduced effective population size in an overexploited population of the Nile crocodile (Crocodylus niloticus). Biol. Conserv. 142: 2335-2341. [ Links ]
Brookfield, J.F.Y. 1996. A simple new method for estimating null allele frequency from heterozygote deficiency. Mol. Ecol. 5: 453-455. [ Links ]
Casas, A.G. 1995. Los cocodrilos de México como recurso natural. Presente, pasado y futuro. Rev. Soc. Mex. Hist. Nat. 46: 153-162. [ Links ]
Casas, A.G. 2002. Hacia la conservación y manejo sustentable del lagarto o cocodrilo de pantano (Crocodylus moreletii) en México, p. 27-45. In L.M. Verdade & A. Larriera (eds.). La conservación y el manejo de caimanes y cocodrilos de América Latina. CN Editoria, Piracicaba, Sao Paulo, Brasil. [ Links ]
Convención Internacional del Comercio y Tráfico de Especies Amenazadas de Flora y Fauna (CITES). 2008. Propuesta para transferir la población mexicana Crocodylus moreletii del Apéndice I al Apéndice II. Acuerdo 23, Documento 18, Ginebra, Suiza. [ Links ]
Davis, L.M., T.C. Glenn, R.M. Elsey, I.L. Brisbin Jr., W.E. Rhodes, H.C. Dessauer & R.H. Sawyer. 2000. Genetic structure of six populations of American alligators: a microsatellite analysis, p. 38-50. In G.C. Grigg, F. Seebacher & C.E. Franklin (eds.). Crocodilian biology and evolution. Surrey Beatty and Sons, Chipping Norton, Australia. [ Links ]
Dever, J.A. & D. Densmore. 2001. Microsatellites in Morelet’s Crocodile (Crocodylus moreletii) and their utility in addressing crocodilian population genetics questions. J. Herpetol. 35: 541-544. [ Links ]
Dever, J.A., R.E. Stratus, T.R. Rainwater, S.T. McMurry & L.D. Densmore. 2002. Genetic diversity, population subdivision and gene flow in Morelet’s Crocodile (Crocodylus moreletii) from Belize, Central America. Copeia 4: 1078-1091. [ Links ]
Diario Oficial de la Federación (DOF). 2001. Norma Oficial Mexicana NOM-059-ECOL-2001, Protección Ambiental-especies nativas de México de Flora y Fauna silvestre, tres categorías de riesgo y especificaciones para su inclusión, exclusión o cambio- Lista de Especies en riesgo. Diario Oficial de la Federación, Distrito Federal, México. [ Links ]
Domínguez-Laso, J. 2006. Determinación del estado actual de las poblaciones silvestres del cocodrilo de pantano (Crocodylus moreletii) en México y evaluación de su estatus en la CITES. Informe final SNIB-CONABIO proyecto No. CS009. Instituto de Historia Natural y Ecología, Distrito Federal, México. [ Links ]
Excoffier, L., G. Laval & S. Schneider. 2005. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evol. Bioinf. Online 1: 47-50. [ Links ]
Excoffier, L., P. Smouse & J. Quattro. 1992. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131: 479-491. [ Links ]
FitzSimmons, N., J.C. Buchan, P.V. Lam, G. Polet, T.T. Hung, N.Q. Thang & J. Gratten. 2002. Identification of purebred Crocodylus siamensis for reintroduction in Vietnam. J. Exp. Zool. 294: 373-381. [ Links ]
FitzSimmons, N., S. Tanksley, M.R. Forstner, E.E. Louis, R. Daglish, J. Gratten & S. Davis. 2000. Microsatellite markers for Crocodylus: new genetic tools for population genetics, mating system studies and forensics, p. 51-57. In G.C. Grigg, F. Seebacher & C.E. Franklin (eds.). Crocodilian biology and evolution. [ Links ]
Surrey Beatty & Sons, Chipping Norton, Australia. Forstner, M. & J.M. Forstner. 2002. Aplicaciones del DNA en la conservación de los cocodrilianos, p. 99-117. In L.M. Verdade & A. Larriera (eds.). La conservación y el manejo de caimanes y cocodrilos de América Latina. CN Editoria, Piracicaba, Sao Paulo, Brasil. [ Links ]
Flint, N.S., F.H. Van Der Bank & J.P. Grobler. 2000. A lack of genetic variation in commercially bred Nile crocodiles (Crocodylus niloticus) in the north-west province of South Africa. Water SA 26: 105-110. [ Links ]
Gartside, D.F., H.C. Dessauer & T. Joanen. 1977. Genic homozygosity in an ancient reptile Alligator mississippiensis. Biochem. Genet. 15: 655-664. [ Links ]
González, D. & A.P. Vovides. 2002. Low intralineage divergence in Ceratozamia (Zamiaceae) detected with nuclear ribosomal DNA ITS and chloroplast DNA trnL-F non-coding region. Syst. Bot. 27: 654-661. [ Links ]
Goudet, J. 2001. FSTAT: a program to estimate and test gene diversities and fixation indices. Institute of Ecology, Biology Building, Université of Lausanne, Suiza. [ Links ]
Glenn, T.C. 2002. DNA tools and resources for crocodilian research, p. 211. In Crocodiles, Proceedings of the 16th Working Meeting of the Crocodile Specialist Group of the Species Survival Commission of IUCN and the World Conservation Union (eds.). Gland, Suiza. [ Links ]
Huchzermeyer, F.W. 2003. Crocodiles: Biology, husbandry and diseases. CABI, Wallingford, Oxfordshire, Reino Unido. [ Links ]
INE. 2000. Proyecto para la conservación, manejo y aprovechamiento sustentable de los Crocodylia de México (COMACROM). Instituto Nacional de Ecología y Secretaria de Marina y Recursos Naturales, Distrito Federal, México. [ Links ]
IUCN, 2010. Red List of Threatened Species, Version 2010.2. International Union for Conservation of Nature and Natural Resources (Consultado: 17 Agosto 2010, http://www.iucnredlist.org). [ Links ]
Jurgens, A., J. Kruger & F.H. Van Der Bank. 1994. Allozyme variation in the Nile crocodile (Crocodylus niloticus) form Southern Africa, p. 72-76. In Crocodiles, Proceedings of the 12th Working Meeting of the Crocodile Specialist Group IUCN and the World Conservation Union (eds.). Gland, Suiza. [ Links ]
Kimura, M. & J.F. Crow. 1964. The number of alleles that can be maintained in a finite population. Genetics 49: 725-38. [ Links ]
Luxmoore, R.A. 1992. Directory of Crocodilian farming operations. IUCN, Gland, Suiza and Cambrigde, Reino Unido. [ Links ]
Moore, S.S., L.L. Sargeant, T.J. King, J.S. Mattick, M. Georges & J.S. Hetzel. 1991. The conservation of dinucleotide microsatellites among mammalian genomes allows the use of heterologous PCR primer pairs in closely related species. Genomics 10: 654-660. [ Links ]
McVay, J.D., D. Rodriguez, T.R. Rainwater, J.A. Dever, S.G. Platt, S.T. McMurry, M.R.J. Forstner & L.D. Densmore. 2008. Evidence of multiple paternity in Morelet’s crocodile (Crocodylus moreletii) in Belize, CA, inferred from microsatellite markers. J. Exp. Zool., Part A. 10: 643-648. [ Links ]
Nei, M. 1987. Molecular evolutionary genetics. Columbia University Press, Nueva York, EEUU. [ Links ]
Oosterhout, C., W.F. Hutchinson, D.P.M. Wills & P. Shipley. 2004. MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004: 535-538. [ Links ]
Porras, M.L.P., J.R.M. Bolaños & B.R. Brarr. 2008. Variación genética y flujo de genes entre poblaciones de Crocodylus acutus (Crocodylia: Crocodylidae) en tres ríos del pacífico central, Costa Rica. Rev. Biol. Trop. 56: 1471-1480. [ Links ]
Queller, D.C., J.C. Strassmann & C.R. Hughes. 1993. Microsatellies and kinship. Trends Ecol. Evol. 8: 285-288. [ Links ]
Raybould, A.F., R.T. Clarke, J.M. Bond, R.E. Welters & C.J. Gliddon. 2002. Inferring patterns of dispersal from allele frequency data, p. 89-108. In J.M. Bullok, R. Kenward & R. Haills (eds.). Dispersal ecology. Blackwell Science, Oxford, Reino Unido. [ Links ]
Ross, J.P. 1998. Crocodiles: Status Survey and Conservation Action Plan. Crocodile Specialist Group, the World Conservation Union, Gland, Suiza. [ Links ]
Serna-Lagunes, R. 2010. Historia de vida y genética cuantitativa de cuatro poblaciones del cocodrilo de pantano (Crocodylus moreletii) bajo condiciones de cautiverio en el trópico mexicano. Tesis de Maestría, Colegio de Postgraduados, Estado de México, México. [ Links ]
Serna-Lagunes, R., J.J. Zúñiga-Vega, P. Díaz-Rivera, F. Clemente-Sánchez, A. Pérez-Vázquez & J.L. Reta- Mendiola. 2010. Variabilidad morfológica y crecimiento corporal de cuatro poblaciones de Crocodylusmoreletii en cautiverio. Rev. Mex. Biodivers. 81: 713-719. [ Links ]
Slatkin, M. & L. Excoffier. 1996. Testing for linkage disequilibrium in genotypic data using the Expectation- Maximization algorithm. Heredity 76: 377-383. [ Links ]
Strassmann, J.E., C.R. Solis, J.M. Peters & D.C. Queller. 1996. Strategies for finding and using highly polymorphic DNA microsatellite loci for studies of genetic relatedness and pedigrees, p. 163-180. In J.D. Ferraris & S.R. Palumbi (eds.). Molecular Zoology: Advances, Strategies and Protocols. Wiley-Liss Inc., Nueva York, EEUU. [ Links ]
Tautz, D., M. Trick & D.A. Dover. 1986. Cryptic simplicity in DNA is a major source of genetic variation. Nature 322: 652-656. [ Links ]
Tautz, D. 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res. 16: 6463-6471. [ Links ]
Wang, Y.Q., W.Q. Zhu, L. Huang, K.Y. Zhou & R.P. Wang. 2006. Genetic diversity of Chinese alligator (Alligator sinensis) revealed by AFLP analysis: An implication on the management of captive conservation. Biodivers. Conserv. 15: 2945-2955. [ Links ]
Weaver, J.P., D. Rodríguez, M. Venegas-Anaya, J.R. Cedeño-Vázquez, M.R.J. Forstner & L.D. Densmore III. 2008. Genetic characterization of captive Cuban crocodiles (Crocodylus rhombifer) and evidence of hybridization with the American crocodile (Crocodylus acutus). J. Exp. Zool., Part A. 10: 649-660. [ Links ]
Weir, B.S. 1996. Genetic data analysis II. Methods for discrete population genetic data. Sinauer Associates, Inc. Publishers, Sunderland, Massachusetts, EEUU. [ Links ] Wu, X., Y. Wang, K. Zhou, W. Zhu, J. Nie, C. Wang & W. Xie. 2002. Genetic variation in captive population of Chinese alligator, Alligator sinensis, revealed by random amplified polymorphic DNA (RAPD). Biol. Conserv. 106: 435-441. [ Links ]
Yang, P., X. Wu, Y. Wang, Z. Jiang, C. Gu & C. Wang. 2006. AFLP Analysis of genetic variation on captivebred Chinese alligators: an application to select individuals for release. Zoo Biol. 6: 479-490. [ Links ]
Yeh, F.C., R.C. Yang & T. Boyle. 1999. POPGENE version 1.32: Microsoft Windows -based freeware for population genetic analysis, quick user guide. University of Alberta, Center for International Forestry Research, Canada. [ Links ]
Zucoloto, R.B., L.M. Verdade & L.L. Coutinho. 2002. Microsatellite DNA library for Caiman latirostris. J. Exp. Zool., Part B. 294: 346-351. [ Links ]
Zucoloto, R.B., P.M.V. Schimidt, L.M. Verdade & L.L. Coutinho. 2006. Cross-species microsatellite amplification in South American Caimans (Caiman spp. and Paleosuchus palpebrosus). Genet. Mol. Biol. 29: 75-78. [ Links ]
*Correspondencia:
Ricardo Serna-Lagunes: Campus Veracruz, Colegio de Postgraduados, km. 88.5 Carretera Federal Xalapa-Veracruz, Tepetates, Municipio de Manlio F. Altamirano, C.P. 91700, Apartado Postal 421, Veracruz, México; rserna@colpos.mx
Dolores González: Laboratorio de Sistemática Molecular. Instituto de Ecología, A.C., Carretera antigua a Coatepec N.º 351, El Haya, Xalapa, C.P. 91070, Apartado Postal 63, Veracruz, México; dolores.gonzalez@inecol.edu.mx
Pablo Díaz-Rivera: Campus Veracruz, Colegio de Postgraduados, km. 88.5 Carretera Federal Xalapa-Veracruz, Tepetates, Municipio de Manlio F. Altamirano, C.P. 91700, Apartado Postal 421, Veracruz, México; pablod@colpos.mx
1. Campus Veracruz, Colegio de Postgraduados, km. 88.5 Carretera Federal Xalapa-Veracruz, Tepetates, Municipio de Manlio F. Altamirano, C.P. 91700, Apartado Postal 421, Veracruz, México; rserna@colpos.mx, pablod@colpos.mx
2. Laboratorio de Sistemática Molecular. Instituto de Ecología, A.C., Carretera antigua a Coatepec N.º 351, El Haya, Xalapa, C.P. 91070, Apartado Postal 63, Veracruz, México; dolores.gonzalez@inecol.edu.mx
Recibido 02-III-2011. Corregido 20-IV-2011. Aceptado 30-V-2011.


{kind=link}
{kind=link}
{kind=link}