










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Embryonic and larval development of Eugerres mexicanus (Perciformes: Gerreidae) in Tenosique, Tabasco, Mexico
Raúl E. Hernández1*, Martha A. Perera1, Alfonso Castillo1 , Emiliano Luna H1,José A. de la Cruz1, Luis M. Gómez2* & José Valdez Zenil3*
*Dirección para correspondencia
Abstract
Most studies on Eugerres mexicanus mainly consider biogeographic and systematic aspects and rarely address reproductive characteristics, which are useful for fishery population management plans. This study aimed at evaluating the ontogeny of E. mexicanus, based on 30 embryos and 30 larvae sampled by induced spawning of breeders, taken in February 2009 from the Usumacinta River in Tenosique, Tabasco, Mexico. All descriptions of the embryonic development were based on morphometric and meristic data and followed standard methods. Eggs, recovered at the gastrula stage, had an average diameter of 1.17mm (SD=0.08). The bud stage appeared during the first three hours of development, in which the posterior side was adhered to the vitellus; Kupffer´s vesicle was visible. Yolk-sac larvae hatched 18 hours after fertilization, exhibiting a light brown color and an average total length of 2.94mm (SD=0.70); the preflexion stage was reached eight days after hatching, with a total average length of 4.67mm (SD=0.50) and a total notochord length of 4.45mm (SD=0.50). The flexion stage was reached on the 16th day, with an average total length of 6.66mm (SD=1.53), while postflexion was reached on the 24th day, with 10.33mm (SD=1.45). The pre-juvenile stage was reached on the 33rd day, with a total length of 14.30mm (SD=0.93), showing IX spines and 10 rays and III spines and eight rays in the dorsal and anal fins, respectively. The juvenile stage was reached by the 45th day, with an average length of 28.16mm (SD=1.93) and average weight of 4.75g (SD=1.49). Prejuveniles showed an initial pigmentation with dark colored dots in the superior and inferior jaw and dispersed on the head, while juveniles presented the same pigmentation pattern, decreasing towards the margin of the caudal peduncle. In conclusion, the embryonic developmental stages of E. mexicanus were typical for the Gerreidae group. However, their morphometric characters were slightly different since the diameter and size of the drop of oil were bigger than those reported for marine species. In addition, regarding pigmentation, the yolk-sac larvae of E. mexicanus were olive and yellow on the margin of the notochord, which differs from those reported for other species. This is the first recorded report on the reproductive biology and early life development of this species.
Key words: reproduction, induction, embryo, larva, Eugerres mexicanus, Tabasco.
Resumen
La ontogenia se basó en 30 embriones y 30 larvas, obtenidos mediante la inducción del desove de reproductores provenientes de la ribera del río Usumacinta en Tenosique, Tabasco, México, recolectados en febrero de 2009. La descripción se fundamentó en el registro morfométrico y merístico. Los huevos fueron recuperados en estado de gástrula y presentaron un diámetro de promedio de 1.17mm (SD=0.08). Durante las primeras tres horas de desarrollo embrionario, se presentó la etapa de capullo, en la que se observó la región caudal adherida al vitelo, apreciándose la vesícula de Kupffer. Las larvas con saco eclosionaron a las 18 horas pos-fertilización, fueron de color marrón claro con un promedio de 2.94mm; (SD=0.70) de longitud total y alcanzaron la preflexión a los ocho días post-eclosión con una longitud total promedio de 4.67mm; (SD=0.50) y una longitud total del notocordio de 4.45; (SD=0.50). A los 16 días de la eclosión alcanzaron la flexión, con un promedio de 6.66mm; (SD=1.53) de longitud total. La postflexión se presentó a los 24 días con 10.33mm; (SD=1.45). Al llegar a los 33 días, se presentó la fase prejuvenil y llegaron a medir 14.30; (SD=0.93) de longitud total, presentando IX espinas y 10 radios en la aleta dorsal y III y ocho en la aleta anal. Los juveniles midieron 28.16; (SD=1.93) de longitud a los 45 días, con un peso promedio de 4.75g; (SD=1.49). Los prejuveniles presentaron una pigmentación inicial en la mandíbula superior e inferior con tintes oscuros en forma de puntos y de manera dispersa sobre la cabeza. En los juveniles se observó el mismo patrón de pigmentación, disminuyendo hacia el margen del pedúnculo caudal. Las características descriptivas de la etapa de desarrollo embrionario de E. mexicanus son típicas del período de desarrollo de los peces de la familia Gerreidae, en particular en el caso de las especies E. brasilianus y E. lineatus que habitan en ambientes marinos. Sin embargo, sus caracteres morfométricos son diferentes con respecto al diámetro y el tamaño de los huevos y de la gota de aceite, ya que en E. mexicanus son más grandes que los de las especies marinas y son similares a los de los peces de agua dulce. Con respecto a la pigmentación, la larva con saco de E. mexicanus presenta un olor olivo y amarillo sobre el margen del notocordio, lo cual difiere a lo reportado para E. lineatus,ya que en esta muestra un grupo de melanóforos entre los miomeros nueve al 13 como principal característica, y para Diapturus peruvianus por presentar tres manchas en el margen dorsal de los intestinos desde la inserción de la aleta pectoral hasta el ano. Los resultados de este estudio son los primeros registrados para esta especie y han generado información sobre aspectos de su biología reproductiva y el desarrollo de la vida temprana.
Palabras clave: reproducción, inducción, embrión, larva, Eugerres mexicanus, Tabasco.
Although Gerreidae are mostly marine fish, they are frequently found in estuaries, particularly near mangroves, and occasionally in continental environments (Castro et al. 1999). At least one species, Eugerres mexicanus, is confined to the freshwater environments of Southeastern Mexico and Northern Guatemala (González 2005, Miller et al. 2005, González & Rodiles 2007, González et al. 2007). Commonly known as White Mojarra, it is distributed throughout the Atlantic watershed of Southern Mexico in the Coatzacoalcos and Usumacinta river basins and in Northern Guatemala (Gilmore et al. 2002). It is a tropical freshwater demersal species found in neotropical ecosystems (Deckert & Greenfield 1987) and caught in the Usumacinta River, especially in the municipality of Tenosique in Tabasco, Mexico, where it is an important food source sold as a fresh or frozen product. Its body is relatively high, with a slightly high convex pre-dorsal profile, ascending softly up to the first dorsal spine. Its dorsal fin has a concave posterior profile. It also has a wide inter-orbital space, a groove or maxillary process without scales, an oval central depression, and big eyes. Its mouth, terminal or sub-terminal, is relatively big and very protractile, with thick lips. Its body is ventrally depressed towards the belly. Substantial morphologic variations have been observed among populations and among the main areas of distribution in the Grijalva and Usumacinta rivers (González 2005).
Studies on E. mexicanus have focused on systematic and biogeographic aspects (González 2005), its meristic and morphometric characters, and its taxonomy (Deckert & Greenfield 1987, Gilmore et al. 2002). However, few studies have been carried out on the reproduction of the species from the genus Eugerres, in contrast with the number of studies on E. lineatus and E. brasilianus, as well as on Diapterus peruvianus (Álvarez et al. 1996, Jiménez et al. 2003, Ortíz et al. 2008). The embryonic and larval development of several species of this genus is unknown. Considering the fishery and ecological importance of E. mexicanus, this study was designed to describe its embryonic and larval development, in order to increase knowledge about the species and provide useful data for future research.
Materials and methods
Study site: This study was carried out in February 2009 at the Recreo locality (17°28’52’’ N and 91°25’17’’ W) on the Usumacinta River. Five females and two males of the species E. mexicanus were collected with a seine net with a mesh size of 1x1cm, 50m long and 3m wide. The specimens were transported to the laboratory located in the División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, Tenosique, Tabasco, Mexico, in containers with 60L of water, aeration and 4mL of Nuetraestres anti-stress per liter of water to decrease the effect of stress.
All specimens were measured, and weighed on a Torrey digital scale (capacity 0.4 to 20 000g). Pasteur pipettes were used for cannulation to confirm the sexual maturity of the specimens. Oocytes in the final stage of vitellogenesis measuring an average of 0.90mm (SD=0.05, n=26) in females and motile sperm in males were observed using a Labomed CXRIII compound microscope. The average number of eggs for females was 27 300.
Spawning induction: Spawning was induced with human chorionic gonadotropin hormone (CGH) Pregnyl using 5 000UI. All five females and two males weighting an average of 133.2g and 85g, respectively, received 50UI/100g/fish. Two ratios were established: female and male at 2:1 and 3:1 to produce spawning. The organisms were placed in 2 000L water containers, where natural nests were simulated with raffia thread. Aeration was provided constantly by an aerator. The water was circulated by a pump with a potency of 1Hp and filtered with a sand filter.
Parameter records: Spawning was carried out maintaining an average temperature of 30.5°C; (SD=1.9), an electric conductivity of 0.29μS (SD=0.40); pH at 8.1 (SD=0.10); dissolved oxygen at 6.8m/L (SD=4.8), total chlorine at 0.15mg/L (SD=0.30); nitrite at 0.70mg/L (SD=0.40) and nitrate at 0.0mg/L. During the incubation stage, parameters were maintained at 28.6°C (3.1); 0.55μS (0.10); 8.2 (0.10); 2.7mg/L (4.8); 0.10 mg/L (0.00), without nitrite and nitrate. At larval stage, parameters were at 27.1°C (1.90): 0.55μS (0.10); 7.9 (1.2); 5.30mg/L (2.1); 0.05mg/L (0.01), without nitrogen compounds.
Measurement of embryos and larvae: A total of 30 eggs with embryos and 30 larvae were extracted every three hours. They were observed with a Motic BA300 binocular microscope, photographed with a Digitan Moticam 2 300 digital camera, and measured using Motic Images Plus software (2.0ML, 3.0 mega pixels). The recording of the embryonic and larval development: preflexion, flexion, postflexion and juveniles, was based on the criteria described in Hardy (1978). Other measurements included the length and width of the mouth, eye, orbital circle, and orbital fossa, and the length of the head, pre-anal and post-anal lengths, and length to the fork. The meristic characters of the fins included width of the dorsal, anal and pectoral fins, as well as the number of spines and rays on the fins during the larval development (Rosas et al. 2008).
Results
The specimens collected in the wild presented viable sexual products. Females presented oocytes in the vitellogenesis stage with an average diameter of 0.90mm (SD=0.05) and males presented spermatic fluid with mobile sperm. The female: male ratio proposed in this study produced a 100% spawning and an egg fertility index of 90-95%. Survival during the larval development was 0.02%.
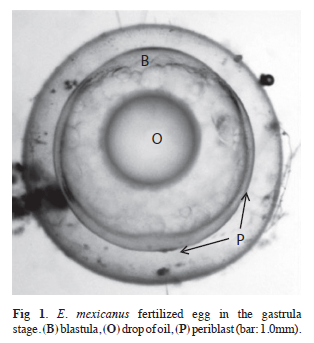
Embryonic development: Fertilized eggs were recovered in the gastrula stage with an average diameter of 1.17mm (SD=0.08). They were adhered to the substrate and presented a big drop of oil (Fig. 1, Table 1 and 3).
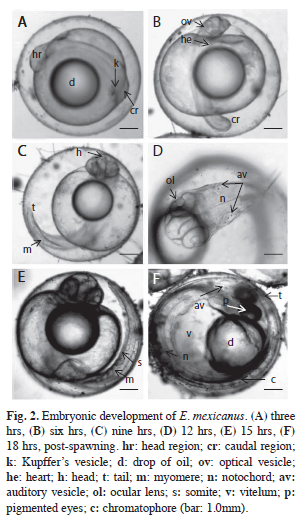
On the first day following fertilization, the early embryo stage was observed three hours post-gastrula, with an average diameter of 1.19 (0.04). The caudal region was adhered to the vitellus, where Kupffer´s vesicle was visible, and the cephalic region had an ovoid projection (Fig. 2A, Table 1). The free tail stage occurred six hours after fertilization, and the developing optical vesicles could be observed in the head and the heart. The heart presented contractions that generated beats (Fig. 2B, Table 1). After nine hours, the embryo increased in size, mainly around the caudal region, with the myomeres or segments of the developed muscles becoming visible (Fig. 2C, Table 1). Twelve hours post-fertilization, the embryo exhibited vibrating movements in the caudal region, where vesicles or ear plaques appeared, and a developed notochord could also be observed (Fig. 2D, Table 1).
Recently hatched larvae: The posthatching yolk-sac larvae were light brown in color, had an average total length of 2.94mm (SD=0.7) and a width of 0.79 (0.10). The notochord had an average length of 2.74mm (0.80). They showed a longitudinally oval vitellum with an average length of 0.98mm (0.20) and a width of 0.59 (0.20). An average big drop of oil could be seen, measuring 0.49mm (0.10) long and 0.49 (0.10) wide (Fig. 3). The larvae on day one, specifically eight hours post-hatching, had an average total length of 4.02mm (0.26) and a width of 0.79 (0.07). The head and the notochord were 0.56mm (0.05) and 3.85mm (0.24) long, respectively. The vitellum had an average length of 0.99mm (0.12) and width of 0.61 (0.09), and was in the process of re-absorption. The drop of oil measured an average of 0.48mm (0.09) long and 0.47 (0.08) wide. On the second day (hour 20) a pair of ear plaques could be seen in the cephalic region, together with dark pigmented spots or melanophores on the body, and the mouth was open. The completely pigmented eyes of the larvae were an average of 0.24mm (0.02) long and 0.22 (0.01) wide. The orbital circle was an average of 0.26mm (0.02) long and 0.31 (0.38) wide, and the average orbital fossa was 0.29mm (0.18) long and 0.28 (0.01) wide. The larvae had an average pre-anal length of 1.44mm (0.23) and a post-anal length of 2.00mm (0.28) (Table 4).
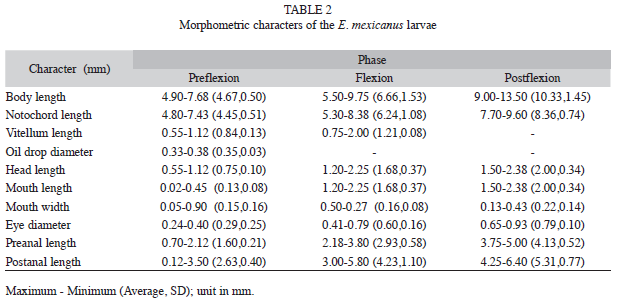
Flexion: This developmental stage started 16 days (372 hours) post-hatching, when flexion of the notochord was observed in the region of the caudal fin of the embryo. A swim bladder could also be seen over the intestinal region and the intestine showed food in the process of decomposition. The primordium of the dorsal fin and a rudimentary anal fin could also be seen, together with small developing rays (Fig. 3, Table 2).
Postflexion: Larvae reached this stage 24 days post-hatching, with an average total length of 10.33mm (SD=1.45) and a total width of 1.73 (0.51) (Table 2 and 4). Aftermetamorphosis, the caudal fin was observed to be forked, with 17 fin rays, each with six segments. The length of the larvae to the fork varied 9.17 (0.44). At hour 578 post-hatching, the larvae presented small canine teeth on the maxilla and mandible in the mouth, measuring an average of 0.35mm (0.09) long and 0.03 (0.01) wide (Fig. 3). The complementary morphometric characters are shown in Table 2. Likewise, these larvae presented totally formed dorsal, anal, pelvic and caudal fins. By hour 692 post-hatching, the dorsal fin measured an average of 2.0mm (0.40) long and 0.62 (0.13) wide and presented IX spines: the first smaller than the others measuring an average of 0.25 (0.22), the second 0.32 (0.16), the third 0.37 (0.20), the fourth 0.31 (0.24), and the fifth 0.18 (0.13), with 10 rays. The anal fin was an average of 1.99mm (0.35) long and 0.47 (0.16) wide and presented III spines, the first small with an average length of 0.55mm (0.35) and the second one with an average length of 0.34mm (0.11) long. Eight rays were also observed measuring an average of 1.08mm (0.60) long. The pectoral and pelvic (n=5) fins were 0.35mm and 0.50mm long, respectively (Fig. 3, Table 5).
Prejuveniles: This phase started 33 days (792 hours) post-hatching, with an average total length of 14.3mm (SD=0.93) and a width of 3.12 (0.21). The mouth-fork length was 12.83mm (0.76). The dorsal fin measured 1.28mm long and 0.08mm wide and presented IX spines and 10 rays. The first spine measured an average of 0.30mm (SD=0.04) long, the second 1.23mm (0.09), the third 1.40mm (0.62), the fourth 1.42mm (0.14), the fifth 1.15mm (0.09) and the sixth 0.88mm (0.12). It was not possible to measure the other spines. Upon reaching hour 845, the anal fin was an average of 0.08 (0.00) wide, with III spines and eight rays. The first spine measured an average of 0.27 (0.02), the second 1.06 (0.08) and the third 1.40 (0.16). The pectoral fins measured 1.32mm (0.26) in length and 0.94 (0.08) in width. The pelvic fins (n=3) were an average of 1.42 (0.08) long and 0.47 (0.20) wide. At the end of this phase, prejuveniles had a length of 18mm to the fork and a total length of 20mm (Fig. 3, Table 5).
Juveniles: This stage occurred 45 days (1 080 hours) post-hatching with an average total length of 28.16mm (SD=1.93, n=5), a width of 30.1 (36.8, 5.0) and an average weight of 4.75g (SD=1.49, n=5) (Table 5).
Pigmentation: The exceptional pigmentation during the larval with sack stage and before preflexion showed two lines of olive and yellow colors in the margin of the notochord. Dark pigmentation was noticed on the body, the posterior side of the head and on the base of the pectoral fin. Five post-anal melanophores were also displayed on myomere five and six. During pre-flexion and before flexion, anal melanophores increased from 13 to 16, starting on the second postanal myomere and ending before reaching the tail. Eyes were completly pigmented with a silver-plated coloration.
During flexion, pigmentation had a notorious silver color in the intestine. The myomerehad a beige coloration going from the posterior cephalic region to the beginning of the tail. During the transition between this stage and the postflexion stage, the larva showed dark pigment and yellowish spots in this same region, while the silver plated coloration remained in the intestines. A total of 10 melanophores were observed on the base of the caudal fin. Prejuveniles presented an initial pigmentation with dark spots on their maxilla and jaw and dispersed on the head. In the medium and posterior parts of the operculum three dark pigments were observed. On the dorsal fin pigmentation went from the base of the first and second spine to the margin while on the rays pigmentation went towards the vertex and continued until the beginning of the caudal peduncle. Ten separated colors were observed towards the base of the anal fin. Pigmentation at the beginning of the caudal peduncle merged, fading before the beginning of the tail, where the base and the margins of the radii were pigmented. Finsturned to a brownish olive color with black margins. The lateral line of the body had yellowish pigmentation from the later part of the operculum to the caudal peduncle, which is a characteristic trait of the species (González 2005). In juveniles the same pigmentation pattern remained on the head and on the dorsal and anal fins, fading in the caudal peduncle.
Discussion
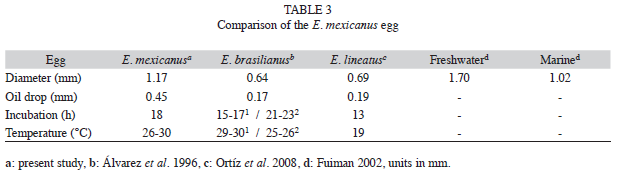
The descriptive characteristics of the embryonic stage of E. mexicanus are typical of this developmental period for fish of the Gerreidae family, particularly the species E. brasilianus and E. lineatus that inhabit marine environments (Álvarez et al. 1996, Ortíz et al. 2008). However, their morphometric characters are different with respect to the diameter and size of the oil drop, as E. mexicanus eggs are bigger than those of marine species, being similar to those of freshwater fish (Fuiman 2002).
The incubation period in hours is similar to that of marine species of the genus Eugerres and the incubation temperature is also similar to that reported for E. brasilianus, but greater than the one recorded during the incubation period for E. lineatus. Regarding the recently hatched larvae, E. mexicanus were bigger (average length of 2.94mm) than E. brasilianus (average length of 1.43mm). This may be observed through the average length of the vitelline sack of E. mexicanus and E. brasilianus measuring 0.98mm and 0.70mm, respectively, and the notochord length of E. lineatus (1.05mm) in relation to the larvae of E. mexicanus (2.74mm) (Álvarez et al. 1996, Ortíz et al. 2008).
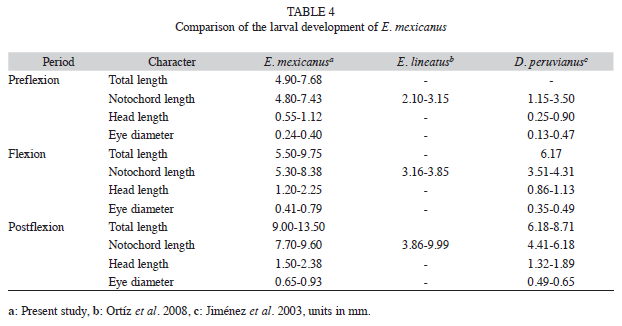
Following the classification of pre-flexion, flexion and post-flexion established to determine the development of larvae, the E. mexicanus larvae were bigger than the la rvae of the species E. brasilianus and E. lineatus of the same genus and the marine larvae of D. peruvianus (Álvarez et al. 1996, Jiménez et al. 2003, Ortíz et al. 2008).
It must be noted that Fuiman (2002) stated that the larvae of marine species are smaller in length than freshwater larvae, with averages of 2.87mm and 5.40mm. E. mexicanus prejuveniles of E. mexicanus measured 13.25mm to 15.00mm in length, similar to D. peruvianus (8.72 to 15.00mm) and E. lineatus prejuveniles (13.00mm to 18.00mm). On the other hand, E. mexicanus juveniles were bigger in length (28.16mm) than that recorded for E. lineatus (17.30mm) and D. peruvianus (17.00mm to 24.00mm) (Jiménez et al. 2003, Ortíz et al. 2008) (Table 5). With respect to the meristic characters, the fins of E. lineatus and D. peruvianus presented a different number of spines and rays during the larval phase. Finally, the juvenile phase presented spines and rays characteristic of E. mexicanus, as described by Deckert & Greenfield (1987) and González (2005).
Regarding pigmentation, the colors of the larva with sack for Eugerres mexicanus wereolive and yellow on the margin of the notochord. This differs from pigmentation reported for E. lineatus since in this sample there is a group of melanophores between myomere 9 to 13 as the main characteristic, and for Diapturus peruvianus because of the three spots displayed in the dorsal margin of the intestines from the insertion of the pectoral fin to the anus (Jiménez et al. 2003, Ortíz et al. 2008). E. mexicanus larvae during flexion followed the same pattern of coloration as the eleuthero embryo; however, during pre-flexion larvae contrasted with E. lineatus, since the latter continues with a similar pigmentation during pre-flexion and flexion, in which only the first melanophores are accentuated. Nevertheless, although E. mexicanus shows similar post-anal spots from 13 to 16 as D. peruvianus, it differs in that one or two of those spots are where the caudal fin will be (Jiménez et al. 2003, Ortíz et al. 2008).
Three spots were observed in the caps, and dark spots remained in the margin of the dorsal fin, which differs from E. peruvianus since in E. mexicanus two groups of fouropercular pigments are present and there is little pigmentation toward the corners of the dorsal fin while in D. peruvianus fins turn yellow and the body line is covered with silver flakes (Jiménez et al. 2003, Ortíz et al. 2008). In E. mexicanus prejuveniles brown pigmentation was observed in the cephalic region, the dark margins of the dorsal and anal fins, and the base of the tail, while pigmentation was yellow in the lateral line. Pigmentation in the mouth, specifically the maxilla and the jaw, is intensified towards the juvenile stage. Three pigments were observed in the operculum. The dark spots remain in the margin of the dorsal fin, which differs from E. peruvianus since in E. mexicanus two groups of four operculum pigments are present and there is little pigmentation towards the vertices of the dorsal fin, while in D. peruvianus fins turn yellow and the line of the body is covered with silver flakes (Jiménez et al. 2003, Ortíz et al. 2008).
Physicochemical parameters played an important role during white mojarra reproduction, as was the case of temperature that was favorable during the spawning and hatching stages. However, it varied from 25.6 to 28.4°C during the larval stage, which may have contributed to the high mortality recorded during this stage (99%). Álvarez et al. (1996) stated that a temperature of 20°C increased mortality in the case of E. brasilianus and also recorded a pH of 7.5 to 8.2 and values of dissolved oxygen of 5.5 to 7.1mg/L, similar to the pH and dissolved oxygen values required for E. mexicanus of 7.5 to 8.4 and 3.8 to 6.7mg/L, respectively. The nitrogen compounds recorded in the present study showed low levels and were considered non-toxic for the fish. Conductivity was low and varied from 0.51 to 0.56μS/cm. The foregoing allowed for the conclusion that, in general, there was no organic pollution during the reproduction, spawning, and embryonic and larval development, as Arredondo & Palafox (1998) mentioned for fish culture in controlled aquacultural systems. Food provided was assumed to be the cause for the recorded mortality, as it was based solely on microscopic green algae of the genus Crucigenia sp., collected from artificial tanks. Álvarez et al. (1996) provided the green alga Nannochloropsis oculata to E. brasilianus larvae at the beginning of their feeding, and later switched to Artemia sp. nauplii and copepods, obtaining a 50% survival.
Eugerres mexicanus specimens spawned after a single dose of 50UI/100g/fish of human chorionic gonadotropin hormone. In contrast, studies carried out in Tunas de Zaza, Cuba, on the induced reproduction of E. brasilianus used 6-10UI/g/fish distributed over two and three doses (Álvarez et al. 1996). In other cases, doses of 3.4-6.3 UI/g/fish were applied to females of Lutjanus griseus (Cabrera et al. 1997), and three doses of hormones were used at concentrations of 3.00, 1.00 and 0.5UI/g/fish for females and of 1.50, 0.50 and 0.25UI/g/fish for males of Haemulon bonarinense (Cuartas et al. 2003), with the doses of 0.50 (females) and 0.25 (males) producing the most viable sexual products. Only the 0.50 dose presented similar results to those obtained for E. mexicanus. Álvarez & Hernández (2001) determined that a period of 10 hours for spawning is relatively short in the case of tropical species, and related this to the short physiological action of the hormones at high environmental temperatures. However, in this study, spawning occurred 48 hours after injection, at temperatures of 29-31°C, indicating that spawning depends on the physiological characteristics of each species in relation to the temperature of the water. Successful egg fertilization (90-95%) in this study is similar to that recorded for E. brasilianus, which equaled to an average 87% success rate (range=60-100%) (Álvarez et al. 1996). Records of other fish species include Lutjanus griseus with 36% and 86% with the use of GCH (Cabrera et al. 1997) and Haemulon donariense with 90% (Cuartas et al. 2003). Other authors used the synthetic analogue of the luteinizing hormone releasing hormone (LHRHA) and obtained 85% for Orthopristis ruber (Mata et al. 2004) and 90% for Centropomus parallelus (Álvarez et al. 2002).
The results of this study are the first recorded for this species and generated information on several aspects of its reproductive biology and early life development. These results have also made it possible to establish a technique for manipulation in captivity, which may be adopted for aquaculture development of this species.
Acknowledgments
The authors acknowledge the CONACYT Fund and the government of the State of Tabasco for financing study TAB-2007-C09-73534.
References
Álvarez, L.L, L.P. Sánchez, O.G.H. Molejón & E.T. Gómez. 1996. Mass production of striped patio Eugerres brasilianus juveniles in Cuba. J. World Aquac. Soc. 27: 347-352. [ Links ]
Álvarez, L.L. & O.G.M. Hernández. 2001. Producción de juveniles de peces estuarinos para un centro en América Latina y el Caribe: diseño, operación y tecnologías. The World Acuaculture Society, Baton Rouge, Lousiana, USA. [ Links ]
Álvarez, L.L., V.R. Cerqueira & M. dos-Reis. 2002. Desarrollo embrionario y primeros estadios larvales del robalo chucumite, Centropomus parallelus poey (Pisces, Centropomidae) con interés para su cultivo. Hydrobiology 12: 89-100. [ Links ]
Arredondo, F. & P. Palafox. 1998. Calidad del agua en acuicultura. AGT editor, Mexico D.F., Mexico. [ Links ]
Cabrera, B.T., J.C. Rosas & J. Millán O. 1997. Reproducción y desarrollo larvario del pargo dientón (Lutjanus griseus L. 1758) (Pisces: Lutjanidae) cultivado en cautiverio. J. Sci. 33: 239-245. [ Links ]
Castro, A.J.L., H.P. Hernández & J.J.S. Schmitter. 1999. Ictiofauna Estuarino-Lagunar y Vicaria de México. Limusa-Noriega, Mexico. [ Links ]
Cuartas, A., J. Rosas, A. Velásquez & T. Cabrera. 2003. Inducción al desove, desarrollo embrionario y larval de corocoro rayado Haemulon bonariense Cuvier, 1830 (Pisces: Haemulidae). Rev. Biol. Mar. Ocean. 38: 27-37. [ Links ]
Deckert, G.D. & D.W. Greenfield. 1987. A Review of the Western Atlantic Species of the Genera Diapterus & Eugerres (Pisces: Gerreidae). Copeia 1987: 182-194. [ Links ]
Fuiman, L.A. 2002. Special considerations of fish eggs and larvae, p. 337. In L.A. Fuiman & R.G. Werner (eds.). Fishery Science Unique Contributions of Earth Life Stages Blackwell, Oxford, England. [ Links ]
Gilmore, R.G., Jr. V. Beach & D.W. Greenfield. 2002. Gerreidae. The living marine resource of the Western Central Atlantic. Vol. 3: Bony Fishes, Part 2 (Opistognathidae to Molidae), sea turtles and marine mammals. [ Links ]
González, A.A.F. 2005. Estudio Sistemático y Biogeográfico del género Eugerres (Perciformes: Gerreidae). Tesis de Doctorado, CICIMAR, La Paz, Baja California Sur, Mexico. [ Links ]
González, A.A.F. & R.H. Rodiles. 2007. Sobre el estatus taxonómico de la mojarra blanca Eugerres mexicanus (Steindachner, 1863). Mesoamericana 11: 52-53. [ Links ]
González, A.A.F., J. de la Cruz-Agüero & J.L. Castro- Aguirre. 2007. A review of the marine Western Atlantic species of the genus Eugerres (Perciformes: Gerreidae). Bull. Mar. Sci. 80: 109-124. [ Links ]
Hardy, J.D. Jr. 1978. Development of fishes of the Mid- Atlantic Bight: An Atlas of egg, larval and juvenile stages. Volume II. Anguillidae through Syngnathidae. U.S. Fish and Wildlife Service, Biological Services Program, FWS/OBS-78/12. [ Links ]
Jiménez, R.S.P.A., E.G. Navarro & R.J.S. Martínez. 2003. Larval, pre-juvenile and juvenile development of Diapterus peruvianus (Perciformes: Gerreidae). Rev. Biol. Trop. 51: 479-488. [ Links ]
Mata, E., J. Rosas, A. Velásquez & T. Cabrera. 2004. Inducción hormonal al desove y descripción larval del corocoro Orthopristis ruder Cuvier (Pisces: Haemulidae). Rev. Biol. Mar. Ocean. 39: 21-29. [ Links ]
Miller, R.R., W.L. Minckley & S.M. Norris. 2005. Freshwater fishes of Mexico. The University of Chicago, Chicago, USA. [ Links ]
Ortíz, G., J.L. Castro-Aguirre, E.F.F. Balart & N.A. Pliego. 2008. Description of early ontogeny and osteological development of streaked mojarra Eugerres lineatus (Teleostei: Percoidei:Gerreidae). Zootaxa 1911: 1-30. [ Links ]
Rosas, J., E. Mata, A. Velásquez & T. Cabrera. 2008. Desarrollo embrionario-larval del pez tropical Hemirhamphus brasiliensis (Beloniformes: Hemirhamphidae) a partir de huevos recolectados. [ Links ]
*Correspondencia:
Raúl E. Hernández: Ingeniería en Acuacultura, División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, km 1 carretera Tenosique-Estapilla, Tenosique, Tabasco, México. CP 86901; raul.hernandez@ujat.mx
Martha A. Perera: Ingeniería en Acuacultura, División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, km 1 carretera Tenosique-Estapilla, Tenosique, Tabasco, México. CP 86901.
Alfonso Castillo: Ingeniería en Acuacultura, División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, km 1 carretera Tenosique-Estapilla, Tenosique, Tabasco, México. CP 86901.
Emiliano Luna H:Ingeniería en Acuacultura, División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, km 1 carretera Tenosique-Estapilla, Tenosique, Tabasco, México. CP 86901.
José A. de la Cruz:Ingeniería en Acuacultura, División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, km 1 carretera Tenosique-Estapilla, Tenosique, Tabasco, México. CP 86901.
Luis M. Gómez: División Académica de Ciencias Agropecuarias, Universidad Juárez Autónoma de Tabasco; luisgomezdd@gmail.com.mx
José Valdez Zenil: Facultad de Biología de Ciencias Biológicas y Agropecuarias, Universidad Veracruzana, carretera Tuxpan-Tampico, Tuxpan, Veracruz; josevaldezz@yahoo.com.mx
1. Ingeniería en Acuacultura, División Académica Multidisciplinaria de los Ríos, Universidad Juárez Autónoma de Tabasco, km 1 carretera Tenosique-Estapilla, Tenosique, Tabasco, México. CP 86901; raul.hernandez@ujat.mx
2. División Académica de Ciencias Agropecuarias, Universidad Juárez Autónoma de Tabasco; luisgomezdd@gmail.com.mx
3. Facultad de Biología de Ciencias Biológicas y Agropecuarias, Universidad Veracruzana, carretera Tuxpan-Tampico, Tuxpan, Veracruz; josevaldezz@yahoo.com.mx
Received 25-X-2010. Corrected 09-VII-2011. Accepted 09-VIII-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}