











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.60 no.1 San José mar. 2012
Distribución de los gasterópodos del manglar, Neritina virgínea (Neritidae) y Littoraria angulifera (Littorinidae) en la Ecorregión Darién, Caribe colombiano
Abstract
Distribution of the mangrove gastropods Neritina virginea (Neritidae) and Littoraria angulifera (Littorinidae) within the Colombian Caribbean Darién Ecoregion. Gastropods are one of the most abundant groups within the Caribbean mangroves, however, little is known about the distribution of particular species at a regional scale. With this aim, we studied the geographic distribution of Littoraria (Littorinopsis) angulifera and Neritina (Vitta) virginea within the Darién Ecoregion in the Caribbean coast of Colombia, from 77 sampling stations along 609km between the Colombian-Panamá border and Córdoba State, Colombia. The fieldwork was conducted in June-August 2009, and a total of 3 963 individuals of both species were hand-picked from the ground, prop-roots and trunks along 50m transects, and shell sizes were measured. The description of geographic patterns considered surface water salinity, mangrove cover and gastropod distribution within the Gulf of Urabá. In the outer-most part of the Gulf, L. angulifera was present in 84.8% of the stations, while N. virginea was only present in 15.2% of the stations. In this part, mangroves areas were patchily distributed, and the gastropods (mainly L. angulifera) were found on woody debris along the supralittoral zone in sandy shores. In the inner-most part, in contrast, N. virginea occurred in 84.6% of the stations, mostly in estuaries, deltas and river margins, while L. angulifera only appeared in Turbo Bay (15.4%). Mean shell size also exhibited a clear geographic pattern: size range was 6-22mm in L. angulifera, and 6-12mm in N. virginea. L. angulifera was found in open-water stations with water salinities >10PSU, but it was absent in sites with lower salinities like the Atrato River Delta and other small rivers. Its presence on coastal woody debris suggests that despite of the recruitment of small individuals from the nearshore stock of larvae, populations are unable to establish due to the absence of mangroves protection. Oppositely, N. virginea was found under estuarine conditions on mangrove roots and ground. Our results confirm that L. angulifera is an esteno-tolerant marine species, and N. virginea is an eury-tolerant estuarine species, thus their geographic distribution is strongly shaped by the large freshwater discharge of the Atrato River. We hypothesize that absence or limited distribution of gastropods in various areas of the Darién Ecoregion may be further explained by the poor conservation state of mangroves.
Key words: mangrove gastropods, marine biogeography, shell size, estuary, Darién ecoregion, Southern Caribbean.
Resumen
Aunque los gasterópodos son uno de los grupos más abundantes en los manglares del Caribe, poco se sabe del ámbito de especies particulares debido al enfoque comunitario de la mayoría de los estudios. El ámbito de las poblaciones de L. angulifera y N. virginea fue estudiado en la Ecorregión Darién, Caribe colombiano. El muestreo se realizó entre junio y agosto 2009, se recolectaron 3 963 individuos de ambas especies, a los cuales se les midió la talla de la concha. La Ecorregión presentó diferencias espaciales en el ámbito de la salinidad del agua a nivel superficial, la cobertura de manglar y el ámbito de las poblaciones. En la parte externa de la Ecorregión, L. angulifera presentó el mayor porcentaje de aparición (84.8%) en comparación a N. virginea (15.2%). En el interior N. virginea presentó el mayor porcentaje (84.6%), mientras L. angulifera apareció solo en bahía Turbo (15.4%). Se encontraron diferencias en el ámbito de tallas promedio de las conchas, donde L. angulifera varió entre 6 y 22mm y N. virginea entre 6 y 12mm dentro del Golfo. Se confirma que L. angulifera es una especie de carácter estenohalina y marina, mientras que N. virginea es eurihalina y estuarina.
Palabras clave: gasterópodos del manglar, biogeografía marina, tamaño de la concha, estuario, Ecorregión Darién, Caribe sur.
Distribution of the mangrove gastropods Neritina virginea (Neritidae) and Littoraria angulifera (Littorinidae) within the Colombian Caribbean Darién Ecoregion. Gastropods are one of the most abundant groups within the Caribbean mangroves, however, little is known about the distribution of particular species at a regional scale. With this aim, we studied the geographic distribution of Littoraria (Littorinopsis) angulifera and Neritina (Vitta) virginea within the Darién Ecoregion in the Caribbean coast of Colombia, from 77 sampling stations along 609km between the Colombian-Panamá border and Córdoba State, Colombia. The fieldwork was conducted in June-August 2009, and a total of 3 963 individuals of both species were hand-picked from the ground, prop-roots and trunks along 50m transects, and shell sizes were measured. The description of geographic patterns considered surface water salinity, mangrove cover and gastropod distribution within the Gulf of Urabá. In the outer-most part of the Gulf, L. angulifera was present in 84.8% of the stations, while N. virginea was only present in 15.2% of the stations. In this part, mangroves areas were patchily distributed, and the gastropods (mainly L. angulifera) were found on woody debris along the supralittoral zone in sandy shores. In the inner-most part, in contrast, N. virginea occurred in 84.6% of the stations, mostly in estuaries, deltas and river margins, while L. angulifera only appeared in Turbo Bay (15.4%). Mean shell size also exhibited a clear geographic pattern: size range was 6-22mm in L. angulifera, and 6-12mm in N. virginea. L. angulifera was found in open-water stations with water salinities >10PSU, but it was absent in sites with lower salinities like the Atrato River Delta and other small rivers. Its presence on coastal woody debris suggests that despite of the recruitment of small individuals from the nearshore stock of larvae, populations are unable to establish due to the absence of mangroves protection. Oppositely, N. virginea was found under estuarine conditions on mangrove roots and ground. Our results confirm that L. angulifera is an esteno-tolerant marine species, and N. virginea is an eury-tolerant estuarine species, thus their geographic distribution is strongly shaped by the large freshwater discharge of the Atrato River. We hypothesize that absence or limited distribution of gastropods in various areas of the Darién Ecoregion may be further explained by the poor conservation state of mangroves.
Key words: mangrove gastropods, marine biogeography, shell size, estuary, Darién ecoregion, Southern Caribbean.
Resumen
Aunque los gasterópodos son uno de los grupos más abundantes en los manglares del Caribe, poco se sabe del ámbito de especies particulares debido al enfoque comunitario de la mayoría de los estudios. El ámbito de las poblaciones de L. angulifera y N. virginea fue estudiado en la Ecorregión Darién, Caribe colombiano. El muestreo se realizó entre junio y agosto 2009, se recolectaron 3 963 individuos de ambas especies, a los cuales se les midió la talla de la concha. La Ecorregión presentó diferencias espaciales en el ámbito de la salinidad del agua a nivel superficial, la cobertura de manglar y el ámbito de las poblaciones. En la parte externa de la Ecorregión, L. angulifera presentó el mayor porcentaje de aparición (84.8%) en comparación a N. virginea (15.2%). En el interior N. virginea presentó el mayor porcentaje (84.6%), mientras L. angulifera apareció solo en bahía Turbo (15.4%). Se encontraron diferencias en el ámbito de tallas promedio de las conchas, donde L. angulifera varió entre 6 y 22mm y N. virginea entre 6 y 12mm dentro del Golfo. Se confirma que L. angulifera es una especie de carácter estenohalina y marina, mientras que N. virginea es eurihalina y estuarina.
Palabras clave: gasterópodos del manglar, biogeografía marina, tamaño de la concha, estuario, Ecorregión Darién, Caribe sur.
Los gasterópodos son uno de los grupos de invertebrados epibénticos más conspicuos y abundantes asociados al suelo, raíces y troncos de los manglares en todo el mundo (Hogarth 2007, Lee 2008). Aunque la malacofauna del manglar ha sido ampliamente estudiada en muchas partes del mundo, aún existen muchos vacíos sobre la autoecología de muchas especies debido al desequilibrio geográfico de los estudios y a las dificultades logísticas para su muestreo y cuantificación (Lee 2008). Aunque en el Caribe y el Atlántico existen varios estudios sobre la malacofauna del manglar (Ellison 2008), los estudios a nivel poblacional son escasos (Merkt & Ellison 1998, Tanaka & Maia 2006). Adicionalmente, existen numerosos estudios sobre la epifauna asociada a las raíces (Ellison 2008), pero es poco lo que se conoce de la asociada a los sedimentos, posiblemente porque permanecen sumergidos la mayor parte del tiempo debido al régimen micromareal.
En el Caribe Colombiano, la malacofauna asociada al manglar ha sido ampliamente estudiada, siendo la ciénaga Grande de Santa Marta y su vecindad una de las áreas más exploradas desde los años setentas (Cosel 1986, Díaz 1994, 1995, Palacio 1978, 1983). La bahía de Cartagena y el golfo de Morrosquillo también han sido históricamente estudiados (revisado por Ortiz & Blanco 2010, Blanco et al. 2010), mientras que el Archipiélago de San Andrés y Providencia ha recibido atención durante la última década (Vilardy & Polanía 2002). Sin embargo, la Ecorregión Darién (Caribe colombiano sur) ha sido poco explorada. Dentro de ésta, se han realizado trabajos sobre la dinámica de la comunidad de macroinvertebrados epibiontes de las raíces en áreas geográficas puntuales del golfo de Urabá (bahía Marirrío: Llano 1998, bahías El Uno y Turbo: García & Palacio 2008). Los dos únicos estudios que han abarcado una porción significativa del golfo de Urabá son el de Rojas & Palacio (2003) y el de Ortiz & Blanco (2010). Sin embargo, el estudio de Ortiz & Blanco (2010) es hasta ahora el de mayor extensión (49 estaciones a lo largo de 609km de costa) y mayor esfuerzo de muestreo (245 raíces, 69 932 individuos). Este estudio, enfocado en los moluscos epibénticos de las raíces de los mangles, confirmó que esta taxocenosis es pobre (ocho especies) dentro de la Ecorregión Darién en comparación al resto del Caribe colombiano y al Gran Caribe, con una dominancia del 94% por parte de dos bivalvos (Mytilopsis cf. sallei y Brachidontes cf. exustus). Este estudio también evidenció que la mayor parte de las especies eran eurihalinas posiblemente debido a la predominancia de condiciones estuarinas en el Golfo la mayor parte del año.
Debido a que la Ecorregión Darién es: a) un área poco conocida en la literatura internacional, b) alberga un área significativa de manglar (6 512ha, García 2007), y c) recibe uno de los aportes de agua dulce más grandes de todo el Caribe sur a través del río Atrato, se decidió explorar los patrones de arreglo espacial a la mesoescala (105m) de Littoraria (Littorinopsis) angulifera (Lamarck, 1822) y Neritina (Vitta) virginea (L.), dos gasterópodos abundantes en los suelos y raíces emergidos de los manglares de la Ecorregión Darién, al igual que en el Gran Caribe (Palacios 1978, Díaz 1994). Adicionalmente, los manglares de la Ecorregión Darién han sido objeto de múltiples presiones a través de los años, por lo cual el reciente reconocimiento de su gran valor ecosistémico y de las múltiples funciones de sus componentes, han llevado a tomar acciones para un manejo sustentable y su conservación (García 2007). Específicamente, se formularon las siguientes preguntas: 1) ¿Cuál es el ámbito de distribución de las poblaciones de L. angulifera y N. virginea en el área de estudio? 2) ¿Cómo varía la estructura de tallas de las poblaciones de ambos gasterópodos? 3) ¿Existe relación entre dichas variables con la variación espacial de la salinidad y el estado de conservación de los manglares?
Materiales y métodos
Especies estudiadas. El gasterópodo N. virginea se encuentra desde Florida y Bahamas hasta el norte de Brasil, y desde el Golfo de México y América Central hasta Las Antillas. El tamaño oscila entre 4 y 25mm, desde los individuos recientemente reclutados hasta los adultos más grandes. Los juveniles y adultos exhiben una coloración verde con un patrón variable de líneas axiales o de pequeños triángulos amarillos. Coloniza las praderas de fanerógamas y los manglares de lagunas costeras, bahías y golfos, pero también se encuentran densas poblaciones desde la desembocadura de los ríos hasta varios kilómetros aguas arriba. También coloniza pequeños ríos y quebradas costeras, presentando un ciclo de vida diádromo. En este ciclo la larva recién eclosionada del huevo es arrastrada aguas abajo hasta el mar. La larva es planctotrófica y se cree que puede durar varios meses a la deriva transportada por las corrientes marinas (Crandall 1999, Blanco & Scatena 2005, 2006, 2007). Al llegar a las costas cercanas a los estuarios, las larvas se transforman en reclutas bénticos que empiezan a migrar río arriba, proceso que dura toda su vida. Al alcanzar la madurez sexual ponen cápsulas ovígeras con cientos de huevos. Información detallada sobre la ecología de la especie en quebradas costeras del Caribe ha sido publicada por Blanco & Scatena (2005, 2006, 2007) y Blanco & Arroyave (2009). En los manglares los individuos hacen migraciones horizontales y verticales dependiendo de los ciclos mareales diarios y mensuales, y se encuentran sobre el fango, hojarasca, neumatóforos y las partes bajas de las raíces aéreas y troncos (J.F. Blanco, pers. observ.).
L. angulifera habita en las zonas supralitorales de los manglares del Atlántico y el Caribe (Tanaka & Maia 2006). Se encuentra principalmente en hojas, troncos y raíces adventicias de los mangles (Rhizophora mangle L., Avicennia germinans, Avicennia marina, Avicennia alba, Avicennia nitida y Laguncularia racemosa) donde se alimentan de algas y hongos marinos (Merk & Ellison 1998). Este gasterópodo es ovivíparo incompleto, con desarrollo larval en la cavidad corporal de la hembra hasta el estado velígero, el cual es liberado en la columna de agua (Gaines et al. 1974). El estado larval planctotrófico tiene un periodo de duración aproximado de 8-10 semanas (Merk & Ellison 1998).
Área de estudio. La Ecorregión Darién en el Caribe comprende el sur de Panamá y Colombia. En el extremo suroccidental del Caribe colombiano, la zona costera está comprendida por los departamentos de Chocó y Antioquia, donde sobresalen las subregiones Caribe chocoano, golfo de Urabá y Caribe antioqueño. El golfo de Urabá (7º50’ - 8º56’ N y longitud: 77º22’ - 76º25’ W) es la más extensa, siendo un cuerpo de agua semicerrado, alargado en dirección norte-sur con una morfología similar a la letra U (aproximadamente 80km de largo y 25km de ancho promedio, con cerca de 8km en su parte más estrecha). Tiene un área de 4 291km2, y es la entrada más grande del mar Caribe en tierra colombiana (García 2007). El Golfo es el estuario de mayor tamaño en el Caribe colombiano ya que en la parte media de su costa occidental desemboca el río Atrato (4 155m3/s), el de segundo mayor caudal del Caribe colombiano (después del río Magdalena), y que exhibe un extenso delta digitado. En el costado suroriental desembocan varios ríos de menor caudal (Turbo, Guadualito, Currulao y León) que forman microdeltas acuminados moldeados por el oleaje.
La estacionalidad de los vientos y de la precipitación (época húmeda y época seca) juegan un papel importante en la distribución espacial de la salinidad. La circulación de los vientos en el área está en fase con el movimiento de la Zona de Convergencia Inter-Tropical cuyo desplazamiento hacia el sur de Colombia en la época seca (diciembre-enero) trae consigo un aumento de los vientos Alisios del noreste, mientras que cuando ésta se desplaza hacia el norte en la época húmeda, se presentan los vientos Alisios del sureste. En la época seca, aunque las precipitaciones son muy bajas, el aumento de los vientos Alisios del norte y nordeste retienen el aporte de agua continental ocasionando que el Golfo se comporte como una enorme “piscina” de agua oligo- a mesohalina a nivel superficial. En la época húmeda los vientos son débiles y se caracterizan por soplar principalmente del sur y suroeste. Además, se presentan fuertes precipitaciones, por lo cual el aporte de agua dulce proveniente delrío Atrato y de otros ríos de menor magnitud es exportado del Golfo, y permanece una masa de agua más salada (polihalina) al interior (bahía Colombia) (García 2007, Osorio et al. 2009).
El delta del río Atrato es el área de mayor extensión y desarrollo de manglar dominada por Rhizophora mangle, con predominio de 80% (densidad relativa), seguida de Laguncularia racemosa. En bahía Colombia se presenta un manglar de tipo ribereño con mezcla de varias especies (río Suriqui). En la costa occidental del Golfo existen pequeñas áreas de manglar de franja, con Rhizophora mangle ubicada en el borde de la costa y Avicennia germinans al interior. La ensenada de Rionegro es una laguna costera ubicada en el límite norte del Golfo, es la mayor área de manglar de la costa oriental, dominada por Rhizophora mangle y Laguncularia racemosa. En el costado suroriental sobresalen los parches de manglar en la desembocaduras de los ríos León y Currulao y en las bahías Turbo y El Uno. Finalmente, en la costa Caribe antioqueña hay unos relictos (caño La Marimonda, río Damaquiel, río Hobo y río Volcán) producto de la actividad antropogénica y la erosión costera (CORPOURABÁ 2003). Para una descripción más detallada de la Ecorregión fue realizada una reciente expedición (Blanco et al. 2011).
Muestreo. Se realizaron dos salidas de campo (3-14 de junio y 3-10 de agosto de 2009) para verificar la existencia de poblaciones de N. virginea y L. angulifera y construir mapas de ámbito de distribución recolectaron entre 50 y 200 individuos al azar de cada especie sobre el suelo, raíces o troncos a lo largo de transectos de 50m dentro de cada sitio de muestreo. Durante 20 minutos se recolectaron entre 50 y 200 individuos al azar de cada especie sobre el suelo, raíces o troncos a lo largo de transectos de 50m dentro de cada sitio de muestreo. Para preservar los individuos recolectados se fijaron en una solución de formol al 10%. En la primera salida de campo se visitaron 39 áreas geográficas en búsqueda de zonas de manglar en los departamentos de Chocó y Antioquia entre cabo Tiburón en el límite con Panamá (al occidente) y quebrada Peñoncito en el límite con el departamento de Córdoba (al oriente). Estas áreas de manglar habían sido identificadas previamente mediante el uso de mapas, imágenes satelitales, aerofotografías y conceptos de expertos. En la segunda salida, se enfocó el muestreo en 32 estaciones dentro del delta del río Atrato (la mayor área de manglar) y dentro de las bahías El Uno y Turbo (Fig. 1).En las desembocaduras de algunas quebradas donde los manglares habían desaparecido, se buscaron y recolectaron individuos de L. angulifera sobre troncos en las bermas de las playas.
En el laboratorio, se midieron las conchas de todos los individuos. A L. angulifera se le midió la longitud axial y a N. virginea se le midió el ancho de la concha sobre la columela utilizando un pie de rey con una precisión de 0.01mm.
En el campo se midió la salinidad o la conductividad dependiendo de la disponibilidad de instrumentos. En los casos que no se obtuvo valor de campo, se estimó utilizando los valores promedios reportados para dicha época climática en el “Atlas del golfo de Urabá” (García 2007). El ámbito de salinidad medido en el campo fue comparado con el reportado en la literatura para validar la interpolación de los datos faltantes.
Los valores georeferenciados de salinidad, áreas de manglar y poblaciones de gasterópodos se proyectaron en el sistema de información geográfica ArcGIS versión 9.3. Los polígonos con las áreas de cobertura de manglares se obtuvieron del inventario más reciente, el cual utilizó ortofotografías a escala 1:10 000 (Urrego et al. 2010, Blanco et al. 2011). Para contestar la pregunta uno, con los datos de frecuencia de las estaciones donde estaba presente cada especie se realizó una tabla de contingencia y una prueba Chi cuadrado para determinar el ámbito de cada especie y si ellas estaban asociadas (Guisande et al. 2006, Ramírez 2005), mediante el uso del programa Statistica 6.0 (edición 98). Para contestar la pregunta dos, se realizó una prueba de Kolmogorov-Smirnof para establecer si las muestras de tallas presentaban distribución normal (Guisande et al. 2006, Ramírez 2005). Las tallas promedio se compararon entre estaciones de muestreo utilizando un Análisis de Varianza (ANOVA) de una vía (Guisande et al. 2006, Ramírez 2005). Los datos de cada salida se trataron en conjunto debido a que las condiciones climáticas y oceanográficas fueron similares. Se realizaron regresiones lineales (Guisande et al. 2006, Ramírez 2005) entre las tallas promedio y la salinidad (pregunta 3). Finalmente, se realizó una prueba t-student para comparar las tallas promedio de L. angulifera entre áreas con o sin manglar dentro del Caribe chocoano y antioqueño (Guisande et al. 2006, Ramírez 2005) (pregunta tres). Esta comparación no se realizó en N. virginea porque no se encontró afuera de los manglares de la Ecorregión.
Resultados
Ámbito de salinidad y áreas de manglar. La salinidad superficial observada osciló entre 0.1 y 37PSU. La parte interna del Golfo (delta del río Atrato) presentó aguas superficiales oligo- y meso-halinas con excepción de la bahía Turbo que presentó los valores más altos de salinidad (polihalina). La salinidad aumentó hacia el Norte como resultado de la mezcla de agua de escorrentia continental con el agua marina, arrojando como resultado aguas polihalinas (18-30PSU) y en algunos casos mixoeuhalinas (30-40PSU). Por lo tanto, las aguas costeras del Golfo presentaron un fuerte gradiente latitudinal de salinidad (Fig. 2).
Los manglares de la Ecorregión Darién cubren actualmente una extensión de 4 912ha. En la parte norte se presentan pequeños relictos de manglar con una cobertura de 33ha. En el Chocó (Titumate y Pinorroa) y el Caribe antioqueño (ríos Damaquiel y Hobo) se encuentran 16 y 17ha, respectivamente, que se ubican dentro de bahías y bocas de los caños y quebradas que los protegen de la erosión. A lo largo de estas subregiones se observaron árboles aislados y algunos talados, indicios de coberturas previas mayores que fueron taladas o erosionadas en algunas localidades. En el departamento de Antioquia, 3 846ha se encuentran en la costa occidental en el delta de río Atrato. La costa oriental presenta un total de 1 034ha, de las cuales 38ha se encuentran en el río Suriqui, 192ha en el río León y punta Coquito, 214ha en los ríos Guadualito y Currulao, 145ha en punta Yarumal y bahía Turbo, 103ha en Caimán Nuevo y 342ha en la ensenada de Rionegro (Fig. 2).
Ámbito espacial de las poblaciones de L. angulifera y N. virginea. Durante la fase de campo (77 estaciones de muestreo) se recolectaron un total de 3 963 individuos de ambas especies: 1 051 de L. angulifera y 2 912 de N. virginea.
El porcentaje de aparición de las poblaciones de cada especie varió considerablemente en el interior y exterior de la Ecorregión Darién. L. angulifera apareció en el 84.8% de las estaciones de la parte norte, mientras que N. virginea apareció solo en un 15.2% de las estaciones de esta parte, particularmente en los manglares protegidos del Chocó (Playa Soledad y Titumate) y la ensenada de Rionegro, el caño de la Marimonda y el río Damaquiel en el Caribe antioqueño. L. angulifera fue la especie dominante del exterior del Golfo (Fig. 3a). Al interior, es decir, en el golfo de Urabá, al sur de punta Yerbasal y Caimán Nuevo, se invirtieron los patrones. L. angulifera apareció en un 15.4% de las estaciones, pero estuvo presente sólo en bahía Turbo, mientras que N. virginea apareció en el 84.6% de las estaciones, en donde las poblaciones establecidas sólo se observaron en los manglares protegidos del oleaje y bien desarrollados de la parte interna(Fig. 3b). Esta especie fue numéricamente dominante en la parte interna.
La prueba Chi2 sobre una tabla de contingencia mostró que no había una asociación significativa entre las especies (p<0.001), por lo que resaltan notables diferencias del ámbito de las especies entre el interior y el exterior de la Ecorregión, a pesar de ser simpátricas en la meso-escala (105).
Ámbito de las tallas de L. angulifera y N. virginea. La muestra de tallas de L. angulifera para la Ecorregión Darién presentó una distribución normal (Prueba de Shapiro-Wilk, p>0.05), mientras que la muestra de tallas de N. virginea no (p<0.001). El ámbito de tallas de L. angulifera, presentó valores promedios entre 6 y 22mm. Este ámbito de tallas promedio de las poblaciones presentó diferencias significativas entre las estaciones de muestreo (F12,1037=61.45, p<0.001). Los individuos más grandes (promedio=21mm) se recolectaron en Titumate y se consideró como una población establecida. En playa Soledad y el río San Juan se observaron tallas promedio alrededor de 15mm, también se consideraron poblaciones establecidas. Sin embargo, estas dos poblaciones presentaron una proporción significativa de individuos <15mm. El resto de estaciones de muestreo presentaron promedios de talla <12mm. En la quebrada La Cartagenita, Pinorroa y caño La Marimonda se observaron los individuos de menor tamaño (≤5mm) (Fig. 4).
Las tallas de N. virginea presentaron una distribución leptocúrtica, con una mayor frecuencia de individuos entre 6 y 12mm (tallas promedio) en la Ecorregión. Se observaron, sin embargo, diferencias significativas entre áreas geográficas (F44, 2868=30.01, p<0.001) pero sin un patrón definido, aunque en el manglar protegido de la bahía El Uno presentó un valor de talla promedio de 12mm y en el manglar de bahía Cocogrande se encontraron las menores tallas promedio (<6mm) (Fig. 4).
Las tallas promedio de L. angulifera en la Ecorregión Darién no exhibieron una correlación significativa (r=0.02, p=0.96) con la salinidad, debido que se encontraron en una zona de alta salinidad y poca variabilidad. Las tallas promedio de N. virginea presentaron una correlación significativa con la salinidad dentro de la Ecorregión (r=0.38, p=0.04), pero no en la parte interna del Golfo (mayor resolución espacial: delta del río Atrato y bahías Turbo y El Uno) (r=0.16, p=0.52).
L. angulifera se encontró ampliamente distribuido dentro y fuera de los manglares de la costa norte, pero las tallas promedio fueron significativamente mayores en manglares establecidos (14±6mm) en el Caribe chocoano y antioqueño y no afuera de ellos (10±3.2mm) (t=4.48, g.l.=8, p=0.01). N. virginea presentó un ámbito limitado a las zonas de manglar, las mayores tallas y abundancias promedio se encontraron en el interior del Golfo.
Discusión
Condicionantes del ámbito de salinidad en la Ecorregión Darién. El continuo aporte de agua dulce del río Atrato al Golfo, confiere condiciones estuarinas durante todo el año, que lo divide en dos sectores (García 2007). El sector norte registra los valores más altos de salinidad, se clasifica como una zona de aguas polihalinas (18-30PSU) y mixoeuhalinas (30-40PSU), resultado de la influencia directa del agua marina y el poco aporte de agua dulce por los ríos de este sector. Los ríos más grandes son Mulatos, San Juan y Damaquiel pero sus caudales no están aforados. La ensenada de Rionegro, por su condición de laguna costera, puede presentar estacionalmente valores altos de salinidad. Rojas & Palacio (2003) informaron que en esta ensenada se midió el valor más alto de las 24 estaciones del litoral antioqueño. El sector sur del Golfo presenta salinidades bajas (0.1-7.9PSU), dado el gran y continuo aporte de agua dulce del río Atrato y otros ríos de menor caudal (ríos Suriquí, León, Currulao, Guadualito, Turbo, Caimán Nuevo y Caimán Viejo), y de corrientes menores (quebradas y caños) (García 2007). Al interior del Golfo, la bahía Turbo presenta valores de salinidad relativamente altos, como resultado del confinamiento de las aguas por la disposición latitudinalmente alargada de la bahía y la dirección del viento local predominantemente sur-norte. Otra posible causa de estas condiciones es un largo periodo de residencia y poca mezcla de sus aguas y la alta presión antrópica a la que está expuesta (puerto y asentamiento humano). Ramírez (2005) plantea que la actividad antrópica en las zonas costeras puede alterar las características fisicoquímicas de las aguas por medio de la contaminación puntual y difusa. Adicionalmente, el poco aporte de agua dulce que recibe y la alta evaporación podrían mantener una masa de agua de mayor salinidad al interior de la bahía, como sucede en otros estuarios del mundo.
Los valores de salinidad superficial observados durante nuestro estudio, fueron característicos de una época seca (junio y agosto). Estos meses normalmente corresponden a un periodo de transición, veranillo de mitad de año (García 2007), con predominancia de vientos débiles con dirección sur y suroeste, que convirten al golfo en una enorme piscina de agua dulce (García 2007).
La salinidad y el ámbito de los gasterópodos. La salinidad es la principal determinante de la distribución de la fauna asociada al manglar (Rojas & Palacio 2003, García & Palacio 2008) y a los estuarios (Vermeij 1971). Por ejemplo, en la ciénaga Grande de Santa Marta (Palacio 1978), la bahía Chengue (Reyes & Campos 1992 a, b), la bahía de Cartagena y la ciénaga Los Vásquez (Pérez & Victoria 1977) en el Caribe colombiano los cambios en la distribución de especies obedecen a cambios en la salinidad. Nuestros resultados apoyan parcialmente lo observado en otras áreas del Caribe colombiano, a pesar de los amplios ámbitos de las especies estudiadas. L. angulifera se encontró restringida a salinidades >15PSU en la parte norte (y disyuntamente en la bahía Turbo), pero N. virginea se encontró en salinidades entre 0 y 37PSU, y muestra un mayor ámbito. Sin embargo la variación de las tallas si es influenciada por la salinidad a la meso escala. La limitada presencia de esta segunda especie en el Caribe chocoano y antioqueño obedece aparentemente a otros factores como se explicará más adelante.
Ámbito de las tallas. Las tallas promedio de L. angulifera no mostraron cambios pronunciados con respecto a otros Littorinidos del Pacífico y el Atlántico (Tailandia, el Pacífico colombiano y Brasil), en donde los adultos oscilan entre 16 y 22mm (Sanpanich et al. 2008), los juveniles entre 9 y 16mm, y los reclutas 6 y 9mm (Tailandia: Sanpanich et al. 2008; Brasil: Tanaka & Maia 2006; Pacífico Colombiano: Blanco & Cantera 1999 y Riascos & Guzmán 2010). En el presente estudio, las tallas promedio de L. angulifera sugieren que hay pocas localidades con poblaciones compuestas por adultos y juveniles, pero la mayoría de las localidades presentan tallas promedio que indican la prevalencia de juveniles y reclutas y la ausencia de adultos. Esto podría ser un indicador de la estabilidad de la población asociada con el estado de conservación del manglar, como se discutirá más adelante.
De acuerdo al ámbito de tallas promedio de las poblaciones es posible que algunas estén relativamente aisladas. Las poblaciones de playa Soledad y Titumate (Caribe chocoano) fueron las de mayor talla promedio y las únicas que se encontraron dentro de manglares relativamente extensos, y mantienen, al menos parcialmente, el aporte local de larvas. Por el contrario, las poblaciones de individuos de tallas mucho menores que se encontraron sobre las bermas de las playas del Caribe chocoano y antioqueño, dependen del aporte extrínseco de larvas, las cuales pueden ser transportadas desde las localidades próximas hacia éstas, por el oleaje y corrientes del Caribe, el cual está dominado fuertemente en este sector por el oleaje tipo Swell (comportamiento independiente entre el oleaje y la magnitud de los vientos) (Osorio et al. 2010).
La talla promedio de N. virginea presentó una correlación significativa con la salinidad en la Ecorregión Darién. Las tallas observadas se encontraron dentro del ámbito observado en Puerto Rico por Blanco & Scatena (2005, 2006, 2007). Ellos informaron que en la parte alta del estuario los individuos presentaron un comportamiento reotáctico hasta los 7mm y por lo tanto eran los responsables del reclutamiento y el mantenimiento de la población (Blanco & Scatena 2005). Si asumimos que un comportamiento similar se presenta en la Ecorregión, dándose un reclutamiento de individuos en el borde externo del manglar y un posterior movimiento de juveniles hacia el interior, las poblaciones que presentan fracciones significativas de individuos entre 4 y 7mm han presentado eventos recientes de reclutamiento. Esto se presentó en 18 de 46 estaciones (40% de las estaciones) y solamente en ocho se identificó una cohorte bien definida (datos no mostrados). Por otro lado, de acuerdo a las tallas promedio (y el ámbito) de las poblaciones en la Ecorregión, no parece existir un aislamiento significativo. Inclusive la cohorte modal es consistente en la mayor parte de las estaciones. Unas pocas estaciones presentaron tallas promedio muy grandes (>10mm) con ámbitos estrechos mientras que otras estaciones, tallas promedio muy pequeñas (<6mm) con ámbitos estrechos. Sin embargo, la limitación de larvas no parece ser un factor determinante para la estructura genética de las poblaciones dentro del Golfo, como se discutirá más adelante. Las diferencias entre poblaciones con respecto a la presencia o no de reclutas pueden ser influenciadas por factores post-reclutamiento. Para explicar los ámbitos de las poblaciones y las frecuencias de tallas de ambas especies proponemos dos hipótesis, una biogeográfica y otra ecológica, las cuales se explican a continuación.
Hipótesis biogeográfica. Al comparar la Ecorregión Darién con el resto del Caribe colombiano es evidente que presenta condiciones oceanográficas particulares (bajas salinidad y temperatura) determinadas por la acción de los vientos, las corrientes marinas y el gran caudal del río Atrato (Ruiz & Bernal 2009, Bernal et al. 2006, Ruiz et al. 2010). La baja salinidad (<10PSU) es una barrera geográfica para la dispersión de los moluscos marinos estenohalinos, tales como L. angulifera, hacia el interior del Golfo. Littoraria es un género predominantemente marino (sensu Sanpanich et al. 2008), a pesar que muchas especies están asociadas a estuarios. Las poblaciones establecidas están en el Chocó (al norte del delta del río Atrato) y en el Caribe antioqueño. La única muestra observada al interior del Golfo (bahía Turbo) apoya las preferencias por salinidades >15PSU. Esta población podría estar aislada genéticamente de las establecidas en el exterior de la Ecorregión, y de la fuente de larvas proveniente del Caribe. Gaines et al. (1974) mostraron que la distancia genética (basada en el alelo de esterasa) de L. angulifera entre poblaciones de islas de manglares en la Florida no estuvo relacionada con la distancia entre islas sino con la latitud, aunque la separación máxima fue de 15km. También hubo gran separación genética aun entre poblaciones distanciadas apenas 300m, posiblemente debido a deriva genética aleatoria o a la selección ambiental. De acuerdo a los autores, la dispersión de las larvas véligeras podría estar limitada por las corrientes marinas. Sin embargo, advirtieron que no necesariamente la dispersión implica asentamiento y reclutamiento exitosos debido a factores ambientales o densodependientes. Se ha mencionado que además de la estrategia ovovivípara en L. angulifera, las larvas tienen una corta duración (<10 semanas) (Gallagher & Reid 1979). En otro estudio, Merkt & Ellison (1998), comparando las costas del Atlántico suramericano y africano, encontraron variabilidad fenotípica significativa, lo que posiblemente indica diferencias genotípicas.
N. virginea presentó una amplio ámbito en la Ecorregión Darién y está en los dos únicos manglares conservados de la costa del Chocó y en Damaquiel en el Caribe antioqueño. Esta es una especie de amplio ámbito en el Caribe que se encuentra en praderas de pastos, manglares y puede subir varios kilómetros río arriba. Este amplio ámbito puede ser explicado por la naturaleza eurihalina observada en las formas adulta, juvenil y larval en los Neritidae (Crandall 1999), asociada posiblemente con su ciclo de vida anfídromo. Aunque no existen estudios al respecto, la evidencia indirecta sugiere que la larva es teleplánica, al igual que en otros Neritidae anfídromos (Myers et al. 2000, Cook et al. 2009). Por ello, las poblaciones de N. virginea de la Ecorregión del Darién podrían estar relacionadas con las de otras poblaciones del Caribe panameño presentes en los manglares y en las praderas de pastos, ambos bajo condiciones más marinas. Es posible que la relación sea mayor con esta área que con la del Caribe colombiano sur (entre la CGSM y golfo de Morrosquillo) debido a que la corriente predominante es occidente-oriente (Giro de Panamá-Colombia, Andrade et al. 2003). Además recientemente, se ha identificado que el patrón de vientos predominante norte-sur represa de manera significativa el caudal de agua dulce del río Atrato, al menos durante la época de seca (García 2007). Sin embargo, debido a esto posiblemente, las poblaciones ubicadas al Sur del delta del río Atrato podrían estar aún más aisladas de las que se encuentran en el Caribe. Las de la ensenada de Rionegro también podrían estar aisladas debido a su condición de laguna costera cuya conexión con el mar es intermitente. Trabajos basados en marcadores moleculares han encontrado que Neritidae anfídromos (N. virginea en Puerto Rico; Cook et al. 2009 y Clithon spinosus en la Polinesia Francesa; Myers et al. 2000) no presentaron estructura genética a ninguna escala de observación, es decir dentro de quebradas, entre quebradas dentro de una misma isla o aun entre islas distanciadas no solamente espacial sino temporalmente (diferente edad geológica). Sin embargo, a partir de estos estudios fue posible identificar varios haplotipos dentro de las poblaciones lo que sugiere los distintos orígenes de las larvas y vías predominantes de transporte como las corrientes marinas. En la Polinesia Francesa (Myers et al. 2000) no se encontró estructura genética aun entre islas separadas más de 140km, lo que apoya la hipótesis de una estrategia larval teleplánica (i.e. duración <1 año).
Hipótesis ecológica. La escasez de poblaciones establecidas de L. angulifera y de N. virgínea en el Caribe antioqueño y posiblemente en la mayor parte del Chocó podría ser explicada por el pobre estado de conservación de los manglares. Muchas áreas de manglar han sido taladas o erosionadas por los mares de leva y la deriva litoral y ahora permanecen unos pocos árboles en pie (Correa & Vernette 2004, Urrego et al. 2010). Como se explicó en la sección anterior, el aporte de larvas no parece ser una limitante para la distribución de ambas especies en la parte externa de la Ecorregión. De hecho, L. angulifera se asentó sobre troncos en las bermas de las playas, lo que sugiere una alta tolerancia a la emersión y desecación como ha sido observado en manglares de otras áreas geográficas (Tanaka & Maia 2006, Merk & Ellison 1998) y en otros littorinidos de los manglares del Pacífico colombiano (Blanco & Cantera 1999). En el golfo de Urabá se han observado individuos en las zonas intermareales altas en las playas y los manglares, pero en las bermas de las playas no se observan individuos grandes (JF Blanco, pers. observ.), posiblemente debido a que los troncos pueden ser movidos con facilidad durante los mares de leva (overwash), matando, así, a los reclutas recientes.
Aunque N. virginea es una especie eurihalina, parece ser sensible a la desecación y se encuentra predominantemente en charcos y neumatóforos o en las partes bajas de las raíces y troncos, desde donde se desplaza hacia el suelo cuando la marea empieza a bajar (JF Blanco, pers. observ.). Además, N. virginea es sensible a la alta turbiedad y a las altas concentraciones de fósforo total en los ríos costeros (Blanco & Scatena 2006). Esto podría explicar su ausencia en los manglares talados, contaminados con aguas residuales y con aguas turbias. Algunos caños del Caribe antioqueño drenan humedales que aportan muchos taninos y por lo tanto producen aguas naturalmente negras que podrían evitar la presencia de N. virginea en los manglares de sus desembocaduras. Ambas especies parecen necesitar los substratos duros provistos por los mangles. L. angulifera se observó dentro de los manglares predominantemente sobre troncos por encima de la línea de la marea, mas no en raíces bajas o en el suelo. Fuera de los manglares, se encontró en sustratos duros como troncos caídos en las bermas de las playas, rocas, e incluso sobre estructuras de muelles (i.e. bahía Turbo). En otras áreas geográficas constituye un importante componente de las comunidades de las costas rocosas (Reid 2001). Estas preferencias de sustrato difieren a lo planteado en diferentes estudios en el Caribe Colombiano que lo asocian exclusivamente a las raíces aéreas de Rhizophora mangle y con un amplio ámbito de salinidad (Blanco et al. 2010). Estudios sobre otros littorininos de manglar en Hong Kong y Brasil sugieren marcadas asociaciones con biotopos particulares (Lee & Williams 2002), diferencias entre especies de mangles y una alta fidelidad a los árboles (Tanaka & Maia 2006). Por lo anterior, se hipotetiza que la tala de árboles o su pérdida por erosión tiene efectos muy negativos sobre las poblaciones de L. angulifera ya que puede contribuir directamente a la mortalidad de los adultos establecidos. Finalmente, el impacto de los mares de leva sobre los sedimentos y superficies de los mangles podría ser otra explicación para la escasez de ambas especies dentro de los manglares del Caribe antioqueño y chocoano ya que casi todos ellos se encuentran desprotegidos y muchos muestran signos de erosión natural.
Se confirma así que L. angulifera es una especie de carácter estenohalina y marina, mientras que N. virginea es eurihalina, con predominancia estuarina, por lo tanto sus ámbitos en la Ecorregión Darién están influenciados por el aporte de agua dulce desde el río Atrato (preguntas uno, dos y tres). La salinidad también tiene una influencia sobre el ámbito de tallas de N. virginea en toda la Ecorregión. También se hipotetiza que la ausencia o limitada distribución en algunas localidades afuera del Golfo está explicada por el pobre estado de conservación del manglar (pregunta 3).
Agradecimientos
Agradecemos a la Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional y EAFIT por la financiación del proyecto “Expedición Estuarina, golfo de Urabá, Fase 1” dentro del megaproyecto Expedición Antioquia 2013. Queremos dar gracias al grupo de estudiantes y profesores del proyecto por su apoyo en el campo y en el laboratorio, así como por las discusiones sobre las ideas aquí presentadas. Se agradece especialmente a Edgar Andrés Estrada Urrea por su apoyo con los sistemas de información geográfica y la logística de campo. Los laboratorios de Limnología y Zoología General de la Universidad de Antioquia proveyeron espacio físico y equipos para procesar las muestras. Se agradece a la Corporación para el Desarrollo Sosteniblede la Región de Urabá (CORPOURABA) por proveer información primaria y secundaria. Los concejos comunitarios de Bocas del Atrato y del Chocó Norte-Río Acandí autorizaron el acceso a sus territorios. Contribución ELICE No. 4.
Referencias
Andrade, C.A., E.D. Barton & C.N. Mooers. 2003. Evidence for an eastward flow along the Central and South American Caribbean Coast. J. Geophys. Res. 108: 3185-3196. [ Links ]
Bernal, G., G. Poveda, P. Roldán & C. Andrade 2006. Patrones de variabilidad de las temperaturas superficiales del mar en la Costa Caribe Colombiana. Rev. Acad. Colomb. Cienc. 30: 195-208. [ Links ]
Blanco, J.F. & J.R. Cantera. 1999. The vertical distribution of mangrove gastropods and environmental factors relative to tide level at Buenaventura Bay, Pacific Coast of Colombia. Bull. Mar. Sci. 65: 617-630. [ Links ]
Blanco, J.F. & F.N. Scatena. 2005. Floods, habitat hydraulics and upstream migration of Neritina virginea (Gastropoda: Neritidae) in northeastern Puerto Rico. Caribb. J. Sci. 41: 55-74. [ Links ]
Blanco, J.F. & F.N. Scatena. 2006. Hierarchical contribution of river-ocean connectivity, water chemistry, hydraulics, and substrate to the distribution of diadromus snails in Puerto Rican streams. J. North Amer. Benthol. Soc. 25: 82-98. [ Links ]
Blanco, J.F. & F.N. Scatena. 2007. The spatial arrangement of Neritina virginea (Gastropoda: Neritidae) during upstream migration in a Split-channel reach. River Res. Appl. 23: 235-245. [ Links ]
Blanco, J.F. & A. Arroyave. 2009. Daños por depredación y tamaño de concha del caracol diádromo Neritina virginea (Gastropoda: Neritidae) en el Río Mameyes, Puerto Rico. Rev. Biol. Trop. 57: 1069-1080. [ Links ]
Blanco, J.F., M. Londoño-Mesa, L. Quan-Young, L. Urrego- Giraldo, J. Polanía, A. Osorio, G. Bernal & I. Correa 2011. The Urabá Gulf Mangrove Expedition of Colombia. ISME/GLOMIS 9: 8-10. [ Links ]
Cook, B.D., C.M. Pringle & J.M. Hughes. 2009. Marine dispersal determines the genetic population structure of migratory stream fauna of Puerto Rico: evidence for island-scale population recovery processes. J. North Amer. Benthol. Soc. 28: 709-718. [ Links ]
Correa, I.D. & G. Vernette. 2004. Introducción al problema de la erosión litoral en Urabá (sector Arbolete- Turbo) Costa Caribe colombiana. Bol. Invest. Mar. Cost. 33: 7-28. [ Links ]
Crandall, E.D. 1999. Early life history aspects of amphidromous neritid snails in Moorea, French Polynesia. Berkeley Sci. 2: 98-103 [ Links ]
Díaz, J.M. 1994. La malacofauna de la zona costera de Santa Marta y el Parque Nacional Natural Tayrona, Caribe Colombiano. An. Inst. Inv. Mar. Punta Betín 23: 15-43. [ Links ]
Díaz, J.M. 1995. Zoogeography of Marine Gastropod in the Southern Caribbean: A New Look at Provinciality. Caribb. J. Sci. 31: 104-121. [ Links ]
Ellison, A.M. 2008. Managing mangroves with benthic biodiversity in mind: Moving beyond roving banditry. J. Sea Res. 59: 2-15. [ Links ]
Gaines, M.S., J. Caldwell & A.M. Vivas. 1974. Genetic variation in the mangrove periwinkle Littorina angulifera. Mar. Biol. 27: 327-332. [ Links ]
Gallagher, S.B. & G.K. Reid. 1979. Population dynamics and zonation of the periwinkle Littoraria angulifera of the Tampa Bay, Florida, region. Nautilus 94:162-178. [ Links ]
García, J.M. & J. Palacio. 2008. Macroinvertebrados asociados a las raíces sumergidas del mangle rojo (Rhizophora mangle) en las bahías Turbo y El Uno, Golfo de Urabá (Caribe Colombiano). Ges. Amb. 11: 55-66. [ Links ]
García, C. (ed.). 2007. Atlas del golfo de Urabá: una mirada al Caribe de Antioquia y Chocó. Instituto de Investigaciones Marinas y Costeras (Invemar) y Gobernación de Antioquia. Serie de Publicaciones Especiales de Invemar Nº 12. Santa Marta, Colombia. [ Links ]
Guisande, C., A. Berrio, I. Maneiro, I. Riveiro, A. Vergara & A. Vaamonde. 2006. Tratamiento de datos. Díaz de Santos, Madrid, España. [ Links ]
Hogarth, P.J. 2007. The biology of mangroves and seagrasses. Oxford University, New York, EEUU. [ Links ]
Kyle, C.J. & E.G. Boulding. 2000. Comparative population genetic structure of marine gastropods (Littorina spp.) with and without pelagic larval dispersal. Mar. Biol. 137: 835-845. [ Links ]
Lee, S.Y. 2008. Mangrove macrobenthos: Assemblages, services, and linkages. J. Sea Res. 59: 16-29. [ Links ]
Llano, R. & J. Palacio. 1988. Ecología de la fauna asociada a raíces de mangle Rhizophora mangle en la Bahía de Marirrío Urabá. Tesis de pregrado, Instituto de Biología, Universidad de Antioquia, Medellín, Colombia. [ Links ]
Merkt, R.E. & A.M. Ellison. 1998. Geographic and habitatspecific morphological variation of Littoraria (Littorinopsis) angulifera (Lamarck, 1822). Malacologia 40: 279-295. [ Links ]
Myers, M.J., C.P. Meyer & V.H. Resh. 2000. Neritid and thiarid gastropods from French Polynesian streams: how reproduction (sexual, parthenogenetic) and dispersal (active, passive) effect population structure. Fresh Wat. Biol. 44: 535-545. [ Links ]
Ortiz, L.F. & J.F. Blanco. 2010. Moluscos (gasterópodos y bivalvos) asociados a las raíces sumergidas en los manglares del golfo de Urabá: patrones espaciales y determinantes a macroescala. Proyecto Expedición Estuarina, golfo de Urabá, fase 1. Informe final. Julio 2010. Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional Sede Medellín, Universidad EAFIT, Medellín, Colombia. [ Links ]
Osorio, A.F., L.G. Molina & J.D. Osorio. 2010. Caracterización hidrodinámica del oleaje local (sea) y de fondo (swell) en el golfo de Urabá. Proyecto Expedición Estuarina, golfo de Urabá, fase 1. Informe final. Julio 2010. Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional Sede Medellín, Universidad EAFIT, Medellín, Colombia. [ Links ]
Palacio, J. 1978. Variación de la fauna de invertebrados del área estuárica de la Ciénaga Grande de Santa Marta en relación con los cambios de salinidad. An. Inst. Inv. Mar. Punta Betín 10: 111-126. [ Links ]
Palacio, J. 1983. Die benthische Makroinvertebraten Fauna der Tropischen Ästuarregion Ciénaga Grande de Santa Marta (Kolumbien) und ihre aktivität im Wschsel zwischen Trockenund Regenzeit. Dissertation Zur erlangune des Grandes eines Doktors der Natur wissenschanften an der Ruhr, Universitat Bochum, Alemania. [ Links ]
Ramírez, A. 2005. Ecología Aplicada, Diseño y Análisis Estadístico. Universidad de Bogotá Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ]
Reid, D.G. 2001. New data on the taxonomy and distribution of the genus Littoraria Griffith & Pidgeon, 1834 (Gastropoda: Littorinidae) in Indo-West Pacific mangrove forest. Nautilus 115: 115-139. In K. Sanpanich, F.E. Wells & Y. Chitramvong. 2008. Reproduction and growth of Littoraria (Gatropoda: Littorinidae) at AngSila, Thailand. Raffles Bull. Zool. 18: 225-233. [ Links ]
Reyes, R. & N. Campos. 1992a. Macroinvertebrados colonizadores de raíces de Rhizophora mangle en la Bahía de Chengue, Caribe Colombiano. An. Inst. Inv. Mar. Punta Betín 21: 101-116. [ Links ]
Reyes, R. & N. Campos. 1992b. Moluscos, Anélidos y Crustáceos asociados a las raíces de Rhizophora mangle Linnaeus, en la región de Santa Marta, Caribe Colombiano. Caldasia 17: 133-148. [ Links ]
Riascos, J.M. & P.A. Guzmán. 2010. The ecological significance of growth rate, sexual dimorphism and size at maturity of Littoraria zebra and L. variegate (Gastropoda: Littorinidae). J. Mollusc. Stud. 76: 289-295. [ Links ]
Rojas, J. & J. Palacio. 2003. Contribución al conocimiento de la fauna de macroinvertebrados asociados a las raíces del manglar en el Golfo de Urabá. Trabajo de grado en Biología, Universidad de Antioquia, Medellín, Colombia. [ Links ]
Ruiz, M. & G. Bernal 2009. Variabilidad estacional e interanual del viento en los datos del reanálisis NCEP/ NCAR en la cuenca Colombia, mar Caribe. Avan. Rec. Hidraul. 20: 7-20. [ Links ]
Ruiz, M., G. Bernal, E. Beier & A. Osorio. 2010. La salinidad en la cuenca Colombia, mar Caribe, p. 39-43. In INVEMAR-ACIMAR (eds.). Libro de resúmenes extendidos XIV Seminario Nacional de Ciencia y Tecnología del Mar (SENALMAR). Serie de Publicaciones Especiales de Invemar No. 21. Santa Marta, Colombia. [ Links ]
Sanpanich, K., F.E. Wells & Y. Chitramvong. 2008. Reproduction and growth of Littoraria (Gatropoda: Littorinidae) at Ang Sila, Thailand. Raffles Bull. Zool. 18: 225-233. [ Links ]
Tanaka, M.O. & R.C. Maia. 2006. Shell morphological variation of Littoraria angulifera among and within mangroves in NE Brazil. Hydrobiology 559: 193-202. [ Links ]
Urrego, L.E., E.C. Molina, J.A. Suárez, H. Ruiz & J. Polanía. 2010. Distribución, composición y estructura de los manglares del golfo de Urabá. Proyecto Expedición Estuarina, golfo de Urabá, fase 1. Informe final. Julio 2010. Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional Sede Medellín, Universidad EAFIT, Colombia. [ Links ]
Vilardy, S. & J. Polanía. 2002. Mollusc fauna of the mangrove root-fouling community at the Colombian Archipelago of San Andrés and Old Providence. Wetl. Ecol. Manag. 10: 273-282. [ Links ]
Vermeij, G.J. 1971. Mollusk in mangrove swamps: Physiognomy, diversity and regional differences. Syst. Zool. 22: 609-624. [ Links ]
*Correspondencia a: Luis Ferney Ortiz. Grupo de Ecología Lótica: Islas, Costas y Estuarios (ELICE), Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Calle 67 Número 53-108, Medellín, Colombia. Apartado Aéreo 1226; lfortiz24@gmail.com
Juan Felipe Blanco. Grupo de Ecología Lótica: Islas, Costas y Estuarios (ELICE), Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Calle 67 Número 53-108, Medellín, Colombia. Apartado Aéreo 1226; blanco@exactas.udea.edu.co
1. Grupo de Ecología Lótica: Islas, Costas y Estuarios (ELICE), Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Calle 67 Número 53-108, Medellín, Colombia. Apartado Aéreo 1226; lfortiz24@gmail.com, blanco@exactas.udea.edu.co
Recibido 11-II-2011. Corregido 12-VII-2011. Aceptado 18-VIII-2011.
En el Caribe Colombiano, la malacofauna asociada al manglar ha sido ampliamente estudiada, siendo la ciénaga Grande de Santa Marta y su vecindad una de las áreas más exploradas desde los años setentas (Cosel 1986, Díaz 1994, 1995, Palacio 1978, 1983). La bahía de Cartagena y el golfo de Morrosquillo también han sido históricamente estudiados (revisado por Ortiz & Blanco 2010, Blanco et al. 2010), mientras que el Archipiélago de San Andrés y Providencia ha recibido atención durante la última década (Vilardy & Polanía 2002). Sin embargo, la Ecorregión Darién (Caribe colombiano sur) ha sido poco explorada. Dentro de ésta, se han realizado trabajos sobre la dinámica de la comunidad de macroinvertebrados epibiontes de las raíces en áreas geográficas puntuales del golfo de Urabá (bahía Marirrío: Llano 1998, bahías El Uno y Turbo: García & Palacio 2008). Los dos únicos estudios que han abarcado una porción significativa del golfo de Urabá son el de Rojas & Palacio (2003) y el de Ortiz & Blanco (2010). Sin embargo, el estudio de Ortiz & Blanco (2010) es hasta ahora el de mayor extensión (49 estaciones a lo largo de 609km de costa) y mayor esfuerzo de muestreo (245 raíces, 69 932 individuos). Este estudio, enfocado en los moluscos epibénticos de las raíces de los mangles, confirmó que esta taxocenosis es pobre (ocho especies) dentro de la Ecorregión Darién en comparación al resto del Caribe colombiano y al Gran Caribe, con una dominancia del 94% por parte de dos bivalvos (Mytilopsis cf. sallei y Brachidontes cf. exustus). Este estudio también evidenció que la mayor parte de las especies eran eurihalinas posiblemente debido a la predominancia de condiciones estuarinas en el Golfo la mayor parte del año.
Debido a que la Ecorregión Darién es: a) un área poco conocida en la literatura internacional, b) alberga un área significativa de manglar (6 512ha, García 2007), y c) recibe uno de los aportes de agua dulce más grandes de todo el Caribe sur a través del río Atrato, se decidió explorar los patrones de arreglo espacial a la mesoescala (105m) de Littoraria (Littorinopsis) angulifera (Lamarck, 1822) y Neritina (Vitta) virginea (L.), dos gasterópodos abundantes en los suelos y raíces emergidos de los manglares de la Ecorregión Darién, al igual que en el Gran Caribe (Palacios 1978, Díaz 1994). Adicionalmente, los manglares de la Ecorregión Darién han sido objeto de múltiples presiones a través de los años, por lo cual el reciente reconocimiento de su gran valor ecosistémico y de las múltiples funciones de sus componentes, han llevado a tomar acciones para un manejo sustentable y su conservación (García 2007). Específicamente, se formularon las siguientes preguntas: 1) ¿Cuál es el ámbito de distribución de las poblaciones de L. angulifera y N. virginea en el área de estudio? 2) ¿Cómo varía la estructura de tallas de las poblaciones de ambos gasterópodos? 3) ¿Existe relación entre dichas variables con la variación espacial de la salinidad y el estado de conservación de los manglares?
Materiales y métodos
Especies estudiadas. El gasterópodo N. virginea se encuentra desde Florida y Bahamas hasta el norte de Brasil, y desde el Golfo de México y América Central hasta Las Antillas. El tamaño oscila entre 4 y 25mm, desde los individuos recientemente reclutados hasta los adultos más grandes. Los juveniles y adultos exhiben una coloración verde con un patrón variable de líneas axiales o de pequeños triángulos amarillos. Coloniza las praderas de fanerógamas y los manglares de lagunas costeras, bahías y golfos, pero también se encuentran densas poblaciones desde la desembocadura de los ríos hasta varios kilómetros aguas arriba. También coloniza pequeños ríos y quebradas costeras, presentando un ciclo de vida diádromo. En este ciclo la larva recién eclosionada del huevo es arrastrada aguas abajo hasta el mar. La larva es planctotrófica y se cree que puede durar varios meses a la deriva transportada por las corrientes marinas (Crandall 1999, Blanco & Scatena 2005, 2006, 2007). Al llegar a las costas cercanas a los estuarios, las larvas se transforman en reclutas bénticos que empiezan a migrar río arriba, proceso que dura toda su vida. Al alcanzar la madurez sexual ponen cápsulas ovígeras con cientos de huevos. Información detallada sobre la ecología de la especie en quebradas costeras del Caribe ha sido publicada por Blanco & Scatena (2005, 2006, 2007) y Blanco & Arroyave (2009). En los manglares los individuos hacen migraciones horizontales y verticales dependiendo de los ciclos mareales diarios y mensuales, y se encuentran sobre el fango, hojarasca, neumatóforos y las partes bajas de las raíces aéreas y troncos (J.F. Blanco, pers. observ.).
L. angulifera habita en las zonas supralitorales de los manglares del Atlántico y el Caribe (Tanaka & Maia 2006). Se encuentra principalmente en hojas, troncos y raíces adventicias de los mangles (Rhizophora mangle L., Avicennia germinans, Avicennia marina, Avicennia alba, Avicennia nitida y Laguncularia racemosa) donde se alimentan de algas y hongos marinos (Merk & Ellison 1998). Este gasterópodo es ovivíparo incompleto, con desarrollo larval en la cavidad corporal de la hembra hasta el estado velígero, el cual es liberado en la columna de agua (Gaines et al. 1974). El estado larval planctotrófico tiene un periodo de duración aproximado de 8-10 semanas (Merk & Ellison 1998).
Área de estudio. La Ecorregión Darién en el Caribe comprende el sur de Panamá y Colombia. En el extremo suroccidental del Caribe colombiano, la zona costera está comprendida por los departamentos de Chocó y Antioquia, donde sobresalen las subregiones Caribe chocoano, golfo de Urabá y Caribe antioqueño. El golfo de Urabá (7º50’ - 8º56’ N y longitud: 77º22’ - 76º25’ W) es la más extensa, siendo un cuerpo de agua semicerrado, alargado en dirección norte-sur con una morfología similar a la letra U (aproximadamente 80km de largo y 25km de ancho promedio, con cerca de 8km en su parte más estrecha). Tiene un área de 4 291km2, y es la entrada más grande del mar Caribe en tierra colombiana (García 2007). El Golfo es el estuario de mayor tamaño en el Caribe colombiano ya que en la parte media de su costa occidental desemboca el río Atrato (4 155m3/s), el de segundo mayor caudal del Caribe colombiano (después del río Magdalena), y que exhibe un extenso delta digitado. En el costado suroriental desembocan varios ríos de menor caudal (Turbo, Guadualito, Currulao y León) que forman microdeltas acuminados moldeados por el oleaje.
La estacionalidad de los vientos y de la precipitación (época húmeda y época seca) juegan un papel importante en la distribución espacial de la salinidad. La circulación de los vientos en el área está en fase con el movimiento de la Zona de Convergencia Inter-Tropical cuyo desplazamiento hacia el sur de Colombia en la época seca (diciembre-enero) trae consigo un aumento de los vientos Alisios del noreste, mientras que cuando ésta se desplaza hacia el norte en la época húmeda, se presentan los vientos Alisios del sureste. En la época seca, aunque las precipitaciones son muy bajas, el aumento de los vientos Alisios del norte y nordeste retienen el aporte de agua continental ocasionando que el Golfo se comporte como una enorme “piscina” de agua oligo- a mesohalina a nivel superficial. En la época húmeda los vientos son débiles y se caracterizan por soplar principalmente del sur y suroeste. Además, se presentan fuertes precipitaciones, por lo cual el aporte de agua dulce proveniente delrío Atrato y de otros ríos de menor magnitud es exportado del Golfo, y permanece una masa de agua más salada (polihalina) al interior (bahía Colombia) (García 2007, Osorio et al. 2009).
El delta del río Atrato es el área de mayor extensión y desarrollo de manglar dominada por Rhizophora mangle, con predominio de 80% (densidad relativa), seguida de Laguncularia racemosa. En bahía Colombia se presenta un manglar de tipo ribereño con mezcla de varias especies (río Suriqui). En la costa occidental del Golfo existen pequeñas áreas de manglar de franja, con Rhizophora mangle ubicada en el borde de la costa y Avicennia germinans al interior. La ensenada de Rionegro es una laguna costera ubicada en el límite norte del Golfo, es la mayor área de manglar de la costa oriental, dominada por Rhizophora mangle y Laguncularia racemosa. En el costado suroriental sobresalen los parches de manglar en la desembocaduras de los ríos León y Currulao y en las bahías Turbo y El Uno. Finalmente, en la costa Caribe antioqueña hay unos relictos (caño La Marimonda, río Damaquiel, río Hobo y río Volcán) producto de la actividad antropogénica y la erosión costera (CORPOURABÁ 2003). Para una descripción más detallada de la Ecorregión fue realizada una reciente expedición (Blanco et al. 2011).
Muestreo. Se realizaron dos salidas de campo (3-14 de junio y 3-10 de agosto de 2009) para verificar la existencia de poblaciones de N. virginea y L. angulifera y construir mapas de ámbito de distribución recolectaron entre 50 y 200 individuos al azar de cada especie sobre el suelo, raíces o troncos a lo largo de transectos de 50m dentro de cada sitio de muestreo. Durante 20 minutos se recolectaron entre 50 y 200 individuos al azar de cada especie sobre el suelo, raíces o troncos a lo largo de transectos de 50m dentro de cada sitio de muestreo. Para preservar los individuos recolectados se fijaron en una solución de formol al 10%. En la primera salida de campo se visitaron 39 áreas geográficas en búsqueda de zonas de manglar en los departamentos de Chocó y Antioquia entre cabo Tiburón en el límite con Panamá (al occidente) y quebrada Peñoncito en el límite con el departamento de Córdoba (al oriente). Estas áreas de manglar habían sido identificadas previamente mediante el uso de mapas, imágenes satelitales, aerofotografías y conceptos de expertos. En la segunda salida, se enfocó el muestreo en 32 estaciones dentro del delta del río Atrato (la mayor área de manglar) y dentro de las bahías El Uno y Turbo (Fig. 1).En las desembocaduras de algunas quebradas donde los manglares habían desaparecido, se buscaron y recolectaron individuos de L. angulifera sobre troncos en las bermas de las playas.
En el laboratorio, se midieron las conchas de todos los individuos. A L. angulifera se le midió la longitud axial y a N. virginea se le midió el ancho de la concha sobre la columela utilizando un pie de rey con una precisión de 0.01mm.
En el campo se midió la salinidad o la conductividad dependiendo de la disponibilidad de instrumentos. En los casos que no se obtuvo valor de campo, se estimó utilizando los valores promedios reportados para dicha época climática en el “Atlas del golfo de Urabá” (García 2007). El ámbito de salinidad medido en el campo fue comparado con el reportado en la literatura para validar la interpolación de los datos faltantes.
Los valores georeferenciados de salinidad, áreas de manglar y poblaciones de gasterópodos se proyectaron en el sistema de información geográfica ArcGIS versión 9.3. Los polígonos con las áreas de cobertura de manglares se obtuvieron del inventario más reciente, el cual utilizó ortofotografías a escala 1:10 000 (Urrego et al. 2010, Blanco et al. 2011). Para contestar la pregunta uno, con los datos de frecuencia de las estaciones donde estaba presente cada especie se realizó una tabla de contingencia y una prueba Chi cuadrado para determinar el ámbito de cada especie y si ellas estaban asociadas (Guisande et al. 2006, Ramírez 2005), mediante el uso del programa Statistica 6.0 (edición 98). Para contestar la pregunta dos, se realizó una prueba de Kolmogorov-Smirnof para establecer si las muestras de tallas presentaban distribución normal (Guisande et al. 2006, Ramírez 2005). Las tallas promedio se compararon entre estaciones de muestreo utilizando un Análisis de Varianza (ANOVA) de una vía (Guisande et al. 2006, Ramírez 2005). Los datos de cada salida se trataron en conjunto debido a que las condiciones climáticas y oceanográficas fueron similares. Se realizaron regresiones lineales (Guisande et al. 2006, Ramírez 2005) entre las tallas promedio y la salinidad (pregunta 3). Finalmente, se realizó una prueba t-student para comparar las tallas promedio de L. angulifera entre áreas con o sin manglar dentro del Caribe chocoano y antioqueño (Guisande et al. 2006, Ramírez 2005) (pregunta tres). Esta comparación no se realizó en N. virginea porque no se encontró afuera de los manglares de la Ecorregión.
Resultados
Ámbito de salinidad y áreas de manglar. La salinidad superficial observada osciló entre 0.1 y 37PSU. La parte interna del Golfo (delta del río Atrato) presentó aguas superficiales oligo- y meso-halinas con excepción de la bahía Turbo que presentó los valores más altos de salinidad (polihalina). La salinidad aumentó hacia el Norte como resultado de la mezcla de agua de escorrentia continental con el agua marina, arrojando como resultado aguas polihalinas (18-30PSU) y en algunos casos mixoeuhalinas (30-40PSU). Por lo tanto, las aguas costeras del Golfo presentaron un fuerte gradiente latitudinal de salinidad (Fig. 2).
Los manglares de la Ecorregión Darién cubren actualmente una extensión de 4 912ha. En la parte norte se presentan pequeños relictos de manglar con una cobertura de 33ha. En el Chocó (Titumate y Pinorroa) y el Caribe antioqueño (ríos Damaquiel y Hobo) se encuentran 16 y 17ha, respectivamente, que se ubican dentro de bahías y bocas de los caños y quebradas que los protegen de la erosión. A lo largo de estas subregiones se observaron árboles aislados y algunos talados, indicios de coberturas previas mayores que fueron taladas o erosionadas en algunas localidades. En el departamento de Antioquia, 3 846ha se encuentran en la costa occidental en el delta de río Atrato. La costa oriental presenta un total de 1 034ha, de las cuales 38ha se encuentran en el río Suriqui, 192ha en el río León y punta Coquito, 214ha en los ríos Guadualito y Currulao, 145ha en punta Yarumal y bahía Turbo, 103ha en Caimán Nuevo y 342ha en la ensenada de Rionegro (Fig. 2).
Ámbito espacial de las poblaciones de L. angulifera y N. virginea. Durante la fase de campo (77 estaciones de muestreo) se recolectaron un total de 3 963 individuos de ambas especies: 1 051 de L. angulifera y 2 912 de N. virginea.
El porcentaje de aparición de las poblaciones de cada especie varió considerablemente en el interior y exterior de la Ecorregión Darién. L. angulifera apareció en el 84.8% de las estaciones de la parte norte, mientras que N. virginea apareció solo en un 15.2% de las estaciones de esta parte, particularmente en los manglares protegidos del Chocó (Playa Soledad y Titumate) y la ensenada de Rionegro, el caño de la Marimonda y el río Damaquiel en el Caribe antioqueño. L. angulifera fue la especie dominante del exterior del Golfo (Fig. 3a). Al interior, es decir, en el golfo de Urabá, al sur de punta Yerbasal y Caimán Nuevo, se invirtieron los patrones. L. angulifera apareció en un 15.4% de las estaciones, pero estuvo presente sólo en bahía Turbo, mientras que N. virginea apareció en el 84.6% de las estaciones, en donde las poblaciones establecidas sólo se observaron en los manglares protegidos del oleaje y bien desarrollados de la parte interna(Fig. 3b). Esta especie fue numéricamente dominante en la parte interna.
La prueba Chi2 sobre una tabla de contingencia mostró que no había una asociación significativa entre las especies (p<0.001), por lo que resaltan notables diferencias del ámbito de las especies entre el interior y el exterior de la Ecorregión, a pesar de ser simpátricas en la meso-escala (105).
Ámbito de las tallas de L. angulifera y N. virginea. La muestra de tallas de L. angulifera para la Ecorregión Darién presentó una distribución normal (Prueba de Shapiro-Wilk, p>0.05), mientras que la muestra de tallas de N. virginea no (p<0.001). El ámbito de tallas de L. angulifera, presentó valores promedios entre 6 y 22mm. Este ámbito de tallas promedio de las poblaciones presentó diferencias significativas entre las estaciones de muestreo (F12,1037=61.45, p<0.001). Los individuos más grandes (promedio=21mm) se recolectaron en Titumate y se consideró como una población establecida. En playa Soledad y el río San Juan se observaron tallas promedio alrededor de 15mm, también se consideraron poblaciones establecidas. Sin embargo, estas dos poblaciones presentaron una proporción significativa de individuos <15mm. El resto de estaciones de muestreo presentaron promedios de talla <12mm. En la quebrada La Cartagenita, Pinorroa y caño La Marimonda se observaron los individuos de menor tamaño (≤5mm) (Fig. 4).
Las tallas de N. virginea presentaron una distribución leptocúrtica, con una mayor frecuencia de individuos entre 6 y 12mm (tallas promedio) en la Ecorregión. Se observaron, sin embargo, diferencias significativas entre áreas geográficas (F44, 2868=30.01, p<0.001) pero sin un patrón definido, aunque en el manglar protegido de la bahía El Uno presentó un valor de talla promedio de 12mm y en el manglar de bahía Cocogrande se encontraron las menores tallas promedio (<6mm) (Fig. 4).
Las tallas promedio de L. angulifera en la Ecorregión Darién no exhibieron una correlación significativa (r=0.02, p=0.96) con la salinidad, debido que se encontraron en una zona de alta salinidad y poca variabilidad. Las tallas promedio de N. virginea presentaron una correlación significativa con la salinidad dentro de la Ecorregión (r=0.38, p=0.04), pero no en la parte interna del Golfo (mayor resolución espacial: delta del río Atrato y bahías Turbo y El Uno) (r=0.16, p=0.52).
L. angulifera se encontró ampliamente distribuido dentro y fuera de los manglares de la costa norte, pero las tallas promedio fueron significativamente mayores en manglares establecidos (14±6mm) en el Caribe chocoano y antioqueño y no afuera de ellos (10±3.2mm) (t=4.48, g.l.=8, p=0.01). N. virginea presentó un ámbito limitado a las zonas de manglar, las mayores tallas y abundancias promedio se encontraron en el interior del Golfo.
Discusión
Condicionantes del ámbito de salinidad en la Ecorregión Darién. El continuo aporte de agua dulce del río Atrato al Golfo, confiere condiciones estuarinas durante todo el año, que lo divide en dos sectores (García 2007). El sector norte registra los valores más altos de salinidad, se clasifica como una zona de aguas polihalinas (18-30PSU) y mixoeuhalinas (30-40PSU), resultado de la influencia directa del agua marina y el poco aporte de agua dulce por los ríos de este sector. Los ríos más grandes son Mulatos, San Juan y Damaquiel pero sus caudales no están aforados. La ensenada de Rionegro, por su condición de laguna costera, puede presentar estacionalmente valores altos de salinidad. Rojas & Palacio (2003) informaron que en esta ensenada se midió el valor más alto de las 24 estaciones del litoral antioqueño. El sector sur del Golfo presenta salinidades bajas (0.1-7.9PSU), dado el gran y continuo aporte de agua dulce del río Atrato y otros ríos de menor caudal (ríos Suriquí, León, Currulao, Guadualito, Turbo, Caimán Nuevo y Caimán Viejo), y de corrientes menores (quebradas y caños) (García 2007). Al interior del Golfo, la bahía Turbo presenta valores de salinidad relativamente altos, como resultado del confinamiento de las aguas por la disposición latitudinalmente alargada de la bahía y la dirección del viento local predominantemente sur-norte. Otra posible causa de estas condiciones es un largo periodo de residencia y poca mezcla de sus aguas y la alta presión antrópica a la que está expuesta (puerto y asentamiento humano). Ramírez (2005) plantea que la actividad antrópica en las zonas costeras puede alterar las características fisicoquímicas de las aguas por medio de la contaminación puntual y difusa. Adicionalmente, el poco aporte de agua dulce que recibe y la alta evaporación podrían mantener una masa de agua de mayor salinidad al interior de la bahía, como sucede en otros estuarios del mundo.
Los valores de salinidad superficial observados durante nuestro estudio, fueron característicos de una época seca (junio y agosto). Estos meses normalmente corresponden a un periodo de transición, veranillo de mitad de año (García 2007), con predominancia de vientos débiles con dirección sur y suroeste, que convirten al golfo en una enorme piscina de agua dulce (García 2007).
La salinidad y el ámbito de los gasterópodos. La salinidad es la principal determinante de la distribución de la fauna asociada al manglar (Rojas & Palacio 2003, García & Palacio 2008) y a los estuarios (Vermeij 1971). Por ejemplo, en la ciénaga Grande de Santa Marta (Palacio 1978), la bahía Chengue (Reyes & Campos 1992 a, b), la bahía de Cartagena y la ciénaga Los Vásquez (Pérez & Victoria 1977) en el Caribe colombiano los cambios en la distribución de especies obedecen a cambios en la salinidad. Nuestros resultados apoyan parcialmente lo observado en otras áreas del Caribe colombiano, a pesar de los amplios ámbitos de las especies estudiadas. L. angulifera se encontró restringida a salinidades >15PSU en la parte norte (y disyuntamente en la bahía Turbo), pero N. virginea se encontró en salinidades entre 0 y 37PSU, y muestra un mayor ámbito. Sin embargo la variación de las tallas si es influenciada por la salinidad a la meso escala. La limitada presencia de esta segunda especie en el Caribe chocoano y antioqueño obedece aparentemente a otros factores como se explicará más adelante.
Ámbito de las tallas. Las tallas promedio de L. angulifera no mostraron cambios pronunciados con respecto a otros Littorinidos del Pacífico y el Atlántico (Tailandia, el Pacífico colombiano y Brasil), en donde los adultos oscilan entre 16 y 22mm (Sanpanich et al. 2008), los juveniles entre 9 y 16mm, y los reclutas 6 y 9mm (Tailandia: Sanpanich et al. 2008; Brasil: Tanaka & Maia 2006; Pacífico Colombiano: Blanco & Cantera 1999 y Riascos & Guzmán 2010). En el presente estudio, las tallas promedio de L. angulifera sugieren que hay pocas localidades con poblaciones compuestas por adultos y juveniles, pero la mayoría de las localidades presentan tallas promedio que indican la prevalencia de juveniles y reclutas y la ausencia de adultos. Esto podría ser un indicador de la estabilidad de la población asociada con el estado de conservación del manglar, como se discutirá más adelante.
De acuerdo al ámbito de tallas promedio de las poblaciones es posible que algunas estén relativamente aisladas. Las poblaciones de playa Soledad y Titumate (Caribe chocoano) fueron las de mayor talla promedio y las únicas que se encontraron dentro de manglares relativamente extensos, y mantienen, al menos parcialmente, el aporte local de larvas. Por el contrario, las poblaciones de individuos de tallas mucho menores que se encontraron sobre las bermas de las playas del Caribe chocoano y antioqueño, dependen del aporte extrínseco de larvas, las cuales pueden ser transportadas desde las localidades próximas hacia éstas, por el oleaje y corrientes del Caribe, el cual está dominado fuertemente en este sector por el oleaje tipo Swell (comportamiento independiente entre el oleaje y la magnitud de los vientos) (Osorio et al. 2010).
La talla promedio de N. virginea presentó una correlación significativa con la salinidad en la Ecorregión Darién. Las tallas observadas se encontraron dentro del ámbito observado en Puerto Rico por Blanco & Scatena (2005, 2006, 2007). Ellos informaron que en la parte alta del estuario los individuos presentaron un comportamiento reotáctico hasta los 7mm y por lo tanto eran los responsables del reclutamiento y el mantenimiento de la población (Blanco & Scatena 2005). Si asumimos que un comportamiento similar se presenta en la Ecorregión, dándose un reclutamiento de individuos en el borde externo del manglar y un posterior movimiento de juveniles hacia el interior, las poblaciones que presentan fracciones significativas de individuos entre 4 y 7mm han presentado eventos recientes de reclutamiento. Esto se presentó en 18 de 46 estaciones (40% de las estaciones) y solamente en ocho se identificó una cohorte bien definida (datos no mostrados). Por otro lado, de acuerdo a las tallas promedio (y el ámbito) de las poblaciones en la Ecorregión, no parece existir un aislamiento significativo. Inclusive la cohorte modal es consistente en la mayor parte de las estaciones. Unas pocas estaciones presentaron tallas promedio muy grandes (>10mm) con ámbitos estrechos mientras que otras estaciones, tallas promedio muy pequeñas (<6mm) con ámbitos estrechos. Sin embargo, la limitación de larvas no parece ser un factor determinante para la estructura genética de las poblaciones dentro del Golfo, como se discutirá más adelante. Las diferencias entre poblaciones con respecto a la presencia o no de reclutas pueden ser influenciadas por factores post-reclutamiento. Para explicar los ámbitos de las poblaciones y las frecuencias de tallas de ambas especies proponemos dos hipótesis, una biogeográfica y otra ecológica, las cuales se explican a continuación.
Hipótesis biogeográfica. Al comparar la Ecorregión Darién con el resto del Caribe colombiano es evidente que presenta condiciones oceanográficas particulares (bajas salinidad y temperatura) determinadas por la acción de los vientos, las corrientes marinas y el gran caudal del río Atrato (Ruiz & Bernal 2009, Bernal et al. 2006, Ruiz et al. 2010). La baja salinidad (<10PSU) es una barrera geográfica para la dispersión de los moluscos marinos estenohalinos, tales como L. angulifera, hacia el interior del Golfo. Littoraria es un género predominantemente marino (sensu Sanpanich et al. 2008), a pesar que muchas especies están asociadas a estuarios. Las poblaciones establecidas están en el Chocó (al norte del delta del río Atrato) y en el Caribe antioqueño. La única muestra observada al interior del Golfo (bahía Turbo) apoya las preferencias por salinidades >15PSU. Esta población podría estar aislada genéticamente de las establecidas en el exterior de la Ecorregión, y de la fuente de larvas proveniente del Caribe. Gaines et al. (1974) mostraron que la distancia genética (basada en el alelo de esterasa) de L. angulifera entre poblaciones de islas de manglares en la Florida no estuvo relacionada con la distancia entre islas sino con la latitud, aunque la separación máxima fue de 15km. También hubo gran separación genética aun entre poblaciones distanciadas apenas 300m, posiblemente debido a deriva genética aleatoria o a la selección ambiental. De acuerdo a los autores, la dispersión de las larvas véligeras podría estar limitada por las corrientes marinas. Sin embargo, advirtieron que no necesariamente la dispersión implica asentamiento y reclutamiento exitosos debido a factores ambientales o densodependientes. Se ha mencionado que además de la estrategia ovovivípara en L. angulifera, las larvas tienen una corta duración (<10 semanas) (Gallagher & Reid 1979). En otro estudio, Merkt & Ellison (1998), comparando las costas del Atlántico suramericano y africano, encontraron variabilidad fenotípica significativa, lo que posiblemente indica diferencias genotípicas.
N. virginea presentó una amplio ámbito en la Ecorregión Darién y está en los dos únicos manglares conservados de la costa del Chocó y en Damaquiel en el Caribe antioqueño. Esta es una especie de amplio ámbito en el Caribe que se encuentra en praderas de pastos, manglares y puede subir varios kilómetros río arriba. Este amplio ámbito puede ser explicado por la naturaleza eurihalina observada en las formas adulta, juvenil y larval en los Neritidae (Crandall 1999), asociada posiblemente con su ciclo de vida anfídromo. Aunque no existen estudios al respecto, la evidencia indirecta sugiere que la larva es teleplánica, al igual que en otros Neritidae anfídromos (Myers et al. 2000, Cook et al. 2009). Por ello, las poblaciones de N. virginea de la Ecorregión del Darién podrían estar relacionadas con las de otras poblaciones del Caribe panameño presentes en los manglares y en las praderas de pastos, ambos bajo condiciones más marinas. Es posible que la relación sea mayor con esta área que con la del Caribe colombiano sur (entre la CGSM y golfo de Morrosquillo) debido a que la corriente predominante es occidente-oriente (Giro de Panamá-Colombia, Andrade et al. 2003). Además recientemente, se ha identificado que el patrón de vientos predominante norte-sur represa de manera significativa el caudal de agua dulce del río Atrato, al menos durante la época de seca (García 2007). Sin embargo, debido a esto posiblemente, las poblaciones ubicadas al Sur del delta del río Atrato podrían estar aún más aisladas de las que se encuentran en el Caribe. Las de la ensenada de Rionegro también podrían estar aisladas debido a su condición de laguna costera cuya conexión con el mar es intermitente. Trabajos basados en marcadores moleculares han encontrado que Neritidae anfídromos (N. virginea en Puerto Rico; Cook et al. 2009 y Clithon spinosus en la Polinesia Francesa; Myers et al. 2000) no presentaron estructura genética a ninguna escala de observación, es decir dentro de quebradas, entre quebradas dentro de una misma isla o aun entre islas distanciadas no solamente espacial sino temporalmente (diferente edad geológica). Sin embargo, a partir de estos estudios fue posible identificar varios haplotipos dentro de las poblaciones lo que sugiere los distintos orígenes de las larvas y vías predominantes de transporte como las corrientes marinas. En la Polinesia Francesa (Myers et al. 2000) no se encontró estructura genética aun entre islas separadas más de 140km, lo que apoya la hipótesis de una estrategia larval teleplánica (i.e. duración <1 año).
Hipótesis ecológica. La escasez de poblaciones establecidas de L. angulifera y de N. virgínea en el Caribe antioqueño y posiblemente en la mayor parte del Chocó podría ser explicada por el pobre estado de conservación de los manglares. Muchas áreas de manglar han sido taladas o erosionadas por los mares de leva y la deriva litoral y ahora permanecen unos pocos árboles en pie (Correa & Vernette 2004, Urrego et al. 2010). Como se explicó en la sección anterior, el aporte de larvas no parece ser una limitante para la distribución de ambas especies en la parte externa de la Ecorregión. De hecho, L. angulifera se asentó sobre troncos en las bermas de las playas, lo que sugiere una alta tolerancia a la emersión y desecación como ha sido observado en manglares de otras áreas geográficas (Tanaka & Maia 2006, Merk & Ellison 1998) y en otros littorinidos de los manglares del Pacífico colombiano (Blanco & Cantera 1999). En el golfo de Urabá se han observado individuos en las zonas intermareales altas en las playas y los manglares, pero en las bermas de las playas no se observan individuos grandes (JF Blanco, pers. observ.), posiblemente debido a que los troncos pueden ser movidos con facilidad durante los mares de leva (overwash), matando, así, a los reclutas recientes.
Aunque N. virginea es una especie eurihalina, parece ser sensible a la desecación y se encuentra predominantemente en charcos y neumatóforos o en las partes bajas de las raíces y troncos, desde donde se desplaza hacia el suelo cuando la marea empieza a bajar (JF Blanco, pers. observ.). Además, N. virginea es sensible a la alta turbiedad y a las altas concentraciones de fósforo total en los ríos costeros (Blanco & Scatena 2006). Esto podría explicar su ausencia en los manglares talados, contaminados con aguas residuales y con aguas turbias. Algunos caños del Caribe antioqueño drenan humedales que aportan muchos taninos y por lo tanto producen aguas naturalmente negras que podrían evitar la presencia de N. virginea en los manglares de sus desembocaduras. Ambas especies parecen necesitar los substratos duros provistos por los mangles. L. angulifera se observó dentro de los manglares predominantemente sobre troncos por encima de la línea de la marea, mas no en raíces bajas o en el suelo. Fuera de los manglares, se encontró en sustratos duros como troncos caídos en las bermas de las playas, rocas, e incluso sobre estructuras de muelles (i.e. bahía Turbo). En otras áreas geográficas constituye un importante componente de las comunidades de las costas rocosas (Reid 2001). Estas preferencias de sustrato difieren a lo planteado en diferentes estudios en el Caribe Colombiano que lo asocian exclusivamente a las raíces aéreas de Rhizophora mangle y con un amplio ámbito de salinidad (Blanco et al. 2010). Estudios sobre otros littorininos de manglar en Hong Kong y Brasil sugieren marcadas asociaciones con biotopos particulares (Lee & Williams 2002), diferencias entre especies de mangles y una alta fidelidad a los árboles (Tanaka & Maia 2006). Por lo anterior, se hipotetiza que la tala de árboles o su pérdida por erosión tiene efectos muy negativos sobre las poblaciones de L. angulifera ya que puede contribuir directamente a la mortalidad de los adultos establecidos. Finalmente, el impacto de los mares de leva sobre los sedimentos y superficies de los mangles podría ser otra explicación para la escasez de ambas especies dentro de los manglares del Caribe antioqueño y chocoano ya que casi todos ellos se encuentran desprotegidos y muchos muestran signos de erosión natural.
Se confirma así que L. angulifera es una especie de carácter estenohalina y marina, mientras que N. virginea es eurihalina, con predominancia estuarina, por lo tanto sus ámbitos en la Ecorregión Darién están influenciados por el aporte de agua dulce desde el río Atrato (preguntas uno, dos y tres). La salinidad también tiene una influencia sobre el ámbito de tallas de N. virginea en toda la Ecorregión. También se hipotetiza que la ausencia o limitada distribución en algunas localidades afuera del Golfo está explicada por el pobre estado de conservación del manglar (pregunta 3).
Agradecimientos
Agradecemos a la Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional y EAFIT por la financiación del proyecto “Expedición Estuarina, golfo de Urabá, Fase 1” dentro del megaproyecto Expedición Antioquia 2013. Queremos dar gracias al grupo de estudiantes y profesores del proyecto por su apoyo en el campo y en el laboratorio, así como por las discusiones sobre las ideas aquí presentadas. Se agradece especialmente a Edgar Andrés Estrada Urrea por su apoyo con los sistemas de información geográfica y la logística de campo. Los laboratorios de Limnología y Zoología General de la Universidad de Antioquia proveyeron espacio físico y equipos para procesar las muestras. Se agradece a la Corporación para el Desarrollo Sosteniblede la Región de Urabá (CORPOURABA) por proveer información primaria y secundaria. Los concejos comunitarios de Bocas del Atrato y del Chocó Norte-Río Acandí autorizaron el acceso a sus territorios. Contribución ELICE No. 4.
Referencias
Andrade, C.A., E.D. Barton & C.N. Mooers. 2003. Evidence for an eastward flow along the Central and South American Caribbean Coast. J. Geophys. Res. 108: 3185-3196. [ Links ]
Bernal, G., G. Poveda, P. Roldán & C. Andrade 2006. Patrones de variabilidad de las temperaturas superficiales del mar en la Costa Caribe Colombiana. Rev. Acad. Colomb. Cienc. 30: 195-208. [ Links ]
Blanco, J.F. & J.R. Cantera. 1999. The vertical distribution of mangrove gastropods and environmental factors relative to tide level at Buenaventura Bay, Pacific Coast of Colombia. Bull. Mar. Sci. 65: 617-630. [ Links ]
Blanco, J.F. & F.N. Scatena. 2005. Floods, habitat hydraulics and upstream migration of Neritina virginea (Gastropoda: Neritidae) in northeastern Puerto Rico. Caribb. J. Sci. 41: 55-74. [ Links ]
Blanco, J.F. & F.N. Scatena. 2006. Hierarchical contribution of river-ocean connectivity, water chemistry, hydraulics, and substrate to the distribution of diadromus snails in Puerto Rican streams. J. North Amer. Benthol. Soc. 25: 82-98. [ Links ]
Blanco, J.F. & F.N. Scatena. 2007. The spatial arrangement of Neritina virginea (Gastropoda: Neritidae) during upstream migration in a Split-channel reach. River Res. Appl. 23: 235-245. [ Links ]
Blanco, J.F. & A. Arroyave. 2009. Daños por depredación y tamaño de concha del caracol diádromo Neritina virginea (Gastropoda: Neritidae) en el Río Mameyes, Puerto Rico. Rev. Biol. Trop. 57: 1069-1080. [ Links ]
Blanco, J.F., M. Londoño-Mesa, L. Quan-Young, L. Urrego- Giraldo, J. Polanía, A. Osorio, G. Bernal & I. Correa 2011. The Urabá Gulf Mangrove Expedition of Colombia. ISME/GLOMIS 9: 8-10. [ Links ]
Cook, B.D., C.M. Pringle & J.M. Hughes. 2009. Marine dispersal determines the genetic population structure of migratory stream fauna of Puerto Rico: evidence for island-scale population recovery processes. J. North Amer. Benthol. Soc. 28: 709-718. [ Links ]
Correa, I.D. & G. Vernette. 2004. Introducción al problema de la erosión litoral en Urabá (sector Arbolete- Turbo) Costa Caribe colombiana. Bol. Invest. Mar. Cost. 33: 7-28. [ Links ]
Crandall, E.D. 1999. Early life history aspects of amphidromous neritid snails in Moorea, French Polynesia. Berkeley Sci. 2: 98-103 [ Links ]
Díaz, J.M. 1994. La malacofauna de la zona costera de Santa Marta y el Parque Nacional Natural Tayrona, Caribe Colombiano. An. Inst. Inv. Mar. Punta Betín 23: 15-43. [ Links ]
Díaz, J.M. 1995. Zoogeography of Marine Gastropod in the Southern Caribbean: A New Look at Provinciality. Caribb. J. Sci. 31: 104-121. [ Links ]
Ellison, A.M. 2008. Managing mangroves with benthic biodiversity in mind: Moving beyond roving banditry. J. Sea Res. 59: 2-15. [ Links ]
Gaines, M.S., J. Caldwell & A.M. Vivas. 1974. Genetic variation in the mangrove periwinkle Littorina angulifera. Mar. Biol. 27: 327-332. [ Links ]
Gallagher, S.B. & G.K. Reid. 1979. Population dynamics and zonation of the periwinkle Littoraria angulifera of the Tampa Bay, Florida, region. Nautilus 94:162-178. [ Links ]
García, J.M. & J. Palacio. 2008. Macroinvertebrados asociados a las raíces sumergidas del mangle rojo (Rhizophora mangle) en las bahías Turbo y El Uno, Golfo de Urabá (Caribe Colombiano). Ges. Amb. 11: 55-66. [ Links ]
García, C. (ed.). 2007. Atlas del golfo de Urabá: una mirada al Caribe de Antioquia y Chocó. Instituto de Investigaciones Marinas y Costeras (Invemar) y Gobernación de Antioquia. Serie de Publicaciones Especiales de Invemar Nº 12. Santa Marta, Colombia. [ Links ]
Guisande, C., A. Berrio, I. Maneiro, I. Riveiro, A. Vergara & A. Vaamonde. 2006. Tratamiento de datos. Díaz de Santos, Madrid, España. [ Links ]
Hogarth, P.J. 2007. The biology of mangroves and seagrasses. Oxford University, New York, EEUU. [ Links ]
Kyle, C.J. & E.G. Boulding. 2000. Comparative population genetic structure of marine gastropods (Littorina spp.) with and without pelagic larval dispersal. Mar. Biol. 137: 835-845. [ Links ]
Lee, S.Y. 2008. Mangrove macrobenthos: Assemblages, services, and linkages. J. Sea Res. 59: 16-29. [ Links ]
Llano, R. & J. Palacio. 1988. Ecología de la fauna asociada a raíces de mangle Rhizophora mangle en la Bahía de Marirrío Urabá. Tesis de pregrado, Instituto de Biología, Universidad de Antioquia, Medellín, Colombia. [ Links ]
Merkt, R.E. & A.M. Ellison. 1998. Geographic and habitatspecific morphological variation of Littoraria (Littorinopsis) angulifera (Lamarck, 1822). Malacologia 40: 279-295. [ Links ]
Myers, M.J., C.P. Meyer & V.H. Resh. 2000. Neritid and thiarid gastropods from French Polynesian streams: how reproduction (sexual, parthenogenetic) and dispersal (active, passive) effect population structure. Fresh Wat. Biol. 44: 535-545. [ Links ]
Ortiz, L.F. & J.F. Blanco. 2010. Moluscos (gasterópodos y bivalvos) asociados a las raíces sumergidas en los manglares del golfo de Urabá: patrones espaciales y determinantes a macroescala. Proyecto Expedición Estuarina, golfo de Urabá, fase 1. Informe final. Julio 2010. Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional Sede Medellín, Universidad EAFIT, Medellín, Colombia. [ Links ]
Osorio, A.F., L.G. Molina & J.D. Osorio. 2010. Caracterización hidrodinámica del oleaje local (sea) y de fondo (swell) en el golfo de Urabá. Proyecto Expedición Estuarina, golfo de Urabá, fase 1. Informe final. Julio 2010. Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional Sede Medellín, Universidad EAFIT, Medellín, Colombia. [ Links ]
Palacio, J. 1978. Variación de la fauna de invertebrados del área estuárica de la Ciénaga Grande de Santa Marta en relación con los cambios de salinidad. An. Inst. Inv. Mar. Punta Betín 10: 111-126. [ Links ]
Palacio, J. 1983. Die benthische Makroinvertebraten Fauna der Tropischen Ästuarregion Ciénaga Grande de Santa Marta (Kolumbien) und ihre aktivität im Wschsel zwischen Trockenund Regenzeit. Dissertation Zur erlangune des Grandes eines Doktors der Natur wissenschanften an der Ruhr, Universitat Bochum, Alemania. [ Links ]
Ramírez, A. 2005. Ecología Aplicada, Diseño y Análisis Estadístico. Universidad de Bogotá Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ]
Reid, D.G. 2001. New data on the taxonomy and distribution of the genus Littoraria Griffith & Pidgeon, 1834 (Gastropoda: Littorinidae) in Indo-West Pacific mangrove forest. Nautilus 115: 115-139. In K. Sanpanich, F.E. Wells & Y. Chitramvong. 2008. Reproduction and growth of Littoraria (Gatropoda: Littorinidae) at AngSila, Thailand. Raffles Bull. Zool. 18: 225-233. [ Links ]
Reyes, R. & N. Campos. 1992a. Macroinvertebrados colonizadores de raíces de Rhizophora mangle en la Bahía de Chengue, Caribe Colombiano. An. Inst. Inv. Mar. Punta Betín 21: 101-116. [ Links ]
Reyes, R. & N. Campos. 1992b. Moluscos, Anélidos y Crustáceos asociados a las raíces de Rhizophora mangle Linnaeus, en la región de Santa Marta, Caribe Colombiano. Caldasia 17: 133-148. [ Links ]
Riascos, J.M. & P.A. Guzmán. 2010. The ecological significance of growth rate, sexual dimorphism and size at maturity of Littoraria zebra and L. variegate (Gastropoda: Littorinidae). J. Mollusc. Stud. 76: 289-295. [ Links ]
Rojas, J. & J. Palacio. 2003. Contribución al conocimiento de la fauna de macroinvertebrados asociados a las raíces del manglar en el Golfo de Urabá. Trabajo de grado en Biología, Universidad de Antioquia, Medellín, Colombia. [ Links ]
Ruiz, M. & G. Bernal 2009. Variabilidad estacional e interanual del viento en los datos del reanálisis NCEP/ NCAR en la cuenca Colombia, mar Caribe. Avan. Rec. Hidraul. 20: 7-20. [ Links ]
Ruiz, M., G. Bernal, E. Beier & A. Osorio. 2010. La salinidad en la cuenca Colombia, mar Caribe, p. 39-43. In INVEMAR-ACIMAR (eds.). Libro de resúmenes extendidos XIV Seminario Nacional de Ciencia y Tecnología del Mar (SENALMAR). Serie de Publicaciones Especiales de Invemar No. 21. Santa Marta, Colombia. [ Links ]
Sanpanich, K., F.E. Wells & Y. Chitramvong. 2008. Reproduction and growth of Littoraria (Gatropoda: Littorinidae) at Ang Sila, Thailand. Raffles Bull. Zool. 18: 225-233. [ Links ]
Tanaka, M.O. & R.C. Maia. 2006. Shell morphological variation of Littoraria angulifera among and within mangroves in NE Brazil. Hydrobiology 559: 193-202. [ Links ]
Urrego, L.E., E.C. Molina, J.A. Suárez, H. Ruiz & J. Polanía. 2010. Distribución, composición y estructura de los manglares del golfo de Urabá. Proyecto Expedición Estuarina, golfo de Urabá, fase 1. Informe final. Julio 2010. Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional Sede Medellín, Universidad EAFIT, Colombia. [ Links ]
Vilardy, S. & J. Polanía. 2002. Mollusc fauna of the mangrove root-fouling community at the Colombian Archipelago of San Andrés and Old Providence. Wetl. Ecol. Manag. 10: 273-282. [ Links ]
Vermeij, G.J. 1971. Mollusk in mangrove swamps: Physiognomy, diversity and regional differences. Syst. Zool. 22: 609-624. [ Links ]
*Correspondencia a: Luis Ferney Ortiz. Grupo de Ecología Lótica: Islas, Costas y Estuarios (ELICE), Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Calle 67 Número 53-108, Medellín, Colombia. Apartado Aéreo 1226; lfortiz24@gmail.com
Juan Felipe Blanco. Grupo de Ecología Lótica: Islas, Costas y Estuarios (ELICE), Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Calle 67 Número 53-108, Medellín, Colombia. Apartado Aéreo 1226; blanco@exactas.udea.edu.co
1. Grupo de Ecología Lótica: Islas, Costas y Estuarios (ELICE), Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Calle 67 Número 53-108, Medellín, Colombia. Apartado Aéreo 1226; lfortiz24@gmail.com, blanco@exactas.udea.edu.co
Recibido 11-II-2011. Corregido 12-VII-2011. Aceptado 18-VIII-2011.


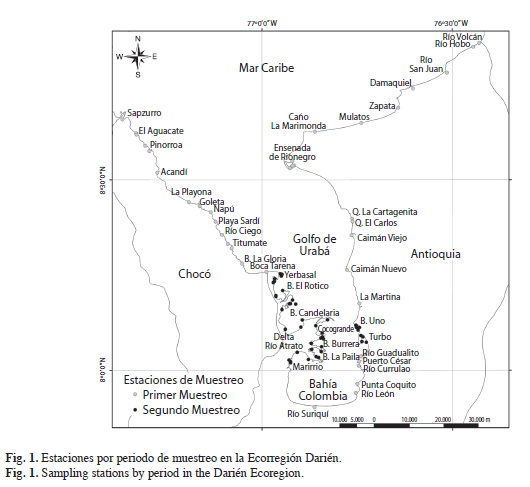{kind=link}
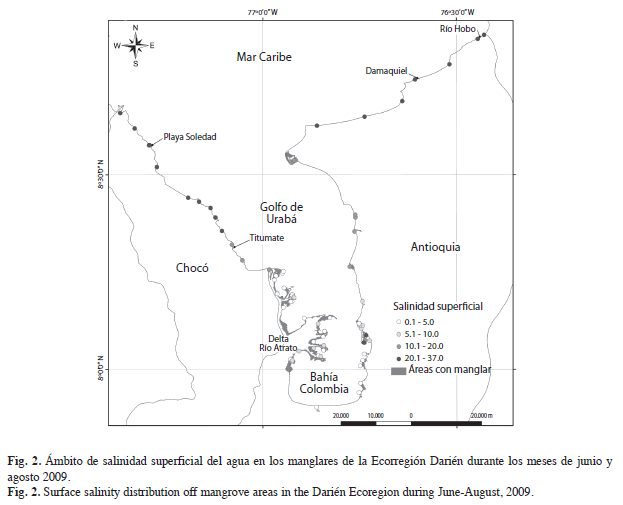{kind=link}
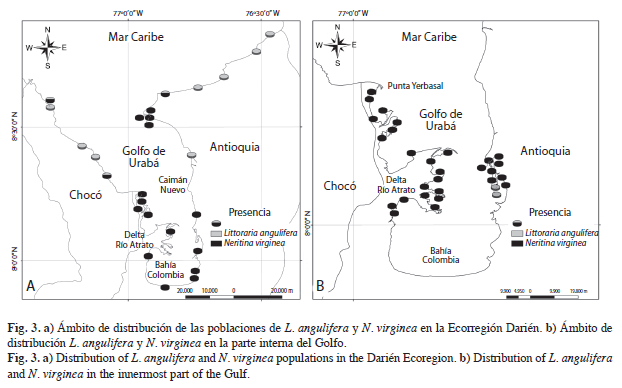{kind=link}
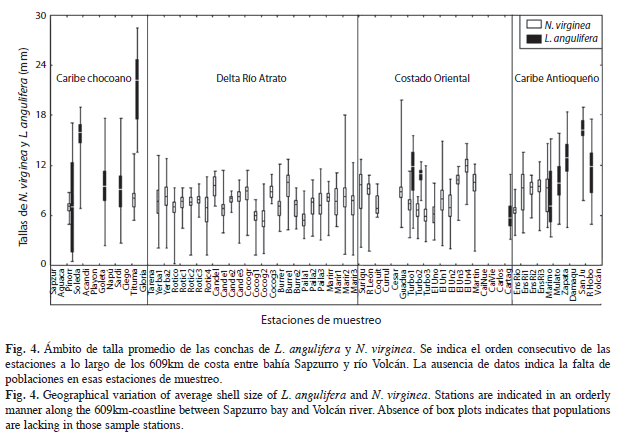{kind=link}