










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.4 San José Dec. 2011
Adriana Guzmán1 & Pablo R. Stevenson2
1. Fundación Malpelo y Otros Ecosistemas Marinos, Carrera 7 No. 32-33, piso 27. Bogotá, Colombia; aguzman@fundacionmalpelo.org
2. Laboratorio de Ecología de Bosques Tropicales y Primatología, Universidad de Los Andes, Carrera 1 No. 18-10. Bogotá, Colombia; pstevens@uniandes.edu.co
Abstract
Most studies on seed dispersal in time have focused on seed dormancy and the physiological triggers for germination. However, seed dispersed by animals with low metabolic and moving rates, and long gutpassage times such as terrestrial turtles, could be considered another type of dispersal in time. This study tests the hypothesis that seeds dispersed in time may lower predation rates. We predicted that seeds deposited below parent trees after fruiting fall has finished is advantageous to minimize seed predators and should show higher survival rates. Four Amazonian plant species, Dicranostyles ampla, Oenocarpus bataua, Guatteria atabapensis and Ocotea floribunda, were tested for seed survival probabilities in two periods: during fruiting and 10-21 days after fruiting. Experiments were carried out in two biological stations located in the Colombian Amazon (Caparú and Zafire Biological Stations). Seed predation was high and mainly caused by non-vertebrates. Out of the four plant species tested, only Guatteria atabapensis supported the time escape hypothesis. For this species, seed predation by vertebrates after the fruiting period increased (from 4.1% to 9.2%) while seed predation by nonvertebrates decreased (from 54.0% to 40.2%). In contrast, seed predation by vertebrates and by non-vertebrates after the fruiting period in D. ampla increased (from 7.9% to 22.8% and from 40.4% to 50.6%, respectively), suggesting predator satiation. Results suggest that for some species dispersal in time could be advantageous to avoid some type of seed predators. Escape in time could be an additional dimension in which seeds may reach adequate sites for recruitment. Thus, future studies should be address to better understand the survival advantages given by an endozoochory time-dispersal process. Rev. Biol. Trop. 59 (4): 1795-1803. Epub 2011 December 01.
Key words: Janzen-Connell theory, endozoochory, escape hypothesis, tropical forest, seed dispersal, seed survival.
Resumen
La mayoría de estudios sobre dispersión de semillas en el tiempo tratan sobre la dormancia de las semillas y los procesos fisiológicos que disparan su germinación. Sin embargo, la dispersión de semillas por animales de bajas tasas metabólicas y largos tiempos de retención, como por ejemplo los reptiles, podría ser considerada otro tipo de dispersión en el tiempo. Este estudio prueba la hipótesis que las semillas dispersadas por tortugas pueden evadir a los depredadores en el tiempo. Semillas depositadas bajo árboles parentales luego de que la cosecha haya terminado es ventajoso para escapar de depredadores denso-dependientes y por lo tanto deberían mostrar mayores tasas de supervivencia. La hipótesis se probó en cuatro especies de plantas amazónicas, Dicranostyles ampla, Oenocarpus bataua, Guatteria atabapensis y Ocotea floribuna, durante dos periodos: durante la cosecha y varios días después de la cosecha de frutos. Los experimentos se llevaron en dos estaciones biológicas de la Amazonia colombiana (Caparú y Zafire). Los principales depredadores de semillas fueron los no vertebrados y por lo tanto fueron el factor que más influyó en la supervivencia de las semillas en ambas áreas de estudio. De las cuatro especies probadas, sólo Guatteria atabapensis validó la hipótesis de la ventaja de la dispersión en el tiempo. Para esta especie, la depredación de semillas por vertebrados después del periodo de cosecha incrementó (de 4.1% a 9.2%) mientras que la depredación de semillas por no vertebrados disminuyó (de 54.0% a 40.2%). Por el contrario, la depredación de semillas por vertebrados y por no vertebrados después del período de la cosecha para D. ampla incrementó (de 7.9% a 22.8% y de 40.4% a 50.6% respectivamente), sugiriendo saciación de depredadores. Los resultados sugieren que para algunas especies, el escape en el tiempo podría ser ventajoso para evadir algunos tipos de depredadores. El escape en el tiempo podría ser una dimensión adicional en donde las semillas podrían alcanzar lugares adecuados para su reclutamiento. Futuros estudios deberían realizarse con el fin de entender mejor las ventajas de supervivencia dadas por la dispersión en el tiempo luego de un proceso de endozoocoria.
Palabras clave: hipótesis del escape, endozoocoria, bosques tropicales, dispersión de semillas, supervivencia de semillas, teoría Janzen-Connell.
The Janzen-Connell hypothesis proposes that seeds have higher survival probabilities when they escape distance-and density-dependent predation below parental trees, affecting plant recruitment and diversity (Janzen 1970, Connell 1971). Thus, seed dispersal away from parent trees become important for plant fitness and survival. Some predictions of the Janzen-Connell hypothesis have been tested in various plant species and habitats, generally providing positive support (Hammond & Brown 1998, Wright 2002, Hyatt et al. 2003, Peterman et al. 2008, Bagchi et al. 2010, Matthesius et al. 2010). However seed dispersal is a process in which seeds may escape enemies in space, but also in time.
Studies on seed dispersal in time have focused on seed dormancy and the physiological triggers for germination (Cohen & Levin 1987, Imbert 1999, Lehouck et al. 2009, Cousens et al. 2010). Also, seed dispersal in time has been proposed when delays in the flowering or fruiting phenologies occur, thus seeds are dispersed later and they either escape predation or suffer unusual high mortality (Augspurger 1981, Curran & Webb 2000, Chuine 2010). A similar effect could be accomplished by endozoochorous seed dispersers due to time spent passing through the digestive system. Gut passage time periods can vary from individuals to species (Lambert 1998), and very long gut passage times have been documented in some chelonian species (i.e. up to 42 days in Chelonoidis spp.) without affecting seed germination (Guzmán 2005, Strong & Fragoso 2006, Guzmán & Stevenson 2008). In general, transit times in terrestrial turtles are characteristically long, due to their low metabolic rates and consequently low food intakes (Stevens & Hume 2004). For example, mean retention time for Chelonoidis denticulata was estimated by Guzmán & Stevenson (2008) to be of 21 days, and for C. denticualta and C. carbonaria by Strong (2005) to range between 10 to 28 days with a peak at 11-15 days. These observations might suggest that reptiles, especially highly frugivorous chelonians, could play an important role in the dispersal of seeds in time.
About 1% of seeds dispersed by endozoochory in tropical rain forest are attributable to reptiles (Van der Pijl 1969, Arbeláez & Parrado-Rosselli 2005), and saurocory has received little attention in these habitats. Although, several studies demonstrate that lizards and tortoises (Iverson 1985, Strong & Fragoso 2006, Guzmán & Stevenson 2008) are efficient seed dispersers, their role should depend on their population densities that are poorly known. For Chelonoidis denticulata -formerly Geochelone denticulata- available data show densities between 0.1-0.3 ind./ha (Moskovits 1985, Stevenson et al. 2007, Guzmán & Stevenson 2008) and for Chelonoidis carbonaria – formerly Geochelone carbonaria an average density of 0.45 ind./ha (Strong 2005).
The effect of seed dispersers on plant populations depend on the number of seeds manipulated and qualitative factors that might affect seed survival and plant establishment (Schupp 2007). Vertebrate and invertebrate seed predators influence distance- and density- dependent seed mortality (Wright 2002). These predators may act in a distance- or density-responsive manner, depending on the season, plant species or resource availability among other factors, with varying effects on seed mortality (Schupp 2007). For example, pigs were the principal cause of mortality where seeds of the tropical palm Normanbya normanbyi, occurred in high concentrations, whereas insects caused higher mortality rates when seeds were dispersed (Lott et al. 1995). Other studies have shown high seed predation by mammals, but no distance-dependent effect, and low invertebrate seed predation highly correlated to the distance- densitydependent effect (Howe et al. 1985, Wright 2003). A possible explanation for this is that invertebrates or pathogens often complete their relatively short life cycles within the area occupied by a large tree, while mammals do it over much larger areas in the course of an annual cycle (Terborgh & Wright 1994, Pizo 1997, Zipparro & Morellato 2005).
In tropical forests small mammals and insects are the most important seed predators (Notman & Gorchov 2001, Wright 2002, Paine & Beck 2007). In either case, seeds dispersed in time would be advantageous for plant species with seeds vulnerable to density- responsive seed predators. Furthermore, phenological timing differences may affect plant-animal interactions, plant fitness and distribution (Chuine 2010).
Time dispersal has received little attention and its importance, as a density-dependent process remains unstudied. This study addresses the following questions: (1) Are seed removal rates below parent trees lower after the fruiting period has finished than during fruiting period? (2) Do different seed predators have the same mortality effect at different phenophases? (3) If there are differences in mortality rates during and after fruiting, can they be associated to invertebrate or mammalian seed predation? In this study we experimentally test the hypothesis that seeds could minimize predation in time and according to this idea we predicted that seed predation will be higher during the fruiting period than after the fruiting period, affecting seed survival.
Materials and methods
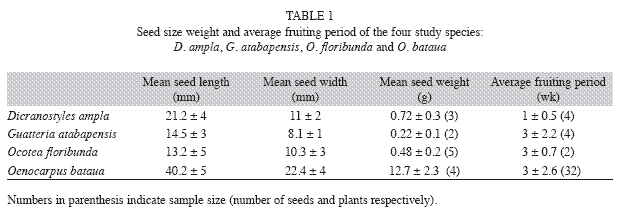
Study site: Experiments were carried out between September and December 2007 in two biological stations at South Western Colombian Amazon forests. We tested two species, Dicranostyles ampla Ducke (Convolvulaceae) and Oenocarpus bataua Mart. (Arecaceae), in Caparú Biological Station (1°04’22" S - 69°31’03" W) (Vargas & Stevenson 2009). We used the same protocol for Guatteria atabapensis D. M. Johnson & N. A. Murray (Annonaceae) and Ocotea floribunda (Sw.) Mez (Lauraceae) in the Zafire Biological Station (4°00’11" S - 069°53’44" W, 115m.a.s.l.) (Castillo & Stevenson 2010). All four plant species produce fleshy endozoochorous fruits and are found in terra firme forest in these forests with continuous and high canopies. The selection of the studied species was based on the availability of fruiting plants, seed size (length>1cm) and adaptations for endozoochorous dispersal (i.e. fleshy parts) (Table 1).
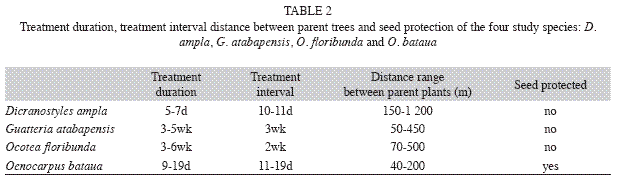
Experimental design: Experiments consisted of two main treatments where seeds were tested for predation below parent trees: (1) during the fruiting period; and (2) several days after the fruiting period had finished. The first treatment simulated no seed dispersal in time, while the second treatment simulated seeds dispersed in time by chelonians. Treatment duration and treatment interval varied among species and among individual plants, however these were similar in both periods (Table 2).
Furthermore, we assessed the potential type of predators by using metal exclosures in half of the plots in both treatments. Exclosures consisted of a metal cage of 50cmx30cmx30cm, with a mesh size of 0.5cm. Plots protected with exclosures allowed only non-vertebrate seed predation, while unprotected plots were exposed to both vertebrate and non-vertebrate seed predators. Seeds placed at the unprotected plots where distributed with the same average distance between them as seeds of the protected plots (ca. 2m). In this experimental design, each tree was used as replicate and the number of parent plants and plots varied according to their availability. For G. atabapensis the experiment was set up under five parent trees, each one with six unprotected plots and six protected plots. D. ampla included four parent plants, each one with six unprotected and six protected plots. O. bataua comprised six parent palms, each with four unprotected and four protected plots. Finally, O. floribunda consisted of four parent trees each with seven unprotected and seven protected plots. Plots were placed randomly under the crown of the parent tree, with ten seeds that were checked daily for disappearance, insect or fungus attack, teeth marks or germination. For the protected plots the fate of seeds was classified into: survived (present, viable or germinated), or predated by non-vertebrates (absent or if present, with evidence of fungus or insect attack). For the unprotected plots, the fate of the seeds was classified in the same way as the protected plots but additionally, seed predation by vertebrates was estimated by subtracting the number of seeds that disappeared in the unprotected plots from the number of seeds that disappeared in the protected plots in each tree, for all species. Mean percentage of seed survival was calculated for each treatment for all species tested. For the post-fruiting treatment, all seeds were replaced and were checked for the same period of time and frequency as seeds of the first treatment, except for O. bataua seeds that were checked once at the end.
Seed survival was determined using logrank survival probability tests (Bland & Altman 2004) in the JMP program (version 3.0), comparing survival curves between treatments. However, it does not test interactions. For O. bataua we conducted a G-test (Pop tools, Excel) for differences between treatments with data on seed predation at the end of the two experimental periods. For species where seed survival differs between periods (during and after), the fate of the seeds was compared in terms of mean seed survival and associated with the type of predator (vertebrate or non-vertebrate).
Results
The principal source of mortality for all species examined was non-vertebrate predation, followed by vertebrate predation (D. ampla: 36%, 15%; G. atabapensis: 16.1%, 6.5%; O. floribunda: 39.7%, 0% and O. bataua: 40%, 0% respectively).
Non-vertebrate seed predation of D. ampla was mainly caused by leaf cutter ants (Atta sp.) while seeds of O. floribunda were repeatedly buried by a medium sized beetle (0.5-1.0cm long), making it difficult to distinguish if seeds were predated or manipulated by invertebrates. Seeds of O. bataua were only predated by a small beetle (<0.3cm long), however some of these seeds were still able to germinate. Missing seeds increased after the fruiting period for D. ampla (8% during fruiting and 22.5% after fruiting) and for G. atabapensis (4% during fruiting and 9% after fruiting).
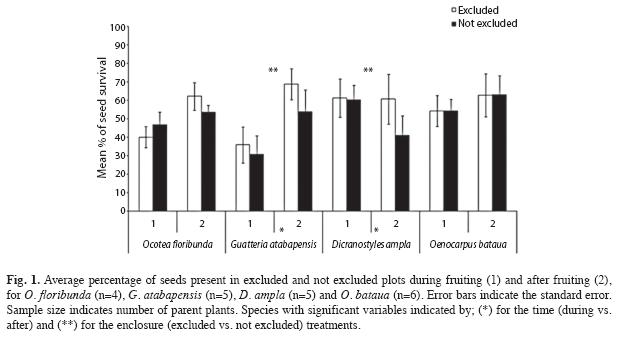
Results on seed survival varied between plant species and treatments. For the time treatment (during and after the fruiting period), we found no statistical differences for O. floribunda and O. bataua (Log-rank test, p=0.64, X2=0.23, n=1120 and G-test, p=0.72, G=0.13, respectively, Fig. 1). Contrary, we found high statistical significance for G. atabapensis and for D. ampla (Log-rank test, p<0.001, X2=14.7, n=1 200 and G-test, p<0.001, X2=18.7, n=960, respectively; Fig. 1). Seed survival was higher during the fruiting period in D. ampla suggesting predator satiation, while seed survival for G. atabapensis was higher after the fruiting period, supporting the time avoidance hypothesis. For D. ampla non-vertebrate seed predation increased with time dispersal; 40.4% (SD=23.3, n=8) of the seeds were predated by non-vertebrates during the fruiting period and 50.6% (SD=32.9, n=8) after the fruiting period. Vertebrate predation also increased with time dispersal; we estimated that 7.9% (SD=7.9, n=4) of the seeds were predated by vertebrates during the fruiting period and 22.8% (SD=15.9, n=4) after de fruiting period. For G. atabapensis, non-vertebrate seed predation decreases with time dispersal; 54.0% (SD=17.7, n=10) of the seeds were predated by non-vertebrates during the fruiting period and 40.2% (SD=21.1, n=10) after the fruiting period; while vertebrate seed predation increased; we estimated that 4.1% (SD=4.4, n=5) of the seeds were predated by vertebrates during the fruiting period and 9.2% (SD=12.4, n=5) after the fruiting period.
For the exclosure treatment (excluded or not-excluded from vertebrate seed predation), seed survival was not statistically significant for O. floribunda and O. bataua (Log-rank test, p=0.7, X2=0.15, n=1120 and G-test, p=0.72, G=0.13, respectively; Fig. 1), while for G. atabapensis and for D. ampla survival was statistically higher in exclosures (Log-rank test, p=0.001, X2=11.6, n=1200 and G-test, p=0.003, X2=8.9, n=960, respectively; Fig. 1).
Discussion
Seeds of most tropical tree species lack dormancy and germinate within few months after dispersal, regardless of environmental conditions (Garwood 1983). Such strategy seems adequate to avoid seed predation, which is a persistent interaction (Crawley 2000) resulting in strong establishment limitation (Paine & Harms 2009). Therefore, many seed dispersal mechanisms that allow higher seed and seedling survival should have a great evolutionary potential.
Seed dispersal by reptiles or saurochory, was initially described as an interaction characterized by fruits with strong smells and predominantly couliflorous plants (van der Pijl 1969). Recently, it has been found that reptiles, such as Chelonoid tortoises are generalist fruit consumers (Moskovits 1985, Guzmán & Stevenson 2008) that eat fruits with a variety of traits (including smelly and odorless fruits). Furthermore, saurochory seems to be prevalent in islands (Olesen & Valido 2003), but saurochory in mainland areas is also more important than previously thought (Traveset 1990, Moll & Jansen 1995). For instance, strong mutualistic relationships between tortoises and plant species have been documented (Rick & Bowman 1961), but we lack evidence that any sort of coevolution has derived in a particular seed dispersal syndrome (Herrera 1985).
The results of this study indicated that predation avoidance in time was advantageous for one of the species tested (G. atabapensis), due to a reduction of seed predation by non-vertebrates after the fruiting period had finished. Seed predation by vertebrates in this study was estimated based on missing seeds, therefore it is possible that missing seeds could have been dispersed instead of being predated. However, removed seeds are highly likely to end up dying in Neotropical forests (Notman & Gorchov 2001, Russo & Augspurger 2004).
Time-dispersal had a negative effect on D. ampla, probably due to predator satiation, phenomenon influencing forest regeneration in many sites (Curran & Webb 2000, Stevenson 2007). Satiation in studies testing seed predation during the fruiting period (early, middle and late), indicate that insect seed predation decreases from the beginning to the end of the fruiting season, while rodent predation tends to increase (Wright 1990, von Allmen et al. 2004). These observations suggest that timedispersal would not be an important process to minimize vertebrate seed predators, but may be advantageous to minimize mortality caused by invertebrates or pathogens. Thus it is strange that in our study site, where seed predation by non-vertebrates in all of the species tested was higher than vertebrate seed predation, only one of the species had an advantage of our timedispersal simulation. Therefore, even though overall quantity of seeds dispersed by tortoise or by other reptiles is low compared to other seed dispersers (i.e. primates and birds), and that the probability of a coevolutionary process evolve with these generalist organisms is low, future studies should be address to better understand survival advantages given by an endozoochory time-dispersal.
Plant phenological differences (e.g. continuous/bursts, and intra-specific synchrony/asynchrony) represent adaptations to biotic and abiotic factors (van Schaik et al. 1993). For example, plant species vulnerable to generalist seed predators should benefit from highly clumped fruiting at supra-annual intervals (Ims 1990). If satiation provides seeds with a survival advantage, plant fitness would be maximized when all plant species synchronize annually for fruiting in order to satiate predators. The best examples of this phenomenon come from South East Asian tropical forests, where plants tend to synchronize massive fruit production to satiate seed predators as a strategy for survival (Herrera et al. 1998, Kelly & Sork 2002). Primary consumers respond to this by a switch in their diet, change in their range use or migration, in order to survive until the next fruiting period. In the tropics, continuous fruiting is rare and evidence supports the hypothesis that species avoid predation by synchronizing their phenological activity (van Schaik et al. 1993). However, in the Neotropics, where most plants depend on animal frugivores, as seed dispersal agents, most plants produce at annual intervals (Stevenson et al. 2008). Therefore, there seems to be an association between more continuous fruit production and large proportion endozoochorous systems, implying that large disperser populations may survive when fruit production relatively continuous, without extended scarcity periods (Terborgh 1986).
Selection pressures favoring seed time dispersal could be similar to those selecting for dormancy in order to reduce risk, escape crowding, and escape sibling competition. For example, in years with unfavorable environmental conditions that increases risk, at unpredictable environments, or when sibling competition is higher locally than elsewhere (Venable & Brown 1988). It is clear that safe sites for plant recruitment are highly unpredictable, because of spatial and temporal variation (Schupp 2007), therefore escape in time is an additional dimension in which seeds may reach temporarily adequate sites for recruitment.
Acknowledgments
We thank Camila Pizano and Nigel Pitman for comments on the manuscript and Santiago Castillo for field assistance. This study was provided financial support by Los Andes University seed grant and the Centro de Investigaciones Ecológicas La Macarena (CIEM).
References
Augspurger, C.K. 1981. Reproductive synchrony of a tropical shrub: experimental studies on effects of pollinators and seed predators on Hybanthus prunifolius (Violaceae). Ecology 62: 775-788. [ Links ]
Arbeláez, M.V. & A. Parrado-Roselli. 2005. Seed dispersal modes of the sandstone plateau vegetation of the middle Caquetá river region, Colombian Amazonia. Biotropica 37: 64-72. [ Links ]
Bagchi, R., T. Swinfield, R.E. Gallery, O.T. Lewis, S. Gripenberg, L. Narayan & R.P. Freckleton. 2010. Testing the Janzen-Connell mechanism: pathogens cause overcompensating density dependence in a tropical tree. Ecol. Lett. 13: 1262-1269. [ Links ]
Bland, J.M. & D.G. Altman. 2004. Statistics notes: The logrank test. BMJ 328: 1073. [ Links ]
Castillo, L.S. & P.R. Stevenson. 2010. Relative Importance of seed-bank and post-disturbance seed dispersal on early gap regeneration in a Colombian Amazon Forest. Biotropica 42: 488-492. [ Links ]
Chuine, I. 2010. Why does phenology drive species distribution? Phil. Trans. R. Soc. B. 365: 3149-3160. [ Links ]
Cohen, D. & S.A. Levin. 1987. The interaction between dispersal and dormancy strategies in varying and heterogeneous environments, p. 110-122. In E. Teramoto & M. Yamaguti (eds.). Mathematical topics in population biology, morphogenesis and neurosciences proceedings. Springer, Berlin, Germany. [ Links ]
Connell, J.H. 1971. On the role of natural enemies in preventing competitive exclusion in some marine animals and rain forests trees, p. 298-312. In P.J. den Boer & G.R Gradwell (eds.). Dynamics of population. Center for Agricultural Publication and Documentation, Wageningen, The Netherlands. [ Links ]
Cousens, R.D., J. Hill, K. French & I.D. Bishop. 2010. Towards a better prediction of seed dispersal by animals. Funct. Ecol. 24: 1163-1170. [ Links ]
Crawley, M.J. 2000. Seed predators and plant population dynamics, p. 167-182. In M. Fenner (ed.). Seeds: the ecology of regeneration in plant communities. CABI Publishing, Oxford, England. [ Links ]
Curran, L.M. & C.O. Webb. 2000. Experimental tests of the spatiotemporal scale of seed predation in mast-fruiting Dipterocarpaceae. Ecol. Monogr. 70: 129-148. [ Links ]
Garwood, N.C. 1983. Seed germination in a seasonal tropical forest in Panama: A community study. Ecol. Monogr. 53: 159-181. [ Links ]
Guzmán, A. 2005. Dispersión de semillas por la tortuga de patas amarillas Geochelone denticulata, en la Amazonía Peruana. M.Sc. Thesis, Universidad de Los Andes, Bogotá, Colombia. [ Links ]
Guzmán, A. & P.R. Stevenson. 2008. Seed dispersal, habitat selection and movement patterns in the Amazonian tortoise Geochelone denticulata. Amphibia-Reptilia 29: 463-472. [ Links ]
Hammond, D.S. & V.K. Brown. 1998. Disturbance, phenology and life-history characteristics: factors influencing distance/density- dependent attack on tropical seeds and seedlings, p. 51-78. In D.M. Newbery, H.H. Prins & N. Brown (eds.). Dynamics of tropical communities. Blackwell Science, London, England. [ Links ]
Herrera, C.M. 1985. Determinants of plant-animal coevolution: the case of mutualistic dispersal of seeds by vertebrates. Oikos 44: 132-141. [ Links ]
Herrera, C.M., P. Jordano, J. Guitián & A. Traveset. 1998. Annual variability in seed production by woody plants and the masting concept: reassessment of principles and relationship to pollination and seed dispersal. Am. Nat. 152: 576-594. [ Links ]
Howe, H.F., E. Schupp & L.C. Westley. 1985. Early consequences of seed dispersal for a neotropical tree (Virola surinamensis). Ecology 66: 781-791. [ Links ]
Hyatt, L.A., M.S. Rosenberg, T.G. Howard, G. Bole, W. Fang, J. Anastasia, K. Brown, R. Grella, K. Hinman, J.P. Kurdziel & J. Gurevitch. 2003. The distance dependence prediction of the Janzen-Connell hypothesis: a meta-analysis. Oikos 103: 590-602. [ Links ]
Imbert, E. 1999. The effects of achene dimorphism on the dispersal in time and space in Crepis sancta (Asteraceae). Can. J. Bot. 77: 508-513. [ Links ]
Ims, R.A. 1990. The ecology and evolution of reproductive synchrony. Trends Ecol. Evol. 5: 135-140. [ Links ]
Iverson, J. 1985. Lizards as seed dispersers? J. Herpetol. 19: 292-293. [ Links ]
Janzen, D.H. 1970. Herbivores and the number of tree species in tropical forests. Am. Nat. 104: 501-528. [ Links ]
Kelly, D. & V.L. Sork. 2002. Mast seeding in perennial plants: why, how, where? Annu. Rev. Ecol. Systemat. 33: 427-447. [ Links ]
Lambert, J.E. 1998. Primate digestion: interactions among anatomy, physiology, and feeding ecology. Evol. Anthropol. 7: 8-20. [ Links ]
Lehouck, V., T. Spanhove, A. Gonsamo, N. Cordeiro & L. Lens. 2009. Spatial and temporal effects on recruitment of an Afromontane forest tree in a threatened fragmented ecosystem. Biol. Conserv. 142: 518-528. [ Links ]
Lott, R.H., G.N. Harrington, A.K. Irvine & S. McIntyre. 1995. Density-dependent seed predation and plant dispersion of the tropical palm Normanbya normanbyi. Biotropica 27: 87-95. [ Links ]
Matthesius, A., H. Chapman & D. Kelly. 2010. Testing for Janzen-Connell effects in a West African montane forest. Biotropica 43: 77-83. [ Links ]
Moll, D. & K.P. Jansen. 1995. Evidence for a role in seed dispersal by two tropical herbivorous turtles. Biotropica 27: 121-127. [ Links ]
Moskovits, D. 1985. The behaviour and ecology of the two Amazon tortoise, Geochelone carbonaria and Geochelone denticulata, in northwestern Brazil. Ph.D. Thesis, Chicago University, Chicago, USA. [ Links ]
Notman, E. & D.L Gorchov. 2001. Variation in postdispersal seed predation in mature Peruvian lowland tropical forest and fallow agricultural sites. Biotropica 33: 621-636. [ Links ]
Olesen, J.M. & A. Valido. 2003. Lizards as pollinators and seed dispersers: an island phenomenon. Trends Ecol. Evol. 18: 177-181. [ Links ]
Paine, T.C.E. & H. Beck. 2007. Seed predation by neotropical rain forest mammals increased diversity in seedling recruitment. Ecology 88: 3076-3087. [ Links ]
Paine, T.C.E. & K.E. Harms. 2009. Quantifying the effects of seed arrival and environmental conditions on tropical seedling community structure. Oecologia 160: 139-150. [ Links ]
Peterman, J.S., A.J.S. Fergus, L.A. Turnbull & B. Schmid. 2008. Janzen-Connell effects are widespread and strong enough to maintain diversity in grasslands. Ecology 89: 2399-2406. [ Links ]
Pizo, M.A. 1997. Seed dispersal and predation in two populations of Cabralea canjerana (Meliaceae) in the Atlantic forest of southeastern Brazil. J. Trop. Ecol. 13: 559-578. [ Links ]
Rick, C.M. & R.I. Bowman. 1961. Galapagos tomatoes and tortoises. Evolution 15: 407-417. [ Links ]
Russo, S.E. & C.K. Augspurger. 2004. Aggregated seed dispersal by spider monkeys limits recruitment to clumped patterns in Virola calophylla. Ecol. Lett. 7: 1058-1067. [ Links ]
Schupp, E.W. 2007. The suitability of a site for seed dispersal is context-dependent, p. 445-462. In A.J. Dennis, E.W. Schupp, R.J. Green & D.A. Westcott (eds.). Seed dispersal: theory and its application in a changing world. CAB International, Wallingford, United Kingdom. [ Links ]
Stevens, C.E. & I.D. Hume. 2004. Comparative physiology of the vertebrate digestive system. Cambridge University, Cambridge, England. [ Links ]
Stevenson, P.R. 2007. A test of the escape and colonization hypothesis for zoochorous tree species in a Western Amazonian forest. Plant. Ecol. 190: 245-258. [ Links ]
Stevenson, P.R., C.A. Borda, A.M. Rojas & M. Álvarez. 2007. Population size, habitat choice and sexual dimorphism of the Amazonian tortoise (Geochelone denticulata) in Tinigua National Park, Colombia. Amphibia-Reptilia 28: 217-226. [ Links ]
Stevenson, P.R., M.C. Castellanos, A.I. Cortés & A. Link. 2008. Flowering patterns in a seasonal tropical lowland forest in Western Amazonia. Biotropica 40: 559-567. [ Links ]
Strong, J.M. 2005. Seed dispersal and the ecological implications of hunting Geochelone carbonaria and Geochelone denticulada in Northwestern Brazil. Master Thesis, State University of New York, USA. [ Links ]
Strong, J.M. & J.M.V. Fragoso. 2006. Seed dispersal by Geochelone carbonaria and Geochelone denticulate in Northwestern Brazil. Biotropica 38: 683-686. [ Links ]
Terborgh, J. 1986. Keystone plant resources in the tropical forest, p. 330-344. In M.E. Soule (ed.). Conservation Biology: The science of scarcity and diversity. Sinauer, Sunderland, Massachusetts, USA. [ Links ]
Terborgh, J.W. & J.S. Wright. 1994. Effect of mammalian herbivores on plant recruitment in two Neotropical forests. Ecology 75: 1829-1833. [ Links ]
Traveset, A. 1990. Ctenosaura similis Gray (Iguanidae) as a seed disperser in a Central American deciduous forest. Am. Midl. Nat. 123: 402-404. [ Links ]
Van der Pijl, L. 1969. Principles of dispersal in higher plants. Springer, Berlin, Germany. [ Links ]
Van Schaik, C.P., J.W. Terborgh & S.J. Wright. 1993. The phenology of tropical forests: adaptative significance and consequences for primary consumers. Annu. Rev. Ecol. Systemat. 24: 353-377. [ Links ]
Vargas I.N. & P.R. Stevenson. 2009. Patrones fenológicos en la Estación Biológica Mosiro Itajura-Caparú: Producción de frutos estimada a partir de transectos fenológicos y trampas de frutos, p. 99-114. In G. Alarcón-Nieto & E. Palacios (eds.). Estación Biológica Mosiro Itajura - Caparú: Biodiversidad en el territorio del Yaigojé-Apaporis. Conservación Internacional Colombia, Bogotá D.C., Colombia. [ Links ]
Venable, D.L. & J.S. Brown. 1988. The selective interactions of dispersal, dormancy, and seed size as adaptations for reducing risk in variable environments. Am. Nat. 13: 360-384. [ Links ]
Von Allmen, C., L.P.C. Morellato & M.A. Pizo. 2004. Seed predation under high seed density condition: the palm Euterpe edulis in the Brazilian Atlantic forest. J. Trop. Ecol. 20: 471-474. [ Links ]
Wright, S.J. 1990. Cumulative satiation of a seed predator over the fruiting season of its hosts. Oikos 58: 272-276. [ Links ]
Wright, S.J. 2002. Plant diversity in tropical forests: a review of mechanisms of species coexistence. Oecologia 130: 1-14. [ Links ]
Wright, S.J. 2003. The myriad effects of hunting for vertebrates and plants in tropical forests. Perspect. Plant Ecol. Evol. Systemat. 6: 73-86. [ Links ]
Zipparro, V.B. & L.P.C. Morellato. 2005. Seed predation of Virola bicuhyba (Schott) Warb. (Myristicaceae) in the Atlantic forest of south-eastern Brazil. Rev. Bras. Bot. 28: 515-522. [ Links ]
Correspondencia a: Adriana Guzmán. Fundación Malpelo y Otros Ecosistemas Marinos, Carrera 7 No. 32-33, piso 27. Bogotá, Colombia; aguzman@fundacionmalpelo.org
Pablo R. Stevenson. Laboratorio de Ecología de Bosques Tropicales y Primatología, Universidad de Los Andes, Carrera 1 No. 18-10. Bogotá, Colombia; pstevens@uniandes.edu.co
Received 06-XII-2010. Corrected 07-IV-2011. Accepted 04-V-2011.


{kind=link}
{kind=link}
{kind=link}