











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.59 no.4 San José Dez. 2011
Reproduction, pollination and seed predation of Senna multijuga (Fabaceae) in two protected areas in the Brazilian Atlantic forest
Marina Wolowski1 & Leandro Freitas2
1. Programa de Pós-Graduação em Botânica, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, rua Pacheco Leão 2040, CEP 22460-030, Rio de Janeiro, RJ, Brasil; marina.wolowski@gmail.com2. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, rua Pacheco Leão 915, CEP 22460-030, Rio de Janeiro, RJ, Brasil; leandro@jbrj.gov.br
Abstract
Key words: bee pollination, biodiversity conservation, bruchids, fecundity, fruit set, Leguminosae, phenology.
Resumen
Palabras clave: polinización de abejas, conservación de la biodiversidad, brúquidos, fecundidad, fructificación, frutos, Leguminosae, fenología.
One of the major strategies for the conservation of biodiversity is the establishment of protected areas, but it is recognised that there is a variety of approaches to establishing and managing such areas important to conservation, and what may be applicable in one place may not be in another (Dudley 2008). Although different kinds of protected areas can meet the general purposes defined in the categorization by the International Union for Conservation of Nature (IUCN) (Dudley 2008), the management activities within protected areas and the threats to the natural and cultural resources are distinct; moreover, they may differ in their conservation effectiveness (Hockings 2003, Gaston et al. 2008). Current knowledge on the extent to which protected areas fulfil their biodiversity protection goals is poor worldwide (Gaston et al. 2006), and there are few field assessments of the ecological effectiveness of protected areas (Brown et al. 2009). Protection effectiveness can be understood in terms of the different levels of hierarchical organization of biodiversity and the associated processes or functions, and it can be examined at several spatial scales (Gaston et al. 2006, 2008).
In spite of these many possibilities, the species is the most commonly used operational unit when addressing questions on biodiversity conservation (Scarano 2007). Inventories of the amount of biodiversity (i.e., number of species and ecosystems or communities) present in protected areas are the most common method used to assess their ecological effectiveness, and this practice is based on the assumption that inventory measures provide an indirect indication of condition of ecological processes within protected areas (Gaston et al. 2006). Nevertheless, criticism of the supposed relationship between biodiversity and ecosystem function (Srivastava & Vellend 2005, Scarano 2007) highlights the need for more direct measures of the persistence of biodiversity elements within protected areas. For instance, reproductive viability and mutualistic and antagonistic interactions are relevant indicators of the functioning and viability of plant populations in protected areas (Pauw 2007). However, to our knowledge, there is a lack of comparative studies on the effectiveness of protected areas with varying levels of restrictions (i.e., different management categories) in maintaining sexual reproduction in plants and their interactions with animals.
The Brazilian Atlantic Forest is a vast heterogeneous region (about 1.5 million km2), including a large variety of vegetation types and assemblages, large numbers of species (1-8% of the world’s total species), and numerous endemic species (Myers et al. 2000, Metzger 2009). This biome is highly imperilled and constitutes one of the most important biodiversity hotspots on Earth (Laurance 2009). Only 1.6% of the entire region is contained within protected areas, and these safeguard 9.3% of the remaining vegetation (11.7% of the original cover) (Ribeiro et al. 2009). We choose two contiguous protected areas in the Atlantic forest as our study system. The first is a national park, which has the aim of ecosystem protection and prohibits any direct use of natural resources. The second unit is an environmental protection area that is equivalent to the "protected landscape" according to the IUCN´s categories (Dudley 2008), which aims to maintain a harmonious interaction between nature and culture through the protection of landscapes and the continuation of traditional land uses, building practices and social and cultural manifestations. The two areas also differ in size and forest age, and in the degree of conservation, human presence and natural resources exploitation. Because of differences in the successional stage of and historical land use between the two areas, we expected higher effectiveness in the conservation of interactions within the more restrictive protected area. Specifically, we evaluated the phenology, pollination, reproductive success and pre-dispersal seed predation of Senna multijuga in those two areas.
Materials and methods
Study species: Senna multijuga is a medium- sized tree 10 to 15m in height that frequently occurs at altitudes between 50m and 950m in secondary forests, clearings, edges, regeneration areas and pastures from Central to South America (Irwin & Barneby 1982). Inflorescences are terminal panicules that bear hermaphrodite and zygomorphous flowers. Flowers have a yellow corolla with a mean diameter of 36.8±2.5mm (mean±standard deviation throughout the text) (n=10), diurnal anthesis, heteranthery, poricide dehiscence, and pollen as the only resource. Voucher (MWTorres 4) is deposited in the herbarium RB on Botanical Garden of Rio de Janeiro.
Only certain bees with the ability to vibrate can pollinate flowers with these characteristics (Buchmann 1983). Accordingly, pollinators of S. multijuga are large bees belonging to the genera Bombus, Centris, Epicharis and Xylocopa (Wolowski & Freitas 2010). Sexual reproduction depends on the transfer of pollen by pollinators, as no fruit is formed by selfpollination due to either inbreeding depression or by a mechanism of late-acting self-incompatibility (Wolowski & Freitas 2010). Seeds are predated by Bruchidae species of the genera Acanthoscelides and Sennius (Sari & Ribeiro-Costa 2005, Wolowski unpubl. res.).
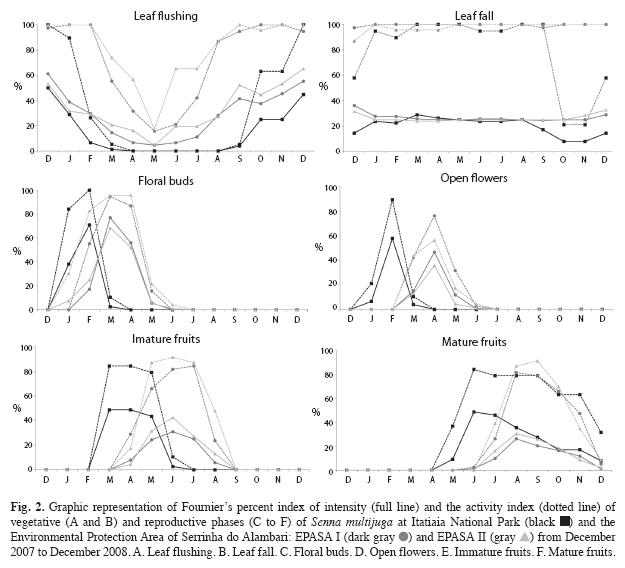
Phenology: Vegetative and reproductive phenodynamics were registered monthly from December 2007 to December 2008 in 80 individuals, respectively, 19 from INP, 38 from EPASA I and 23 from EPASA II. Presence and intensity of leaf flushing, leaf fall, floral buds, open flowers, and immature and mature fruits were recorded with binoculars. The Fournier’s percentage index of intensity (Fournier 1974) and the activity index (Bencke & Morellato 2002) were calculated.
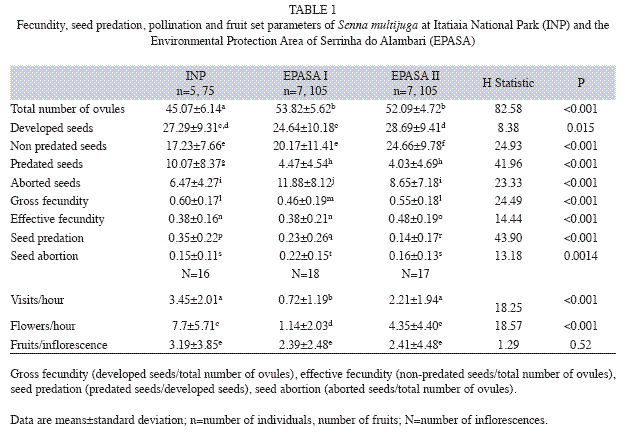
Fecundity and seed predation: 15 mature fruits per individual were collected prior to opening (in July and August 2008) from a total of 19 individuals, respectively, five from INP, seven from EPASA I and seven from EPASA II. They were dissected with nippers and stiletto and the number of non-developed ovules (i.e., scars of ovules) and seeds were counted with the help of a stereomicroscope. The number of developed seeds, aborted seeds (i.e., with incomplete development), predated seeds (i.e., developed seeds with the presence of bruchids or with predation orifices), and non-predated seeds were also counted. The total number of ovules (i.e., the sum of non-developed ovules, developed seeds, and aborted seeds) was calculated.
The following reproductive parameters were estimated: gross fecundity (number of developed seeds per total number of ovules), seed abortion (number of aborted seeds per total number of ovules), seed predation (number of predated seeds per number of developed seeds), and effective fecundity (number of non-predated seeds per total number of ovules).
Original and transformed data were tested for normality and homoscedasticity, and nonparametric statistical tests were performed in Statistica 6.0 (Statsoft 2001). The Wilcoxon paired sample test was used to compare phenodynamics between sites. The Kruskal- Wallis test (H) was used to compare all variables (fecundity, seed predation, pollination and fruit set) among sites and to verify possible differences in floral visitation among time intervals of observation in each area. Significant pairwise difference was tested with multiple comparisons.
Results
Phenology: Leaf flushing was not observed from April to August at INP, and it was sparse (<30%) at EPASA I and II in the same period (Fig. 2). Leaf fall was similar between the two areas and constant over time (Fig. 2), except from October to December. Activity and intensity indices of vegetative phases were statistically different (p<0.05) between the pairs INP-EPASA I (flush activity: Z=2.97; flush intensity: Z=3.18; fall activity: Z=2.55; fall intensity: Z=2.49) and INP-EPASA II (flush activity: Z=2.93; flush intensity: Z=3.18; fall activity: Z=2.50; fall intensity: Z=2.04). Leaf flushing activity at EPASA I differed from EPASA II (Z=2.40; p<0.05).
Flowering was annual with intermediary duration (sensu Newstrom et al. 1994). Flowering time differed between the areas, with a brief overlap (Fig. 2). Flowering peaks were two months apart. Fruit development started in March and extended to June at INP, while it was from April to August at EPASA I and II (Fig. 2). The timing of seed dispersal also differed, as the intensity of mature fruits was higher in June and July at INP and in August and September at EPASA I and II (Fig. 2). Activity and intensity indices of reproductive phases did not differ between sites (p>0.05), except floral buds activity between EPASA I and II (Z=2.20; p<0.05) and mature fruits intensity between INP and EPASA I (Z=2.52; p<0.05) and between INP and EPASA II (Z=2.38; p<0.05).
Fecundity and seed predation: Differences in overall fecundity were detected among the three sites; gross fecundity was higher at INP and EPASA II, and effective fecundity was higher at EPASA II (Table 1). Although the total number of ovules was higher at EPASA I and II, and the number of predated seeds and seed predation were higher at INP (Table 1).
Pollination and fruit set: The number of visits by pollinators was comparable among the different periods of observation for each site (INP: H=0.69, p=0.98; EPASA I: H=8.29, p=0.22; EPASA II: H=8.95, p=0.18). Thus, we merged the data and compared the accumulated frequencies among the inflorescences from the three sites. The frequency of visits and the number of flowers visited per hour were different among sites (Table 1). This difference was significant between INP and EPASA I (visits/hour and flowers visited/hour; p<0.001) and, between EPASA I and EPASA II (visits/hour; p=0.02 and flowers visited/hour; p=0.01) (Table 1). Fruit set was similar among sites (Table 1).
Discussion
The delayed occurrence of leaf flushing at EPASA may be due to differences in microclimate between the studied sites (Saunders et al. 1991, Murcia 1995), such as the higher solar radiation and temperatures at EPASA. Vegetative phenophases can have time-lag effects on reproductive phenology (Borchert 1992). This seems to be the case for S. multijuga, as it flowers after leaf flushing, which may explain the delayed flowering peak at EPASA. The time of fruit development and maturation of S. multijuga differed between areas. However, seed dispersal was in the dry season in both areas, when conditions are more favourable for anemocoric species (Frankie et al. 1974).
Effective fecundity affects other phases of the plant life cycle the most because it directly influences population recruitment. This parameter was higher in EPASA II than INP, because antagonistic interactions were more intense at the latter. It means that the relative reproductive success of S. multijuga reflects the balance between pollination and seed predation.
Pollinators are fundamental to the reproduction of S. multijuga (Wolowski & Freitas 2010). Pollination differences may be associated with factors that regulate bee populations, such as plant resources and nest sites in fragmented areas (Aizen & Feinsinger 1994b, Tscharntke et al. 1998, Cane 2001, Goulson et al. 2008, Steffan-Dewenter & Schiele 2008). Although differences in reproductive success may not be related to fragmentation (Aizen & Feinsinger 1994a, Dick 2001, Lopes & Buzato 2007, Ramos & Santos 2006, Dunley et al. 2009), pollination reduction negatively affects reproductive success in many selfincompatible species in fragmented habitats (Aizen & Feinsinger 1994a, Ghazoul et al. 1998, Gigord et al. 1999, Steffan-Dewenter et al. 2002, Aguilar & Galetto 2004, Aguilar et al. 2006). Then, we expected lower levels of pollination in EPASA II, because its intensive use of natural resources, but this was not observed. High frequency of visits to Senna multijuga in EPASA II may be due to lower richness and abundance of plant species and so fewer pollen resources for bees.
In contrast, higher seed predation at INP and EPASA I is consistent with other studies in which herbivory was greater in habitats with more vegetation cover (Cascante et al. 2002, Kolb et al. 2007, Herrerías-Diego et al. 2008). Specialisation, short life cycles and phenology synchronisation with the host plant can affect the population dynamics of bruchid species (Harper 1977) (i.e. the predators of seeds of S. multijuga). Moreover, these insects are sensitive to fluctuations in food resources and variation of abiotic factors, such as luminosity and humidity (Cascante et al. 2002, Lewis & Basset 2007).
Acknowledgments
We thank G. Bernadello, P.E.A.M. Oliveira and A.P. Fernandez for comments on the first version of this manuscript, V.S. Silvestre and L. Silva for field support, M.A. Santos, E.F. Gomes and M. Siqueira for mapping support and PROAP-CAPES and PETROBRAS-PMA for financial support. LF thanks CNPq for a researcher grant. This paper is part of the MSc dissertation of the first author presented to the ‘Programa de Pós-Graduação em Botânica, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro.
References
Aguilar, R. & L. Galetto. 2004. Effects of forest fragmentation on male and female reproductive success in Cestrum parqui (Solanaceae). Oecologia 138: 513-520. [ Links ]
Aguilar, R., L. Ashworth, L. Galetto & M.A. Aizen. 2006. Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecol. Lett. 9: 968-980. [ Links ]
Aizen, M.A. & P. Feinsinger. 1994a. Fragmentation, pollination and plant reproduction in a Chaco Dry Forest. Argentina. Ecology 75: 330-351. [ Links ]
Aizen, M.A. & P. Feinsinger. 1994b. Habitat fragmentation, native insect pollinators and feral honey bees in Argentine Chaco Serrano. Ecol. Appl. 4: 378-392. [ Links ]
Almeida, M.C. & S. Laroca. 1988. Trigona spinipes (Apidae, Meliponinae): Taxonomia, bionomia e relações tróficas em áreas restritas. Acta Biol. Par. 17: 67-108. [ Links ]
Bencke, C.S.C. & L.P.C. Morrelato. 2002. Comparação de dois métodos de avaliação da fenologia de plantas, sua interpretação e representação. Rev. Bras. Bot. 25: 269-275. [ Links ]
Bissonette, J.A. & I. Storch. 2002. Fragmentation: is the message clear? Conserv. Ecol. 6: 14 (Also available on line: www.consecolorg/vol6/iss2/art14). [ Links ]
Borchert, R. 1992. Computer simulation of tree growth periodicity and climatic hydroperiodicity in tropical forest. Biotropica 24: 385-395. [ Links ]
Brasil. 2000. SNUC Lei nº 9985, de 18 de julho de 2000 Institui o Sistema Nacional de Unidades de Conservação e dá outras providências. Brasília, DF. (Downloaded: February 3, 2008, www.planaltogovbr/ccivil/leis/L9985htm). [ Links ]
Brown, K.A., J.C. Ingram, D.F.B. Flynn, R. Razafindrazaka & V. Jeannoda. 2009. Protected area safeguard tree and shrub communities from degradation and invasion: a case study in eastern Madagascar. Environ. Manag. 44: 136-148. [ Links ]
Buchman, S.L. 1983. Buzz pollination in angiosperms, p. 73-113. In C.E. Jones & R.J. Little (eds.). Handbook of experimental pollination biology. Scientific and academic editions, New York, New York, USA. [ Links ]
Cane, J. 2001. Habitat fragmentation and native bees: a premature verdict? Conserv. Ecol. 5: 149-161. [ Links ]
Carvalho, P.E.R. 2004. Pau-Cigarra - Senna multijuga. Circular técnica 92: 1-11. (Downloaded: February 10, 2011, www.cnpf.embrapa.br/publica/circtec/edicoes/circ-tec92.pdf). [ Links ]
Cascante, A., M. Quesada, J.J. Lobo & E.A. Fuchs. 2002. Effects of dry tropical forest fragmentation on the reproductive success and genetic structure of the tree Samanea saman. Conserv. Biol. 16: 37-147. [ Links ]
Dick, C.W. 2001. Genetic rescue of remmant tropical trees by alien pollinator. Proc. Roy. Soc. Lond. B. Biol. Sci. 268: 2391-2396. [ Links ]
Dudley, N. 2008. Guidelines for applying protected area management categories. IUCN, Gland, Nyon, Switzerland. [ Links ]
Dunley, B., L. Freitas & L. Galetto. 2009. Reproduction of Byrsonima sericea (Malpighiaceae) in Restinga fragmented habitats in Southeastern Brazil. Biotropica 41: 1-8. [ Links ]
Fournier, L.A. 1974. Un método cuantitativo para la medición de características fenolólogicas en árboles. Turrialba 24: 422-423. [ Links ]
Frankie, G.N., H.G. Baker & P.A. Opler. 1974. Comparative phenological studies of trees in tropical wet and dry forest in the lowlands of Costa Rica. J. Ecol. 62: 881-913. [ Links ]
Gaston, K.J., K. Charman, S.F. Jackson, P.R. Armsworth, A. Bonn, R.A. Briers, C.S.Q. Callaghan, R. Catchpole, J. Hopkins, W.E. Kunin, J. Latham, P. Opdam, R. Stoneman, D.A. Stroud & R. Tratt. 2006. The ecological effectiveness of protected areas: The United Kingdom. Biol. Conserv. 132: 76-87. [ Links ]
Gaston, K.J., S.F. Jackson, L. Cantú-Salazar & G. Cruz-Piñon. 2008. The ecological performance of protected areas. Ann. Rev. Ecol. Evol. Systemat. 39: 93-113. [ Links ]
Ghazoul, J., K. Liston & T.J.B. Boyle. 1998. Disturbanceinduced density-dependent seed set in Shorea siamensis (Dipterocarpaceae), a tropical forest tree. J. Ecol. 86: 462-473. [ Links ]
Gigord, L., F. Picot & J.A. Shykoffb. 1999. Effects of habitat fragmentation on Dombeya acutangula (Sterculiaceae), a native tree on La Réunion (Indian Ocean). Biol. Conserv. 88: 43-51. [ Links ]
Goulson, D., G.C. Lye & B. Darwill. 2008. Decline and conservation of bumblebees. Ann. Rev. Entomol. 53: 191-208. [ Links ]
Harper, J.L. 1977. Population biology of plants. Academic, New York, New York, USA. [ Links ]
Herrerías-Diego, Y., M. Quesada, K.E. Stoner, J.A. Lobo, Y. Hernández-Flores & G.S. Montoya. 2008. Effect of forest fragmentation on fruit and seed predation of the tropical dry forest tree Ceiba aesculifolia. Biol. Conserv. 141: 241-248. [ Links ]
Hockings, M. 2003. Systems for assessing the effectiveness of management in protected areas. BioScience 53: 823-832. [ Links ]
Irwin, M.S. & R.C. Barneby. 1982. The american Cassinae a synoptical revision of Leguminosae tribe Cassieae subtribe Cassinae in the New World. Mem. New York Bot. Gard. 35: 1-918. [ Links ]
Kalil-Filho, N.A., A.F. Santos, A.P. Carvalho, A.C. Medeiros, A.R. Ferratti, A.C. Nogueira, C.A. Ferreira & P.E.R. Carvalho. 2002. Espécies recomendadas para a restauração da Mata Atlântica, p. 77-130. In A.P.M. Galvão & A.C.S. Medeiros (eds.). Restauração da Mata Atlântica em áreas de sua primitiva ocorrência natural. Embrapa, Colombo, Paraná, Brasil. [ Links ]
Kolb, A., R. Leimu & J. Ehrlén. 2007. Environmental context influences the outcome of a plant-seed predator interaction. Oikos 116: 864-872. [ Links ]
Laurance, W.F. 2009. Conserving the hottest of the hotspots Biol. Conserv. 142: 1137. [ Links ]
Lewis, O.T. & Y. Basset. 2007. Insect conservation in tropical forest, p 34-56. In A.J.A. Stewart, T.R. New & O.T. Lewis (eds.). Insect Conservation Biology. CABI, Wallingford, Oxfordshire, United Kingdom. [ Links ]
Lopes, L.E. & S. Buzato. 2007. Variaton in pollinator assemblages in a fragmented landscape and its effects on reproductive stages of a self-incompatible treelet, Psychotria suterella (Rubiaceae). Oecologia 154: 305-314. [ Links ]
Mendonça-Filho, C.V. 1996. Braúna, angico, jacarandá e outras leguminosas de Mata Atlântica: Estação Biológica de Caratinga, Minas Gerais. Fundação Botânica, Margaret Mee, Fundação Biodiversitas, AP.EBC, IEF, FZB-BH, SB-MG, Belo Horizonte, Minas Gerais, Brasil. [ Links ]
Metzger, J.P. 2009. Conservation issues in the Brazilian Atlantic forest. Biol. Conserv. 142: 1138-1140. [ Links ]
Murcia, C. 1995. Edge effects in fragmented forests: implications for conservation. Trees 10: 58-62. [ Links ]
Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A.B. da Fonseca & J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858. [ Links ]
Newstrom, L.E., G.W. Frankie & H.G. Baker. 1994. A new classification for plant phenology based on flowering patterns in lowland tropical rain forest trees at La Selva, Costa Rica. Biotropica 26: 141-159. [ Links ]
Pauw, A. 2007. Collapse of a pollination web in small conservation areas. Ecology 88: 1759-1769. [ Links ]
Ramos, F.N. & F.A.M. Santos. 2006. Floral visitors and pollination of Psychotria tenuinervis (Rubiaceae): distance from the anthropogenic and natural edges of an Atlantic forest fragment. Biotropica 39: 383-389. [ Links ]
Ribeiro, M.C., J.P. Metzger, A.C. Martensen, F.J. Ponzoni & M.M. Hirota. 2009. The Brazilian Atlantic forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conser. 142: 1141-1153. [ Links ]
Sari, L.T. & C.S. Ribeiro-Costa. 2005. Predação de sementes de Senna multijuga (Rich) HS Irwin and Barneby (Caesalpinaceae) por bruquídeos (Coleóptera: Chrysomelidae). Neotrop. Entomol. 34: 521-525. [ Links ]
Sari, L.T., C.S. Ribeiro-Costa & J. Roper. 2005. Dinâmica populacional de bruquíneos Coleoptera: Chrysomelidae) em Senna multijuga L.C. Richard I. & B. (Caesalpinaceae). Rev. Bras. Zool. 22: 169-174. [ Links ]
Saunders, D.A., R.J. Hobbs & C.R. Margules. 1991. Biological consequences of ecosystem fragmentation: a review. Biol. Conserv. 5: 18-32. [ Links ]
Scarano, F.R. 2007. Perspectives on biodiversity science in Brazil. Sciencia Agricola 64: 439-447. [ Links ]
Segadas-Vianna, F. & L. Dau. 1965. Ecology of Itatiaia range, southeastern Brazil II, Climates and altitudinal climatic zonation. Arquivos do Museu Nacional Rio de Janeiro 53: 31-53. [ Links ]
Srivastava, D.S. & M. Vellend. 2005. Biodiversity-ecosystem function research: Is it relevant to conservation? Ann. Rev. Ecol. Evol. Systemat. 36: 267-294. [ Links ]
StatSoft. 2001. STATISTICA: Data Analysis Software System version 60. StatSoft, Oklahoma, USA. [ Links ]
Steffan-Dewenter, I. & S. Schiele. 2008. Do resources or natural enemies drive bee population dynamics in fragmented habitats? Ecology 89: 1375-1387. [ Links ]
Steffan-Dewenter, I., U. Münzenberg & T. Tscharntke. 2001. Pollination, seed set and seed predation. Proc. Roy. Soc Lond. B. Biol. Sci. London B. 268: 1685-1690. [ Links ]
Steffan-Dewenter, I., U. Münzenberg, C. Büerger, C. Thies & T. Tscharntke. 2002. Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83: 1421-1432. [ Links ]
Tscharntke, T., A. Gathmann & I. Steffan-Dewenter. 1998. Bioindication using trap-nesting bees and wasps and their natural enemies: community structure and interactions. J. Appl. Ecol. 35: 708-719. [ Links ]
Umetsu, F. & R. Pardini. 2007. Small mammals in a mosaic of forest remmants and anthropogenic habitats – evaluating matrix quality in an Atlantic forest landscape. Landsc. Ecol. 22: 517-530. [ Links ]
Veloso, H.P., A.L.R. Rangel-Filho & J.C.A. Lima. 1991. Classificação da vegetação brasileira, adaptada a um sistema universal. Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro, Rio de Janeiro, Brasil. [ Links ]
Wolowski, M. & L. Freitas. 2010. Sistema reprodutivo e polinização de Senna multijuga (Fabaceae) em Mata Atlântica Montana. Rodriguésia 61: 167-179. [ Links ]
Correspondencia a: Marina Wolowski. Programa de Pós-Graduação em Botânica, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, rua Pacheco Leão 2040, CEP 22460-030, Rio de Janeiro, RJ, Brasil; marina.wolowski@gmail.com
Leandro Freitas. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, rua Pacheco Leão 915, CEP 22460-030, Rio de Janeiro, RJ, Brasil; leandro@jbrj.gov.br
Received 25-X-2010. Corrected 20-III-2011. Accepted 26-IV-2011.


{kind=link}
{kind=link}
{kind=link}