










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.4 San José Dec. 2011
Can rice field channels contribute to biodiversity conservation in Southern Brazilian wetlands?
Leonardo Maltchik1, Ana Silvia Rolon1,2, Cristina Stenert1,2, Iberê Farina Machado1 & Odete Rocha2
2. UFSCar, São Carlos, Brazil; asrolon@gmail.com, cstenert@unisinos.br, doro@ufscar.br
Abstract
Conservation of species in agroecosystems has attracted attention. Irrigation channels can improve habitats and offer conditions for freshwater species conservation. Two questions from biodiversity conservation point of view are: 1) Can the irrigated channels maintain a rich diversity of macrophytes, macroinvertebrates and amphibians over the cultivation cycle? 2) Do richness, abundance and composition of aquatic species change over the rice cultivation cycle? For this, a set of four rice field channels was randomly selected in Southern Brazilian wetlands. In each channel, six sample collection events were carried out over the rice cultivation cycle (June 2005 to June 2006). A total of 160 taxa were identified in irrigated channels, including 59 macrophyte species, 91 taxa of macroinvertebrate and 10 amphibian species. The richness and abundance of macrophytes, macroinvertebrates and amphibians did not change significantly over the rice cultivation cycle. However, the species composition of these groups in the irrigation channels varied between uncultivated and cultivated periods. Our results showed that the species diversity found in the irrigation channels, together with the permanence of water enables these man-made aquatic networks to function as important systems that can contribute to the conservation of biodiversity in regions where the wetlands were converted into rice fields. The conservation of the species in agriculture, such as rice field channels, may be an important alternative for biodiversity conservation in Southern Brazil, where more than 90% of wetland systems have already been lost and the remaining ones are still at high risk due to the expansion of rice production. Rev. Biol. Trop. 59 (4): 1895-1914. Epub 2011 December 01.
Key words: conservation, plant, macroinvertebrate, anuran, agroecosystem, cultivation cycle.
Resumen
La conservación de especies en los agroecosistemas ha llamado la atención. Los canales de riego pueden mejorar los hábitats y ofrecer condiciones para la conservación de especies de agua dulce. Desde el punto de vista de conservación de la biodiversidad surgen dos preguntas: 1) ¿Pueden los canales de riego mantener una rica diversidad de vegetación acuática, invertebrados y anfibios en el ciclo de cultivo del arroz? 2) ¿Puede la riqueza, abundancia y composición de especies acuáticas cambiar durante el ciclo de cultivo del arroz? Para ello, en el Condado Mostardas, Brazil un grupo de cuatro canales de los campos de arroz fue seleccionado al azar. En cada canal, se llevaron a cabo seis muestreos durante el ciclo de cultivo del arroz (junio 2005 hasta junio 2006). Un total de 160 táxones fueron identificados en los canales de riego, que incluyen 59 especies de macrófitos, 91 táxones de macroinvertebrados y 10 especies de anfibios. La riqueza y la abundancia de vegetación acuática, invertebrados y anfibios no cambió significativamente durante el ciclo de cultivo del arroz. Sin embargo, la composición de las especies de estos grupos en los canales de riego varía entre períodos no cultivados y cultivados. Nuestros resultados muestran que la diversidad de especies en los canales de riego, junto con la permanencia del agua permite que estas redes acuáticas provocadas por el ser humano puedan funcionar como sistemas importantes que contribuyan con la conservación de la biodiversidad en las regiones donde los humedales se han convertido en campos de arroz. La conservación de las especies en la agricultura, tales como los campos de los canales de arroz, que pueden ser una alternativa importante para la conservación de la biodiversidad en el sur de Brasil, donde más del 90% de los sistemas de humedales ya se han perdido y los restantes aún se encuentran en alto riesgo debido a la expansión de la producción de arroz.
Palabras clave: conservación, plantas, macroinvertebrados, anuros, agraecosistemas, ciclo de cultivo.
Wetlands are important sites for biological conservation, since they support a rich biodiversity and present high productivity (Mitsch & Gosselink 2000). However, biodiversity in wetlands has been reduced worldwide (Shine & Klemm 1999). Globally, more than 50% of these ecosystems were lost in the last century due to human activities (Shine & Klemm 1999). The impact of wetland loss on biodiversity was evidenced by the decline of populations of several wetland-dependent species (Millennium Ecosystem Assessment 2005). Agricultural expansion is one of the main human activities responsible for the decline of natural wetlands throughout the world, with severe consequences for species conservation (Czech & Parsons 2002). In 2003, approximately 151 million hectares of land were used worldwide for the cultivation of rice, and Asia accounted for 89% of that (FAOSTAT 2008). In some parts of Vietnam, India, and Thailand, wetlands were greatly reduced (Scott & Poole 1989). Worldwide, rice fields have been recognized as having considerable potential value for many species of aquatic invertebrates, plants, and vertebrates such as fish, amphibians, and birds (Czech & Parsons 2002, Bambaradeniya & Amarasinghe 2003, Elphick & Oring 2003) – likewise in Southern Brazil (Stenert et al. 2009, Machado & Maltchik 2010, Rolon & Maltchik 2010).
Based on the fact that agricultural wetlands, such as rice crops, depend on natural wetlands, the important question from the point of view of biodiversity conservation is whether these agricultural wetlands can function adequately or not; to maintain high levels of biodiversity. Therefore, the development of new concepts and management practices, which reconcile the sustainability of rice fields with the conservation of species, will demand greater understanding of these complex agroecosystems. For instance, Californian rice producers often flood their plantations after harvesting to accelerate straw decomposition. This may be an important strategy for biodiversity conservation, since such action creates important habitats for waterbird communities (Elphick & Oring 2003). Considering that protected areas cover only 11.5% of the surface of the planet (Rodrigues et al. 2004), rice fields and their large inundated areas may be essential in the conservation of wetland species.
Conservation of species in agroecosystems has attracted attention, mainly in Europe, where agricultural policy pays farmers to modify their farming practice to increase environmental benefits (CEC 1985). Ditches and ponds in agricultural landscapes offer conditions for freshwater species conservation (Twisk et al. 2000, 2003, Brainwood & Burgin 2009). Ditches are characterized as aquatic systems that remain in the landscape and improve habitats for the establishment of several organisms (Armitage et al. 2003, Herzon & Helenius 2008). Some studies showed the contribution of ditches to biodiversity conservation (Armitage et al. 2003, Mazerolle 2004, Goulder 2008), including in rice field systems (Bambaradeniya 2000, Katano et al. 2003). In the agricultural landscape, the role of ditches for biodiversity conservation can be compared with the natural waterbodies, contributing to the regional diversity (Armitage et al. 2003, Williams et al. 2003, Davies et al. 2008). Furthermore, the network of ditches can be used as ecological corridors for animal dispersion (Mazerolle 2004). The colonization of rice fields by different groups of organisms is related to water exchange between wetlands and rice fields through channels of irrigation (Fernando 1993).
Although, several studies address the importance of these channels for the conservation of biodiversity in temperate ecosystems (Herzon & Helenius 2008), no information regarding the importance is available from neotropical regions. In Southern Brazil, the rice crop area represents approximately 1 000 000ha (IRGA 2007) and irrigated channels are important elements in the agricultural landscape of Southern Brazil – corresponding to up 10% of the total farmland area. Studies with this approach are greatly needed in Southern Brazil, where more than 90% of wetland systems have already been lost, mainly due to rice field expansion.
Irrigated rice fields in Southern Brazil present a dynamic hydrologic regime, with variation between aquatic and terrestrial phases, and fields remaining without surface water for two years during the fallow phase. The fallow phase represents the period when the agricultural land remains without rice culture. In this phase, the agricultural land is drained and the surface water is not present. Nevertheless, the irrigated channels retained surface water throughout the cultivation cycle. Considering that rice fields maintain an important fraction of the macrophytes, macroinvertebrate and amphibians diversity of natural wetlands in Southern Brazil, and that agricultural practices influence the richness and composition of these communities over the cultivation cycle (Stenert et al. 2009, Machado & Maltchik 2010, Rolon & Maltchik 2010), two questions arises: 1) Can the irrigated channels maintain a rich diversity of macrophytes, macroinvertebrates and amphibians over the cultivation cycle? 2) Do richness, abundance and composition of aquatic species change over the rice cultivation cycle? Furthermore, we analyzed the influence of environmental variables (nitrate, soluble reactive phosphorus, conductivity, water temperature, depth and percentage of organic matter) on richness and abundance of macrophytes, macroinvertebrates and amphibians in irrigation channels.
Materials and methods
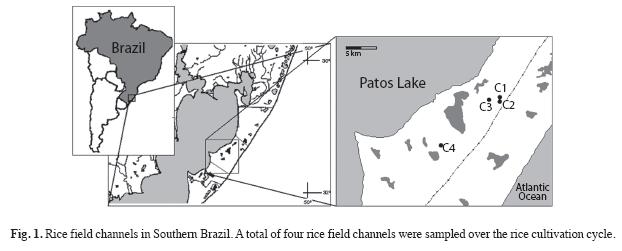
Study area: The state of Rio Grande do Sul (RS) is located in Southern Brazil (27º04’-33º45’ S, 49º42’-57º38’ W) and has an area of 282 184km2 (Fig. 1). The Coastal Plain of the State of Rio Grande do Sul is an important irrigated rice producer in South America (Azambuja et al. 2004), and also one of the regions with the highest concentration of wetlands (Maltchik 2003) and high aquatic species diversity (Rolon et al. 2008, Stenert et al. 2008). The climate is humid subtropical and the mean annual temperature is 17.5ºC. The mean annual rainfall reaches 1 250mm/yr and ranges between 1 150-1 450mm/yr (Tagliani 1995).
The study was carried out in Mostardas County (30º54’ 46.9" S - 50º48’46.2" W). Mostardas is the eighth largest producer of irrigated rice in Southern Brazil, presenting a rice crop area of approximately 33 397ha (IRGA 2007). A set of four rice field channels was randomly selected (Fig. 1). The water of the irrigation channels that supply the rice crops is captured from Patos Lake, a large coastal lagoon (10 200km2) (Fig. 1). The width of the studied channels was approximately 2m, and 100m in length.
Environmental data: In each of the four channels, six sample collection events were carried out over the rice cultivation cycle (June 2005 to June 2006): two collections in the fallow phase (June 2005 and September 2005), one collection during the tillage phase (November 2005), two collections in the rice growing season (rice emergence in January 2006 and tillering in March 2006) and one collection after harvesting (June 2006). The surface water exchange between the irrigation channels and the rice crops occurred during the cultivation phases (from January to March).
One sediment sample was collected per sampling occasion in each irrigation channel for granulometric analysis and organic matter percentage. The granulometric analysis was performed according to Suguio (1973). The sediment was classified into two categories: (1) sand (mainly composed of mineral particles larger than 0.05mm); and (2) silt (predominantly formed by fine mineral particles smaller than 0.05mm). The organic matter was determined by drying the sediment at 60ºC for 24 hours and burning it for five hours in an oven at 550ºC. Then, to describe the variation in nutrient concentration over the rice cultivation cycle, one sample of surface water was collected per sampling occasion (six phases) using polyethylene bottles (500mL) in each irrigation channel. All samples were placed on ice in dark containers and taken to the laboratory, and immediately filtered (Whatman® GF/F fibreglass filters, pore size 0.7mm). Concentrations of nitrate (mg/L), soluble reactive phosphorus (mg/L), and water conductivity (μS/cm) were obtained following the methods provided by APHA (1989). Water temperature was measured in situ using a mercury thermometer accurate to 0.1ºC and water depth was measured with a graduated rule at each sampling location. The parameters pH and dissolved oxygen were not measured due to the lack of Water Quality Checker.
Biological sampling: Aquatic plant richness and biomass were measured using the quadrat method (Downing & Anderson 1985). Three quadrats (50cmx50cm) were sampled at random per sampling occasion in each irrigation channel. The upland and the facultative upland species were excluded from sampling. All the above-ground biomass inside the quadrat was removed. Plant material was washed to remove periphyton and deposited organic and inorganic materials. Washed plants were separated by species and dried in an oven at 60°C until they reached a constant weight (ca. 72h). Aquatic plant species were identified following taxonomic keys specific to Southern Brazil (Irgang & Gastal 1996). The biomass was expressed in grams of dry weight per square meter (gDW/m2).
Macroinvertebrate richness and abundance were measured using two methods. The total richness and abundance was the sum of species and individuals found in the two methods, respectively. Three quantitative samples of benthic macroinvertebrates were randomly taken in each irrigation channel using a 7.5cm diameter core, inserted to a depth of 10cm in the sediment. Moreover, a qualitative sample was conducted on each sampling occasion through a kick net (D-shaped, 30cm width, 250mm mesh). The samples were collected by kicking up the substrate and then sweeping above the disturbed area to capture dislodged or escaping macroinvertebrates (Rosenberg et al. 1997). Three random sweeps of one meter each were carried out in each irrigation channel, encompassing ~1m2 of area sampled per channel. Sweeps were pooled into 3.5L plastic buckets and preserved in situ with 10% formaldehyde. In the laboratory, the samples were washed through a 250 mm sieve and leaves, stems, and other debris were removed. The resulting material was preserved with 80% ethanol. Macroinvertebrates were separated and identified to family or genus/species levels (whenever possible), according to Lopretto & Tell (1995), Merritt & Cummins (1996), and Fernández & Domínguez (2001). The macroinvertebrates were deposited in a Reference Collection of the Laboratory of Ecology and Conservation of Aquatic Ecosystems of UNISINOS. The macroinvertebrade and aquatic plants were sampled between 9:00AM and 13:00PM.
The amphibians were sampled through visual transects between sunset and 01:00AM (Crump & Scott 1994). A total of three random transects (15min) was conducted per collection in each irrigation channel. The sequence of the visited channels was altered in each sampling period. All of the observed individuals were counted and identified with species guides (Cei 1980, Loebmann 2005, Rosset 2008).
For data analysis, total biological richness was corresponded to the total number of taxa (aquatic macrophyte, macroinvertebrate and amphibian) in each irrigation channel per sampling occasion. Aquatic macrophyte and amphibian richness corresponded to the number of species and macroinvertebrate richness was quantified by the lowest taxonomic category possibly identifiable.
Differences of total biological richness (three groups) over time and differences among the mean richness of each taxonomic group (aquatic macrophytes, macroinvertebrates and amphibians) over time were tested using repeated measures ANOVA. Furthermore, differences in abundance of each taxonomic group over time were tested using repeated measures ANOVA. In order to assure homoscedasticity and normality of residuals, abundance values of three taxonomic groups were square-root transformed, and to compare abundance among three taxonomic groups, abundance values from each group were standardized (range 0-1). Levene’s test verified the homogeneity of variance, and the Mauchy’s sphericity test acknowledged the sphericity assumption. Assuming Mauchly’s sphericity, no adjustment was made for the F test.
The concordance patterns in species richness among the three taxonomic groups were tested using the Pearson correlation. The influence of environmental variables (nitrate, soluble reactive phosphorus, conductivity, percentage of organic matter, temperature and depth) on total biological richness and on each taxonomic group (macrophyte, macroinvertebrate and amphibian) was analyzed using multiple linear regression. The best fitted model was selected by AIC criteria (Akaike 1974) and by selection method of both directions (combination of forward and backward methods). In order to assure two assumptions of linear regression (homoscedasticity and normality of residuals), the values of conductivity, organic matter percentage and depth were square-root transformed. The analyses were performed using vegan package (Oksanen et al. 2009) in R statistical program version 2.9.0 (R Development Core Team 2009).
The concordance patterns among species composition of three taxonomic groups were tested using Mantel correlation test. Distance matrices used in Mantel tests were based on species abundance and were constructed using the Euclidean distance. The significance of correlations was tested by permutations (9 999 permutations).
The variation in species composition during the study period and the influence of the environmental variables on species composition were assessed through Canonical Correspondence Analysis (CCA). The ordination was performed using standardized abundance of species of four rice field channels over the rice cultivation cycle (six phases). Only species occurring in more than one sampling occasion were included in the analysis. In order to assure normality, all environmental variables were log-transformed. The significance of axis was tested by permutations (5 000 permutations). The best fitted model was selected by AIC criteria (Akaike 1974) and by selection method of both directions (combination of forward and backward methods).
A Permutation Multivariate Analysis of Variance was used to compare differences in species composition between agricultural periods (uncultivated and cultivated periods). To assess differences in macrophyte composition we used an Euclidean distance matrix and 9999 permutations. An Indicator Species Analysis (Dufrene & Legendre 1997) was performed to determine which species discriminated the two agricultural periods. The analysis was done using labdsv package (Roberts 2007), and the significance of the discriminating power was obtained by 9 999 permutations.
Results
A total of 160 taxa were identified in irrigated channels, including 59 macrophyte species, 91 taxa of macroinvertebrate and 10 amphibian species (Appendixes 1, 2, 3). The most frequent macrophyte species were: Limnobium laevigatum, Ludwigia peploides, Luziola peruviana, Salvinia herzogii and Salvinia minima, which were found in all irrigated channels and all phases of cultivation cycle. Hydrophytes (floating and submerged species) corresponded to 51% of all macrophyte species and emergent species corresponded to 49% of macrophyte diversity. Much of macrophytes in irrigation channels were amphibious plants (67%). Cyperaceae and Poaceae species corresponded to 85% of emergent plants. Macrophyte total richness varied from 22 to 34 species in irrigated channels and fluctuated between 15 to 33 species over the rice cultivation cycle.
A total of 13 800 individuals distributed among 91 macroinvertebrate taxa were collected. Aquatic insects accounted for most of the macroinvertebrate families observed (50%). Hyalella (Dogielinotidae) represented the majority of the individuals (23.2%), and oligochaetes and insects corresponded to 16.5% and 12.5% of the macroinvertebrates collected, respectively. Ten families were represented only by a single genus or species. The macroinvertebrate total richness ranged from 46 to 62 taxa in irrigated channels and fluctuated between 16 to 42 taxa over the rice cultivation cycle.
The amphibian species Leptodactylus latrans, Pseudis minuta, Dendropsophus sanborni and Hypsiboas pulchellus were present in all irrigation channels. L. latrans represented 36% of all individuals surveyed, P. minuta and Physalaemus biligonigerus corresponded to 26% and 15% of all individuals, respectively. The families with the highest representation in the irrigation channels were Hylidae (five species) and Leiuperidae (three species). Amphibian total richness varied from five to eight species in irrigated channels and fluctuated from one to seven species over the rice cultivation cycle.
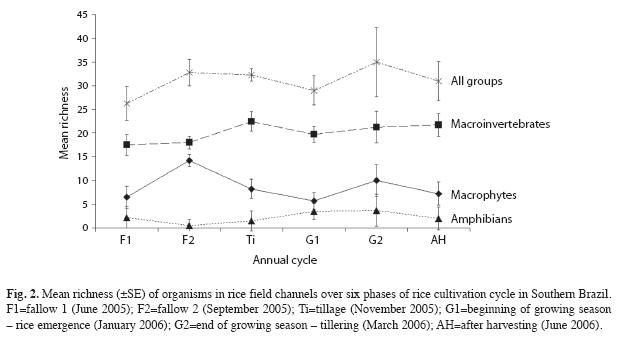
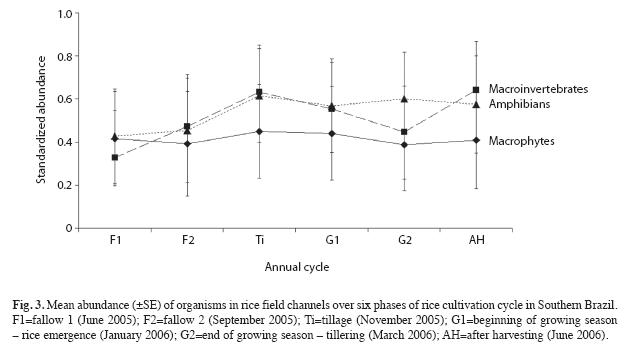
Total species richness did not change over the rice cultivation cycle (F5.15=0.720, p=0.619) (Fig. 2), and the pattern in variation of richness over the cultivation cycle was similar among taxonomic groups (F10.45=0.744, p=0.594) (Fig. 2). The abundance of the three taxonomic groups did not changed significantly over the rice cultivation cycle (F10.45=0.083, p=0.999) (Fig. 3).
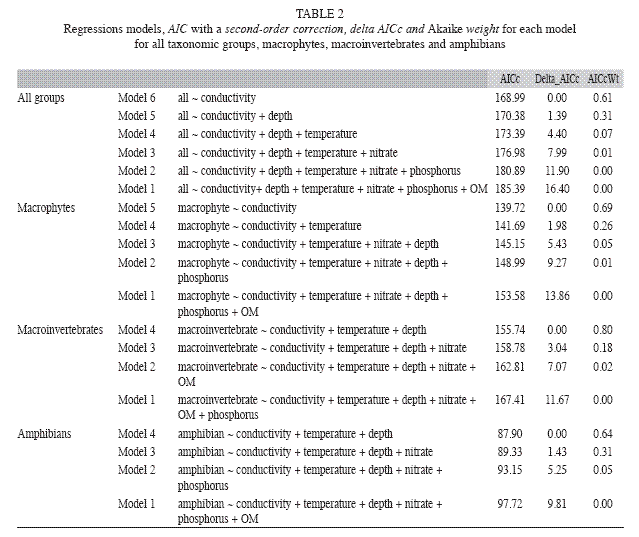
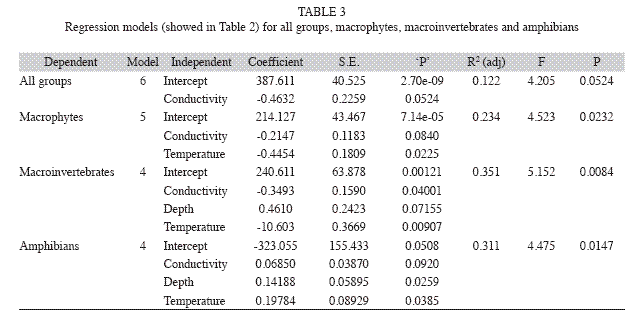
The richness of macrophytes, macroinvertebrates and amphibians were not correlated (Table 1). At marginal significance, the total richness was negatively correlated to water conductivity (R2adj=0.235, F1.22=4.523, p=0.052) (Table 2, 3). Macrophyte richness was negatively influenced by water conductivity and temperature (R2adj=0.122, F2.21=4.205, p=0.023) whereas macroinvertebrate and amphibian richness was influenced by water conductivity, temperature and depth (R2adj=0.351, F3.20=5.152, p=0.008; R2adj=0.312, F3.20=4.475, p=0.015, respectively) (Table 2, 3). Macroinvertebrate richness was positively related to water temperature and negatively related to water conductivity and depth, whereas amphibian richness was positively related to water conductivity, temperature, and depth (Table 2, 3).
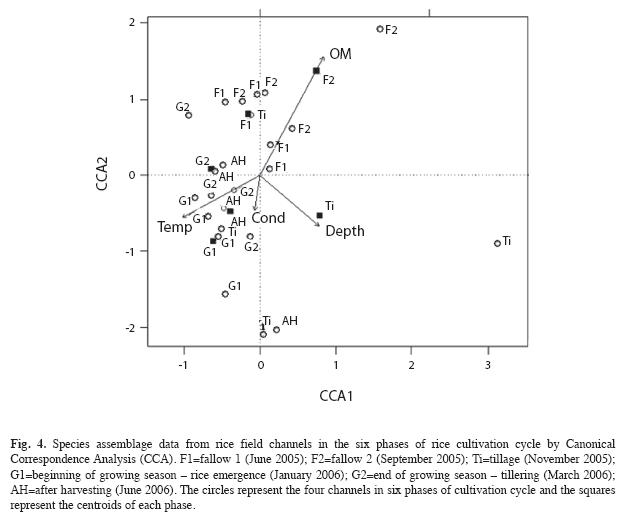
The composition of macrophyte, macroinvertebrate and amphibian assemblages in irrigation channels were not correlated (Table 1). The all canonical (constrained) axes explained 49% of the variation in species composition. The first two axes generated by CCA explained 33% of the variation explained by canonical axes (CCA1=19%, p=0.007 and CCA2=14%, p=0.042) (Fig. 4). The species-environment correlation was 0.97 for the first axis and 0.98 for the second. The environmental variables related to composition variation in irrigation channels were: cultivation phases, percentage of organic matter, conductivity, temperature, and depth. Temperature and depth were related to axis one and two, and percentage of organic matter and conductivity were associated to axis two (Fig. 4). According to the position of the centroids of the sampling phases in the ordination diagram, the species composition in second fallow phase and tillage phase differed from the others along the first axis and from each other along the second axis (Fig. 4).
Species composition in the irrigation channels varied between uncultivated and cultivated periods (F1.22=1.4517, p=0.0054). The uncultivated period was characterized by the aquatic macrophytes Azolla filiculoides (Indicator Species, IV=0.500, p=0.014), and Lemna valdiviana (IV=0.333, p<0.001). The cultivated period was characterized by the presence of the aquatic macrophyte Nymphoides indica (IV=0.332, p=0.038), by the aquatic insects Corixidae (IV=0.739, p=0.012), Belostomatidae (IV=0.594, p=0.022), and by the amphibian species Leptodactylus latrans (IV=0.738, p=0.001).
Discussion
Biodiversity conservation in agricultural landscapes is an ecological challenge (Marshall et al. 2003). Several studies have demonstrated the contribution of drainage channels in providing habitats for macrophyte, invertebrate and vertebrate establishment (Goulder 2008, Herzon & Helenius 2008). Macrophyte richness in the studied channels (59 species) correspond to about half of the total macrophyte richness of natural wetlands registered for this region (105 species) (Rolon et al. 2008). The number of macroinvertebrate families found in the irrigation channels (57) corresponded to more than 75% of the number of families observed in natural wetlands of Southern Brazil (Stenert & Maltchik 2007, Stenert et al. 2008). Additionally, the relative dominance of the aquatic insect families was similar to the dominance verified in Southern Brazil natural wetlands (Stenert & Maltchik 2007, Stenert et al. 2008). The 10 species of amphibians found in the irrigation channels corresponded to 63% of the richness observed in the studied region (Loebmann 2005), and represent 83% of the species that use rice crops as habitat in the studied region (Machado & Maltchik 2010). Our results suggest that such agroecosystem may support a large part of the biological diversity of the Brazilian Southern Coastal Plain.
Although, some species found in the irrigation channels occur in surrounding natural wetlands, the species composition of the irrigation channels surveyed during the present study differed from natural systems (Rolon et al. 2008, Stenert et al. 2008). The high proportion of hydrophytes (51% of species) was one of the differences found in the composition of the macrophytes in the irrigation channel, whereas in natural wetlands, the hydrophytes represented only a small part of the community (e.g. 25%, Rolon et al. 2004). The morphology of the channels (mean water depth of 60cm) limits the establishment of emergent species to the littoral zone; moreover, the depth of studied channels provided more availability of habitats for the establishment of hydrophytes. Another difference in the composition of macrophytes was the high proportion of grasses and sedges, which have represented almost the totality of the emergent species (85% of emergent species). The high dominance of grasses and sedges verified in studied channels (43%) was higher than observed in rice fields of Southern Brazil and Sri Lanka, where these species represented 46% and 39% of total richness, respectively (Bambaradenya et al. 2004, Rolon & Maltchik 2010).
Oligochaetes and Hyalella represented the majority of the invertebrate individuals collected (~40%) in the studied rice channels. This result was different from that observed in natural coastal wetlands of the same region, where aquatic insects were the most dominant macroinvertebrates (Stenert et al. 2008). In rice irrigation channels, aquatic plants can develop large biomasses (1-13t dry weight/ha), comprising the main sources of organic matter and nutrients (Roger & Watanabe 1984). The main part of the diet of Hyalella and oligochaetes consists of organic matter (debris and other types of food such as phytoplankton, periphytic algae, and bacteria (Brinkhurst et al. 1972, Brönmark & Hansson 1998), favouring the predominance of these two taxa in the rice irrigation channels. In addition, oligochaetes are important components of the benthic fauna in rice fields and their related water bodies (Bambaradeniya & Amerasinghe 2003). Oligochaete populations of about 10 000 and 40 000 individuals/m2 were found in Asian rice fields (Roger 1996). The amphibian community was dominated by L. latrans, P. minuta, and P. biligonigerus, corresponding to 77% of the individuals sampled. The predominance of these three species may be related to their ecological characteristics with regard to the use of the habitat (Loebmann 2005). While L. ocellatus and P. biligonigerus were more present at the margin of the systems, P. minuta was present exclusively in the water column of the channels.
The management of irrigation channels and the management practices used in the surrounding crops, influence the biological diversity of the rice crops (Twisk et al. 2000, Manhoudt et al. 2007). In the studied irrigation channels, the richness and abundance of macrophytes, macroinvertebrates and amphibians have not changed over the rice cultivation cycle. Despite variation of the water depth along the cycle (from 5 to 150cm), the presence of surface water during the entire period may be an important factor for the richness and abundance of species. Many studies point to the importance of the hydroperiod in the dynamic of aquatic communities in natural wetlands (Rolon et al. 2008, Stenert et al. 2008) and rice fields (Stenert et al. 2009, Machado & Maltchik 2010, Rolon & Maltchik 2010). The presence of water is critical for several species (Williams 1996), thus the surface water permanence in the irrigation channels may contribute to the dispersion of the organisms among the wetlands (Armitage et al. 2003, Mazerolle 2004).
Several studies have investigated concordant spatial patterns among taxonomic groups (Grenouillet et al. 2007), however, concordance of assemblage structure over time in managed systems remains unknown. Concordant patterns may result from biotic interactions among taxonomic groups, common environmental determinants or covariance of factors that affect different taxonomic groups independently (Heino 2002). The lack of concordant patterns in the species richness and composition of three taxonomic groups in irrigation channel can be related to differences in biotic response to environmental variables, such as, water conductivity, temperature and depth.
The water conductivity, temperature and depth were the variables that better explained the species richness in the irrigation channels studied. However, the influence of those variables was different among the taxonomic groups studied. The water conductivity influenced negatively the richness of macrophyte and macroinvertebrates, but it influenced positively the amphibian richness in irrigation channels. Some ecologists have assumed that water chemistry is an important predictor on macrophyte and invertebrate community in natural wetland systems (Batzer & Wissinger 1996, Thomaz et al. 2003). Strong variations of water chemistry can decrease the wetland macrophyte and macroinvertebrate richness (e.g. Friday 1987, Brodersen et al. 1998, Murphy et al. 2003), however, the water conductivity does not appear to have a great affect on amphibians, except in highly stressed anthropogenic situations (Beebee 1985). The water temperature negatively affected macrophyte richness and positively affected macroinvertebrate and amphibian richness, whereas, water depth influenced macroinvertebrates richness negatively and amphibian richness positively. Stenert et al. (2008) also found a negative relationship between water depth and macroinvertebrate richness in natural coastal wetlands of Southern Brazil. This pattern was also observed in natural wetlands of North America (Studinski & Grubbs 2007). At lower water depths, emergent vegetation can be used by macroinvertebrates as a substrate for the deposition of eggs, as a shelter against predation, and as food and support (Campeau et al. 1994). For amphibians temperature influences several amphibian physiological and behaviour processes, such as water balance, calling, metamorphosis, development and immune response (Rome et al. 1992), and the variations in water level may affect the abundance and diversity of species of amphibians in wetland systems (Pechmann et al. 1989). Other variables not measured in our study (e.g. pH, dissolved oxygen), also could have influenced the species richness in the rice field channels. Nonetheless, its effects can have been minimized since the total species richness observed did not change over the rice cultivation cycle. Besides, the water that supplies all rice field channels was captured from the same coastal lagoon.
Information regarding the biological diversity in agricultural areas and actions concerning the minimization of the impact of the agricultural activities on the conservation of biodiversity are extremely relevant to integrate biodiversity conservation in sustainable agriculture. The species diversity found in the irrigation channels together with the permanence of water enables these man-made aquatic networks to function as important systems that can contribute to the conservation of biodiversity in regions where the wetlands were converted into rice fields. However, it is important to mention that although such system may provide stability for populations between phases of the cultivation cycle, therefore contributing to the conservation of several species, the irrigation channels must not be viewed as surrogate systems for natural wetlands. The conservation of the species in rice field channels may be an important alternative for biodiversity conservation, where more than 90% of wetland systems have already been lost and the remaining ones are still at high risk due to the expansion of rice production. In this sense, management practices, that reconcile aquatic biodiversity conservation and rice production, such as, permanence of surface water in irrigation channels, could be important strategies to be considered during the agricultural development planning for Southern Brazil.
Acknowledgments
This research was supported by funds from UNISINOS (02.00.023/00-0) and CNPq (52370695.2). Leonardo Maltchik holds a Brazilian Research Council, CNPq Research Productivity grant. We thank the landowners that allowed access to rice fields on their properties. We declare that the data collection complied with the Brazilian current laws.
References
Akaike, H. 1974. A new look at the statistical model identification. IEEE Trans. Autom. Control 19: 716-723. [ Links ]
APHA– American Public Health Association. 1989. Standard methods for the examination of water and wastewater. American Public Health Association, American Water Works Association, and Water Pollution Control Federation, Washington, DC, USA. [ Links ]
Armitage, P.D., K. Szoszkiewicz, J.H. Blackburn & I. Nesbitt. 2003. Ditch communities: a major contributor to floodplain biodiversity. Aquat. Conserv. Mar. Freshw. Ecosyst. 13: 165-185. [ Links ]
Azambuja, I.H.V., J.F.J. Vernetti & J.A.M. Magalhães. 2004. Aspectos socioeconômicos da produção do arroz, p. 23-44. In A.S. Gomes & J.A.M. Magalhães (eds.). Arroz Irrigado no Sul do Brasil. Embrapa, Pelotas, Rio Grande do Sul, Brazil. [ Links ]
Bambaradeniya, C.N.B. 2000. Ecology and biodiversity in an irrigated rice field ecosystem in Sri Lanka. Ph.D. Thesis, University of Peradeniya, Peradeniya, Sri Lanka. [ Links ]
Bambaradeniya, C.N.B. & F.P. Amerasinghe. 2003. Biodiversity associated with the rice field agro-ecosystem in Asian countries: a brief review. International Water Management Institute (IWMI), Sri Lanka. (Downloaded: May 12, 2009, www.iwmi.cgiar.org/assessment/Research_Projects/working_papers.htm). [ Links ]
Bambaradeniya, C.N.B., J.P. Edirisinghe, D.N. Silva, C.V.S. Gunatilleke, K.B. Ranawana & S. Wijekoon. 2004. Biodiversity associated with an irrigated rice agroecosystem in Sri Lanka. Biodivers. Conserv. 13: 1715-1753. [ Links ]
Batzer, D.P. & S.A.Wissinger. 1996. Ecology of insect communities in nontidal wetlands. Annu. Rev. Entomol. 41: 75-100. [ Links ]
Beebee, T.J.C. 1985. Discriminant analysis of amphibian habitat determinants in south-east England. Amphibia-Reptilia 6: 35-43. [ Links ]
Brainwood, M. & S. Burgin. 2009. Hotspots of biodiversity or homogeneous landscapes? Farm dams as biodiversity reserves in Australia. Biodivers. Conserv. 18: 3043-3052. [ Links ]
Brinkhurst, R.O., K.E. Chua & N.K. Kaushick. 1972. Interspecific interactions and selective feeding of tubificid oligochates. Limn. and Oceanography 17: 122-133. [ Links ]
Brodersen, K.P., P.C. Dall & C. Lindegaard. 1998. The invertebrate fauna in the upper stony littoral of danish lakes: macroinvertebrates as trophic indicators. Freshw. Biol. 39: 577-592. [ Links ]
Brönmark, C. & L.A. Hansson. 1998. The biology of lakes and ponds. Oxford, New York, New York, USA. [ Links ]
Campeau, S., H.R. Murkin & R.D. Titman. 1994. Relative importance of algae and emergent plant litter to freshwater marsh invertebrates. Can. J. Fish. Aquat. Sci. 51: 681-692. [ Links ]
CEC- Commission of the European Communities. 1985. Perspectives for the common agricultural policy. COM, Brussels, Belgium. [ Links ]
Cei, J.M. 1980. Amphibians of Argentina. Monitore Zoologico Italiano 2: 1-609. [ Links ]
Crump, M.L. & N.J. Scott Jr. 1994. Standard techniques for inventory and monitoring: Visual encounter surveys, p. 84-92. In W.R. Heyer, M.A. Donnell, R.W. McDrarmid, L.C. Hayek & M.S. Foster (eds.). Measuring and monitoring biological diversity. Standard methods for amphibians. Smithsonian Institution, Washington, DC, USA. [ Links ]
Czech, H.A. & K.C. Parsons. 2002. Agricultural wetlands and waterbirds: a review. Waterbirds 25: 56-65. [ Links ]
Davies, B., J. Biggs, P. Williams, M. Whitfield, P. Nicolet, D. Sear, S. Bray & S. Maund. 2008. Comparative biodiversity of aquatic habitats in the European agricultural landscape. Agric. Ecosyst. Environ. 125: 1-8. [ Links ]
Downing, J.A. & M.R. Anderson. 1985. Estimating the standing biomass of aquatic macrophytes. Can. J. Fish. Aquat. Sci. 42: 1860-1869. [ Links ]
Dufrene, M. & P. Legendre. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67: 345-366. [ Links ]
Elphick, C.S. & L.W. Oring. 2003. Conservation implications of flooding rice fields on winter for waterbirds. Agric. Ecosyst. Environ. 94: 17-29. [ Links ]
FAOSTAT. 2008. FAO Statistical Databases. (Downloaded: October 14, 2009, http://faostat.fao.org). [ Links ]
Fernández, H.R. & E. Dominguez. 2001. Guía para la determinación de los artrópodos bentónicos sudamericanos. Universidad Nacional de Tucumán, Tucumán, Argentina. [ Links ]
Fernando, C.H. 1993. Rice field ecology and fish culture – an overview. Hydrobiologia 259: 91-113. [ Links ]
Friday, L.E. 1987. The diversity of macroinvertebrate and macrophyte communities in ponds. Freshw. Biol. 18: 87-104. [ Links ]
Goulder, R. 2008. Conservation of aquatic plants in artificial watercourses: Are main drains a substitute for vulnerable navigation canals? Aquat. Conserv. Mar. Freshw. Ecosyst. 18: 163-174. [ Links ]
Grenouillet, G., S. Brosse, L. Tudesque, S. Lek, Y. Baraillé & G. Loot. 2007. Concordance among stream assemblages and spatial autocorrelation along a fragmented gradient. Divers. Distrib. 14: 592-603. [ Links ]
Heino, J. 2002. Concordance of species richness patterns among multiple freshater taxa: a regional perspective. Biodivers. Conserv. 11: 137-147. [ Links ]
Herzon, I. & J. Helenius. 2008. Agricultural drainage ditches, their biological importance and functioning. Biol. Conserv. 141: 1171-1183. [ Links ]
IRGA – Instituto Riograndense do Arroz. 2007. Arroz irrigado: safra 2006/2007. Seção de Política Setorial – DCI, Porto Alegre, Rio Grande do Sul, Brazil. [ Links ]
Irgang, B.E. & C.V.S. Gastal Jr. 1996. Aquatic macrophytes of Coastal Plain of RS. Porto Alegre, Rio Grande do Sul, Brazil. [ Links ]
Katano, O., K. Hosoya, K. Iguchi, M. Yamaguchi, Y. Aonuma & S. Kitano. 2003. Species diversity and abundance of freshwater fishes in irrigation ditches around rice fields. Environ. Biol. Fishes 66: 107-121. [ Links ]
Loebmann, D. 2005. Guia Ilustrado: Os anfíbios da região costeira do extremo sul do Brasil. USEB, Pelotas, Rio Grande do Sul, Brazil. [ Links ]
Lopretto, E.C. & G. Tell. 1995. Ecosistemas de aguas continentales: metodologías para su estudio. Ediciones Sur, La Plata, Argentina. [ Links ]
Machado, I.F. & L. Maltchik. Can management practices in rice fields contribute to amphibian conservation in southern Brazilian wetlands? Aquatic Conserv. Mar. Freshw. Ecosyst. 20: 39-46. [ Links ]
Maltchik, L. 2003. Three new wetlands inventories in Brazil. Interciencia 28: 421-423. [ Links ]
Manhoudt, A.G.E., A.J. Visser & G.R. Snoo. 2007. Management regimes and farming practices enhancing plant species richness and ditch banks. Agric. Ecosyst. Environ. 119: 353-358. [ Links ]
Marshall, E.J.P., V.K. Brown, N.D. Boatman, P.J.W. Lutman, G.R. Squire & L.K. Ward. 2003. The role of weeds in supporting biological diversity within crop fields. Weed Res. 43: 77-89. [ Links ]
Mazerolle, M.C. 2004. Drainage ditches facilitate frog movements in a hostile landscape. Landscape Ecol. 20: 579-590. [ Links ]
Merritt, R.W. & K.W. Cummins. 1996. An introduction to the aquatic insects of North America. Kendall/Hunt, Dubuque, Iowa, USA. [ Links ]
Millennium Ecosystem Assessment. 2005. Ecosystems and human well-being: wetlands and water synthesis. World Resources Institute, Washington, USA. [ Links ]
Mitsch, W.J. & J.G. Gosselink. 2000. Wetlands. Wiley, New York, New York, USA. [ Links ]
Murphy, K.J., G. Dickinson, S.M. Thomaz, L.M. Bini, K. Dick, K. Greaves, M.P. Kennedy, S. Livingstone, H. McFerran, J.M. Milne, J. Oldroyd & R.A. Wingfield. 2003. Aquatic plant communities and predictors of diversity in a sub-tropical river floodplain: the upper Rio Paraná, Brazil. Aquat. Bot. 77: 257-276. [ Links ]
Oksanen, J., R. Kindt, P. Legendre, B. O’Hara, G.L. Simpson, P. Solymos, M.H.H. Stevens & H. Wagner. 2009. Vegan: Community ecology package. R package version 1.15-2. (Downloaded: March 10, 2011, CRAN.R-project.org/package=vegan). [ Links ]
Pechmann, J.H.K., D.E. Scott, J.W. Gibbons & R.D. Semlitsch. 1989. Influence of wetland hydroperiod on diversity and abundance of metamorphosing juvenile amphibians. Wetlands Ecol. Manage. 1: 3-11. [ Links ]
R Development Core Team. 2009. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. (Downloaded: March 10, 2011, www.R-project.org). [ Links ]
Roberts, D.W. 2007. Labdsv: Ordination and multivariate analysis for ecology. R package version 1.3-1 (Downloaded: March 10, 2011, http://ecology.msu.montana.edu/labdsv/R). [ Links ]
Rodrigues, A.S.L., S.J. Andelman, M.I. Bakarr, L. Boitani, T.M. Brooks, R.M. Cowling, L.D.C. Fishpool, G.A.B. Fonseca, K.J. Gaston, M. Hoffmann, J.S. Long, P.A. Marquet, J.D. Pilgrim, R.L. Pressey, J. Schipper, W. Sechrest, S.N. Stuart, L.G. Underhill, R.W. Waller, M.E.J. Watts & X. Yan. 2004. Effectiveness of the global protected area network in representing species diversity. Nature 428: 640-643. [ Links ]
Roger, P.A. 1996. Biology and management of floodwater ecosystem in rice fields. International Rice Research Institute, Manila, Philippines. [ Links ]
Roger, P.A. & I. Watanabe. 1984. Algae and aquatic weeds as a source of organic matter and plant nutrients for wetland rice, p. 147-168. In S. Banta (ed.). Organic matter and rice. International Rice Research Institute, Manila, Philippines. [ Links ]
Rolon, A.S., T. Lacerda, L. Maltchik & D.L. Guadagnin. 2008. The influence of area, habitat and water chemistry on richness and composition of macrophyte assemblages in southern Brazil wetlands. J. Veg. Sci. 19: 221-228. [ Links ]
Rolon, A.S. & L. Maltchik. 2010. Does flooding of rice fields after cultivation contribute to wetland plant conservation in southern Brazil? Appl. Veg. Sci. 13: 26-35. [ Links ]
Rolon, A.S., L. Maltchik & B.E. Irgang. 2004. Levantamento de macrófitas aquáticas em áreas úmidas do Rio Grande do Sul, Brasil. Acta Biologica Leopoldensia 26: 17-35. [ Links ]
Rome, L.C., E.D. Stevens & H.B. John-Adler. 1992. The influence of temperature and thermal acclimation on physiological function, p. 183-205. In M.E. Feder & W.W. Burggren (eds.). Environmental physiology of the amphibians. University of Chicago, Chicago, USA. [ Links ]
Rosenberg, D.M., I.J. Davies, D.G. Cobb & A.P. Wiens. 1997. Ecological monitoring and assessment network (EMAN – Environment Canada) – Protocols for measuring biodiversity: Benthic macroinvertebrates in freshwaters. Department of Fisheries & Oceans. Freshwater Institute, Winnipeg, Manitoba, Canada. [ Links ]
Rosset, S.D. 2008. New species of Odontophrynus (Reinhardt & Lütken 1862) (Anura: Neobatrachia) from Brazil and Uruguay. J. Herpetol. 42: 134-144. [ Links ]
Scott, D.A. & C.M. Poole. 1989. A status overview of Asian wetlands. Asian Wetland Bureau, Kuala Lumpur, Malaysia. [ Links ]
Shine, C. & C. Klemm. 1999. Wetlands, water and the law: Using law to advance wetland conservation and wise use. IUCN, Gland, Switzerland. [ Links ]
Stenert, C., R.C. Bacca, L. Maltchik & O. Rocha. 2009. Can hydrologic management practices of rice fields contribute to macroinvertebrate conservation in southern Brazil wetlands? Hydrobiologia 635: 339-350. [ Links ]
Stenert, C., R.C. Bacca, C.C. Mostardeiro & L. Maltchik. 2008. Environmental predictors of macroinvertebrate communities in coastal wetlands of southern Brazil. Mar. Freshw. Res. 59: 540-548. [ Links ]
Stenert, C. & L. Maltchik. 2007. Influence of area, altitude and hydroperiod on macroinvertebrate communities in southern Brazil wetlands. Mar. Freshw. Res. 58: 993-1001. [ Links ]
Studinski, J.M. & S.A. Grubbs. 2007. Environmental factors affecting the distribution of aquatic invertebrates in temporary ponds in Mammoth Cave National Park, Kentucky, USA. Hydrobiologia 575: 211-220. [ Links ]
Suguio, K. 1973. Introdução a sedimentologia. Edgard Blucher, São Paulo, São Paulo, Brazil. [ Links ]
Tagliani, P.R.A. 1995. Estratégia de planificação ambiental para o sistema ecológico da Restinga da Lagoa dos Patos - Planície Costeira do Rio Grande do Sul. Ph.D. Thesis. Federal University of São Carlos, São Carlos, Brazil. [ Links ]
Thomaz, S.M., D.C. Souza & L.M. Bini. 2003. Species richness and beta diversity of aquatic macrophytes in a large subtropical reservoir (Itaipu Reservoir, Brazil): the limnology and morphometry. Hydrobiologia 505: 119-128. [ Links ]
Twisk, W., M.A.W. Noordervliet & W.J. Keurs. 2000. Effects of ditch management on caddisfly, dragonfly and amphibian larvae in intensively farmed peat areas. Aquat. Ecol. 34: 397-411. [ Links ]
Twisk, W., M.A.W. Noordervliet & W.J. Keurs. 2003. The nature value of the ditch vegetation in peat areas in relation to farm management. Aquat. Ecol. 37: 191-209. [ Links ]
Williams, D.D. 1996. Environmental constraints in temporary freshwater and their consequences for the insect fauna. J. N. Am. Benthol. Soc. 15: 634-650. [ Links ]
Williams, P., M. Whitfield, J. Biggs, S. Bray, G. Fox, P. Nicolet & D. Sear. 2003. Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol. Conserv. 115: 329-341. [ Links ]
Correspondencia a:Leonardo Maltchik & Iberê Farina Machado. Laboratory of Ecology and Conservation of Aquatic Ecosystems Av. Unisinos, 950 CEP 93.022-000, UNISINOS, São Leopoldo, RS, Brazil; maltchik@unisinos.br, ibermachado@gmail.com
Ana Silvia Rolon & Cristina Stenert. Laboratory of Ecology and Conservation of Aquatic Ecosystems Av. Unisinos, 950 CEP 93.022-000, UNISINOS, São Leopoldo, RS, Brazil / UFSCar, São Carlos, Brazil; asrolon@gmail.com, cstenert@unisinos.br
Odete Rocha. UFSCar, São Carlos, Brazil; doro@ufscar.br
Received 03-IX-2010. Corrected 10-II-2011. Accepted 15-III-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}