










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.4 San José Dec. 2011
Desarrollo post-embrionario, fecundidad y consumo de alimento de Dichroplus exilis (Orthoptera: Acrididae) bajo condiciones controladas
Christian Bardi1, Yanina Mariottini1, Maria L. De Wysiecki1 & Carlos E. Lange1,2
1. Centro de Estudios Parasitológicos y de Vectores (CEPAVE) (CCT La Plata - CONICET - UNLP), Calle 2 Nº 584, CP 1900, La Plata, Buenos Aires, Argentina; bardi_c@yahoo.com.ar, ymariottini@cepave.edu.ar, mlw@cepave.edu.ar2. Comisión de Investigaciones Científicas (CIC) de la provincia de Buenos Aires; carlosl@cepave.edu.ar
Dirección para correspondencia
Abstract
Postembryonic development, fecundity and food consumption of Dichroplus exilis (Orthoptera: Acrididae) under controlled conditions. Dichroplus exilis is a widely distributed species in Southern South America. Although there have been reports of D. exilis as an agricultural pest, some recent observations suggest that the damage attributed to D. elongatus may actually have been caused by D. exilis. This study was conducted to determine the postembryonic life cycle stages, fertility and food consumption of this species under controlled conditions (30°C, 14L-10D, 40% RH).Individuals employed belong to the laboratory-hatched first generation (F1), from adults (n=64, ♀=28, ♂=36) collected in natural grasslands near Rafaela, Santa Fe province in North- Eastern Argentina. Three cohorts of 16, 17 and 20 individuals were monitored independently in acetate tubes on a daily basis, until death of the last insect. Average fecundity was 381.84, 38.54 eggs per female. Egg-pod incubation time was 14.4, 1.08 days and six nymphal instars were recorded. Nymphal development time was 41.38, 0.71 days (I=8.73, 0.20; II=6.38, 0.24; III=5.64, 0.33; IV=7.15; 0.43; V=9.76, 0.54; IV=7.85, 0.95). The recorded food consumption was 9.89, 1.08 (mg/ind/day) for nymphs IV, 18.04, 0.73 (mg/ind/day) for nymphs V-IV, 16.76, 1.06 (mg/ind/day) for pre-reproductive males, 28.09, 1.81 (mg/ind/day) for pre-reproductive females, 7.71, 0.91 (mg/ind/day) for reproductive males and 13.06, 0.71 (mg/ind/day) for reproductive females, while the average adult food consumption, regardless of sex and reproductive status, was 16.41, 4.32mg/day. Average food consumption of adult females was 17.47, 1.15mg, and was significantly higher than that of males (10.83, 0.91mg). Data obtained in this study showed that D. exilis exhibits at least some of the biological attributes needed to configure an actual or potential agricultural pest, albeit not yet recognized as such. Field monitoring of grasshopper communities in areas where damage by D. exilis is suspected is envisaged in order to determine its possible status as a pest. Rev. Biol. Trop. 59 (4): 1579-1587. Epub 2011 December 01.
Key words: Agricultural pest, Argentina, consumption tests, Dichroplus elongatus, grasshopper, nymphal instars.
Resumen
La Subfamilia Melanoplinae tiene una relevancia central dentro de la acridiofauna Argentina. Varias especies suelen ser numéricamente dominantes en las comunidades de acridios del país y algunas constituyen serias plagas. El objetivo de este trabajo fue conocer el ciclo de vida postembrionario, la fecundidad y supervivencia de las hembras y el consumo de alimento de Dichruplus exilis bajo condiciones controladas (30°C; 40%HR y 14L-10O). Para ello se realizó el seguimiento de tres cohortes de 16, 17 y 20 individuos. El tiempo de incubación fue de 14.4±1.08 días. La duración del desarrollo ninfal fue 41.38; 0.71 días (I 8.73; 0.20, II 6.38; 0.24, III 5.64; 0.33, IV 7.15; 0.43, V 9.76; 0.54; IV 7.85; 0.95); la fecundidad promedio fue de 381.84, 38.54huevos/hembra. El consumo registrado fue de 9.89; 1.08 (mg/ind./día) para las ninfas de IV, 18.04; 0.73 (mg/ind./día) para las de V y IV, mientras que el consumo promedio de los adultos, sin diferenciar sexo y estado reproductivo, fue de 16.41; 4.32mg/día. Estos parámetros, al ser comparados con los de otras especies afines, indican que D. exilis exhibe atributos biológicos que condicen con los de otros melanoplinos considerados plaga.
Palabras clave: Acridio, Argentina, Dichroplus elongatus, estadio ninfal, plaga agrícola, pruebas de consumo.
Dichroplus exilis (Giglio-Tos) es un melanoplino de amplia distribución geográfica en la región sur de América del Sur; se encuentra en Uruguay, Paraguay, Sur de Brasil (Rio Grande do Sul) y en el norte y gran parte del centro de la Argentina (Carbonell et al. 2006). D. exilis pertenece al "grupo elongatus" que incluye especies con fuerte similitud en la anatomía externa y los patrones de coloración (Ronderos et al. 1968). Es una especie poco estudiada, multivoltina, de régimen alimentario polífago y con diapausa embrionaria facultativa (Barrera & Turk 1977, Turk & Barrera 1979). Si bien hasta el presente no se han mencionado perjuicios económicos para el agro por parte de esta especie, es posible que daños a cultivos y pasturas debidos a su accionar hayan sido erróneamente atribuidos a Dichroplus elongatus (Giglio-Tos), especie simpátrica con la cual puede ser fácilmente confundida cuando la identificación se basa sólo en caracteres morfológicos externos (Ronderos et al. 1968, COPR 1982).
La subfamilia Melanoplinae constituye el grupo más diverso de acridios en Argentina, con un total de 60 especies conocidas (Carbonell et al. 2006), muchas de las cuales suelen ser numéricamente dominantes en las comunidades de acridios del país, por ejemplo Cigliano et al. (2000), en el Oeste de la Región Pampeana registraron que la subfamilia Melanoplinae fue la más abundante, representando el 68.8% del total de individuos recolectados, mientras que De Wysiecki et al. (2004), determinaron que los melanoplinos representaron el 85% de los individuos en el Sudeste de la misma región. Dentro de esta familia, se encuentran algunas de las especies más perjudiciales (Dichroplus elongatus, Dichroplus maculipennis, Dichroplus pratensis, Dichroplus vitattus, Baeacris punctulatus, Ronderosia bergi y Scotussa lemniscata), para el agro argentino. (COPR 1982, Cigliano et al. 2002, Lange et al. 2005, Carbonell et al. 2006). No obstante, aún son limitadas las contribuciones que brindan información sobre aspectos biológicos de estas especies. Considerando esta situación, el objetivo del presente trabajo fue estudiar el ciclo de vida, la fecundidad y supervivencia de las hembras y el consumo de alimento de D. exilis bajo condiciones controladas.
Materiales y métodos
Parámetros biológicos: Los individuos empleados en este estudio correspondieron a la primera generación obtenida en bioterio a partir de ejemplares adultos (n=64, ♀=28, ♂=36 originalmente recolectados en pastizales naturales cercanos a la localidad de Rafaela, provincia de Santa Fe (31°15’25.94» S - 61°29’14.48» W), en el noreste de Argentina. Los acridios fueron criados en jaulas de aluminio de (20x20x30cm) y alimentados con hojas frescas de lechuga, diferentes especies de gramíneas, repollo y salvado de trigo, y mantenidos bajo condiciones controladas (30ºC, 14L:10O, 40% HR). La dieta y las condiciones ambientales fueron similares a las utilizadas en la mayoría de los centros de cría de acridios a nivel mundial (Henry 1985, Hinks & Erlandson 1994).
Cada jaula fue provista de un sitio de oviposición para las hembras, que consistió en un recipiente cilíndrico de 8cm de diámetro y 10cm de profundidad, conteniendo como sustrato, partes iguales de arena y tierra tamizada y esterilizada. La termorregulación y la cópula fueron estimuladas con lámparas de 75W suspendidas a unos 15cm sobre las jaulas. Al carecer D. exilis de diapausa embrionaria obligatoria (Barrera & Turk 1977, Turk & Barrera 1979), las posturas fueron inmediatamente incubadas a 30°C hasta su nacimiento.
Para conocer el desarrollo post-embrionario se realizó el seguimiento diario de tres cohortes de 16, 17 y 20 individuos recién emergidos, hasta la muerte del último insecto, considerándose una cohorte a un conjunto de individuos de la misma edad. Cada cohorte fue mantenida de manera independiente en tubos transparentes de acetato con extremos de alambre tejido (Henry 1985) hasta alcanzar el estado adulto. Se registró la supervivencia diaria, la duración total del desarrollo postembrionario, el número y la duración de los estadios ninfales, la duración total del ciclo ninfal y la longevidad y supervivencia de las hembras adultas. Una vez en estado adulto se formaron 25 parejas para estimar la fecundidad. Cada pareja (1♂, 1♀) fue colocada en una jaula de aluminio (12x12x15cm) provista de un recipiente (sustrato) para la oviposición. Los sustratos fueron tamizados semanalmente para conocer el número promedio de posturas por hembra. Cincuenta posturas fueron abiertas bajo microscopio simple, con el fin de conocer el número de huevos por postura. Con estos datos se calculó el número de posturas por hembra a lo largo de toda su vida. El porcentaje de eclosión se estimó a partir de la incubación de siete posturas sin abrir y el posterior recuento de los huevos sin eclosionar presentes en las mismas.
Se construyeron las curvas de supervivencia (lx) y de fecundidad específica por edades (mx) utilizando la porción hembra de la cohorte, y el intervalo de clases de edad sucesivas fue de una semana. A partir de estos datos se calculó la tasa neta de reproducción (R0)
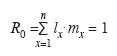
Los parámetros estimados fueron consumo promedio diario y total por individuo de cada estadio y, pérdida total promedio por individuo (consumo+material caído). El consumo de un individuo en la edad adulta se estimó promediando el consumo de los estados prereproductivo y reproductivo, multiplicado por la longevidad promedio. Se estimó la biomasa promedio de los individuos adultos de ambos sexos, expresada como peso seco, mediante la ponderación directa de los individuos, y se estimó la tasa de consumo relativo diaria (mg/mg.individuo/día) para ambos sexos.
Los datos se analizaron mediante la prueba de Kruskal-Wallis seguido de Mann-Whitney. Todos los análisis se realizaron con XLSTAT 7.5.3. (Addinsoft 2005).
Resultados
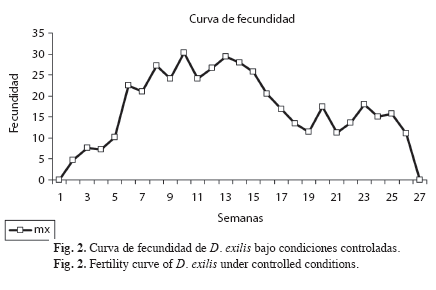
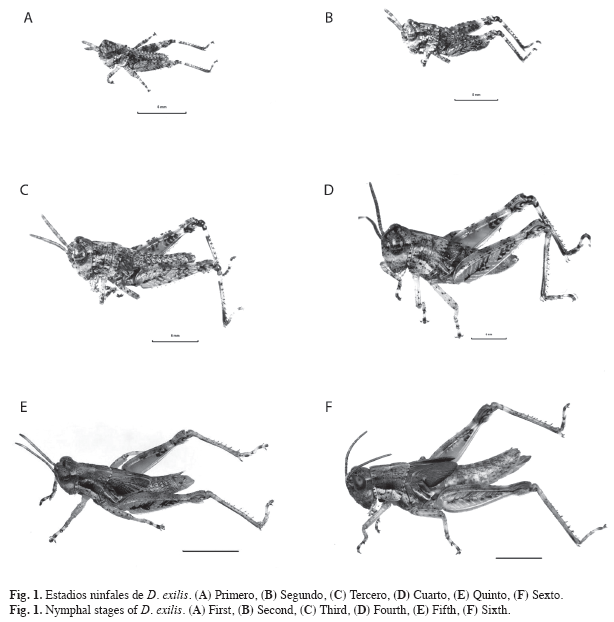
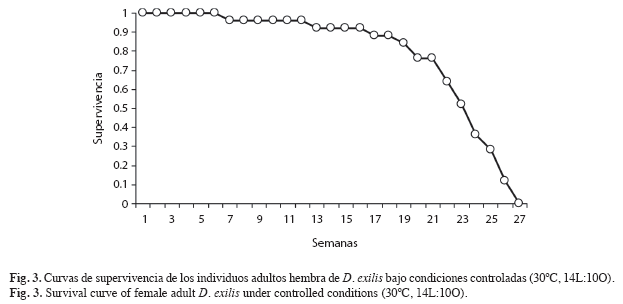
Parámetros biológicos: Se registró un total de seis estadios ninfales para ambos sexos (Fig. 1). La duración del estado ninfal fue de 41.38; Error Estándar 0.71; ámbito (34-51) días (Cuadro 1). Las primeras cópulas, manifestación externa de la madurez sexual en el macho (Richard & Davies 1983), se observaron a partir del quinto día de haber alcanzado el estado adulto. La primera oviposición, indicio de la madurez sexual en la hembra (Michel & Terán 2006), se observó a los once días de iniciado el estado adulto. El tiempo promedio de incubación fue de 14.4; 1.08; (9-19) días. El número promedio de posturas por hembra fue 19.96; 1.94; (0-37). El número promedio de huevos por postura fue 15.6; 1.03; (7-26). La fecundidad promedio (número de huevos/ hembra) fue 381.84, 38.54; (0-652) (Fig. 2). La tasa de oviposición fue de 2.36; 0.27 (huevos/hembra/día). El porcentaje de eclosión fue de 94.33%. La tasa neta de reproducción (R0) fue de 176.78. La longevidad promedio de las hembras adultas fue de 158.64; 6.66; (48-194) días (Fig. 3). La duración promedio de las cohortes desde el nacimiento hasta la muerte fue de 201.56; 6.53 días.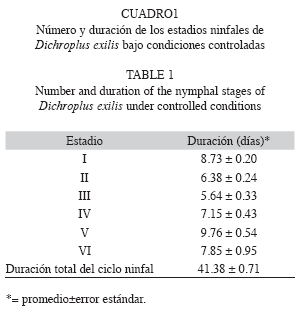
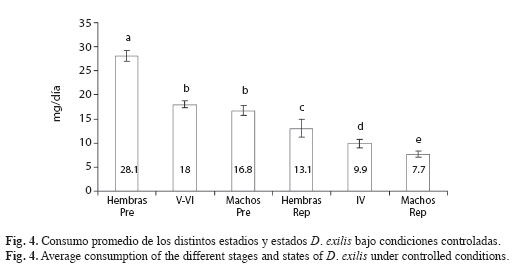
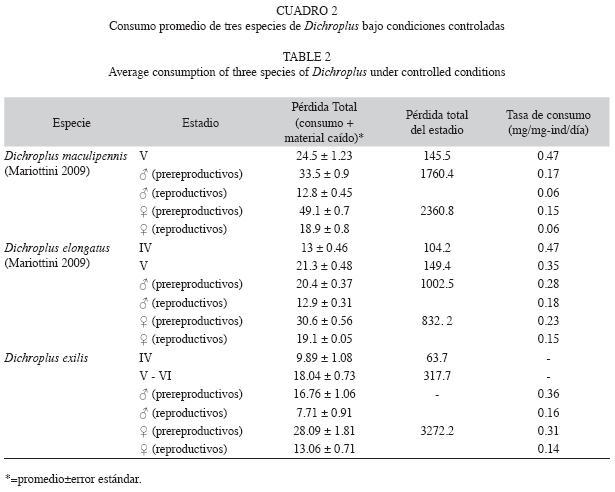
Consumo: D. exilis no dejó caer comida al alimentarse, por lo que se consideró que la pérdida total fue igual al consumo. El contenido de agua de las raciones testigo (factor de corrección) fue de 80.03; 0.69%. El consumo promedio fue de 9.89; 1.08 (mg/ind./día) para las ninfas de IV estadio, 18.04; 0.73 (mg/ind./día) para las ninfas de V-VI estadio, 16.76; 1.06 (mg/ind./día) para los machos pre-reproductivos,28.09; 1.81 (mg/ind./día) para las hembras pre-reproductivas, 7.71; 0.91 (mg/ind./día) para los machos reproductivos y 13.06; 0.71 (mg/ ind./día), para las hembras reproductivas. El consumo de los individuos en los distintos estadios de desarrollo fue significativamente diferente (Kruskal Wallis, p<0.0001). Se observó un aumento en el consumo desde el IV estadio ninfal hasta los adultos pre-reproductivos, descendiendo en los adultos reproductivos (Fig. 4). El consumo de las hembras pre-reproductivas (♀Pre) fue significativamente superior al consumo de los individuos en IV (p<0.001), V-VI (p<0.0001), ♂ Pre (p<0.0001), ♀ Rep (p<0.0001) y ♂ Rep (p<0.0001). El consumo de los machos pre-reproductivos (♂ Pre) fue significativamente superior al consumo de los individuos en IV (p<0.001), ♂ Rep (p<0.0001), ♀ Rep (p=0.003). El consumo de los individuos de V-VI fue significativamente superior al consumo de los individuos en IV (p<0.001), ♂ Rep (p<0.0001) y ♀ Rep (p<0.0001). El consumo de las hembras reproductivas (♀ Rep) fue significativamente superior al consumo de los individuos en IV (p=0.043) y ♂ Rep (p<0.0001). El consumo de los machos reproductivos (♂ Rep) fue significativamente inferior al de los individuos en IV (p=0.026). No se registraron diferencias significativas entre el consumo de los individuos en V-VI y ♂ Pre (p=0.255). El consumo promedio de los adultos de D. exilis, sin diferenciar sexo y estado reproductivo, fue de 16.41; 4.32mg/día. El consumo promedio de una hembra en estado adulto fue de 17.47; 1.15mg/día, y resultó significativamente mayor que el de los machos, 10.83; 0.91mg/día (Kruskal-Wallis, p<0.001). El consumo relativo en los adultos fue ligeramente mayor en los machos 0.263 (mg/ mg.individuo/día) que en las hembras 0.227 (mg/mg.individuo/día). La biomasa promedio de D. exilis fue de 90.74; 4.67mg para las hembras y 46.60; 1.32mg para los machos.
Discusión
La Subfamilia Melanoplinae incluye el mayor número de especies de acridios en América, con aproximadamente 330 especies conocidas en América del Norte, unas 235 para 116.77; 32.60 para Dichroplus schulzi y Mariottini et al. (2008) y Mariottini (2009) registraron valores de 46.03; 6.84 y de 86.06; 13.03 para Beacris (Dichroplus) pseudopunctulatus y Dichroplus maculipennis, respectivamente. Dichroplus exilis, en cambio, presentó un promedio de 381.84, 38.54 huevos por hembra, con un máximo de 652huevos/hembra. Estos valores resultan más cercanos a los observados para Melanoplus que para otros Dichroplus.
Será necesario realizar prospecciones en las zonas donde se presume que D. exilis es la especie realmente causante de daños y no D. elongatus, tales como la provincia de Santa Fe y el noroeste de la región Pampeana (noroeste de Buenos Aires, Norte de La Pampa, Sur de Córdoba), para dilucidar su eventual estatus como plaga.
Adicionalmente, dado que D. exilis carece de diapausa embrionaria obligatoria y tiene en laboratorio alta longevidad y fecundidad puede ser considerada como una especie apta para el desarrollo y mantenimiento de colonias de cría, las cuales son de utilidad para la realización de estudios experimentales, en particular con agentes de control biológico (Lange 2002, 2003).
Agradecimientos
Esta investigación fue financiada en parte por el "Fondo para la Investigación Científica y Tecnológica (FONCYT, proyecto PICT 2006/679)" y "Comisión de Investigaciones Científicas (CIC) de la provincia de Buenos Aires".
Referencias
Addinsoft. 2005. XLSTAT Pro version 7.5.3 (disponible en línea: http://www.xlstat.com/en/home). [ Links ]
Amédégnato, C., W. Chapco & G. Litzenberger. 2003. Out of South America? Additional evidence for a southern origin of melanopline grasshoppers. Mol. Phylogenet. Evol. 29: 115-119. [ Links ]
Barrera, M. & S.Z. Turk. 1977. Acridios del NOA II: Contribución al conocimiento de huevos, desove y hábitos de postura de algunas especies de tucuras (Orthoptera: Acrididae) de la provincia de Tucumán. Acta Zool. Lilloana 32: 167-188. [ Links ]
Carbonell, C.S., M.M. Cigliano & C.E. Lange. 2006. Acridomorph (Orthoptera) species of Argentina and Uruguay. CD-ROM. Publications on Orthopteran Diversity, The Orthopterists Society at the "Museo de La Plata", Argentina. [ Links ]
Chapco, W., G. Litzenberger & W.R. Kuperus. 2001. A molecular biogeographic analysis of the relationship between North American melanopline grasshoppers and their Eurasian and South American relatives. Mol. Phylogenet. Evol. 18: 460-466 [ Links ]
Cigliano, M.M., M.L. De WysieckI & C.E. Lange. 2000. Grasshopper (Orthoptera, Acrididae) species diversity in the Pampas, Argentina. Divers. Distrib. 6: 81-91. [ Links ]
Cigliano, M.M., S. Torrusio & M.L. De Wysiecki. 2002. Grasshopper (Orthoptera: Acrididoidea) community composition and temporal variation in The Pampas, Argentina. J. Orthop. Res. 11: 215-221. [ Links ]
Centre for Overseas Pest Research (COPR). 1982. The locust and grasshopper agricultural manual, COPR, London. [ Links ]
De Wysiecki, M.L. & N. Sánchez. 1992. Dieta y remoción de forraje de Dichroplus pratensis (Orthoptera, Acrididae) en un pastizal de la provincia de La Pampa, Argentina. Ecol. Austral. 2: 19-27. [ Links ]
De Wysiecki, M.L., M.M. Cigliano & C.E. Lange. 1997. Fecundidad y longevidad de adultos de Dichroplus elongatus (Orthoptera: Acrididae) bajo condiciones controladas. Rev. Soc. Entomol. Argent. 56: 101-104. [ Links ]
De Wysiecki, M.L., S. Torrusio & M.M. Cigliano. 2004. Caracterización de las comunidades de acridios del partido de Benito Juárez, sudeste de la provincia de Bs. As, Argentina. Rev. Soc. Entomol. Argent. 63: 87-96. [ Links ]
Fielding, D.J. & M.A. Brusven. 1995. Ecological correlates between rangeland grasshopper (Orthoptera: Acrididae) and plant communities of southern Idaho. Environ. Entomol. 24: 1432-1441. [ Links ]
Gangwere, S.K. & R.A. Ronderos. 1978. The current status of New World acridology. Interciencia 3: 16-19. [ Links ]
Henry, J.E. 1985. Melanoplus spp., p. 451-464. In P. Singh & R.F. Moore (eds.). Handbook of insect rearing, Vol. 1. Elsevier, Amsterdam. [ Links ]
Hewitt, G.B. & R.J. Onsager. 1983. Control of grasshopper on rangeland in the United States, a perspective. J. Range Manage. 3: 202-207. [ Links ]
Hinks, C.F. & M.A. Erlandson. 1994. Rearing grasshoppers and locust: review, rationale and update. J. Orthop. Res. 3: 1-10. [ Links ]
Joern, A. & S.B. Gaines. 1990 Populations dynamics and regulation in grasshoppers, p. 415-482. In R.F. Chapman & A. Joern (eds.). Biology of grasshoppers. John Wiley and Sons, New York, EEUU. [ Links ]
Lange, C.E. 2002. La amebiasis debilitativa de los ortópteros y su potencial para el control biológico de acridios (Orthoptera: Acridoidea) en la Argentina. Rev. Invest. Agropecu., Ser. 2. 31: 25-38. [ Links ]
Lange, C.E. 2003. Niveles de esporulación experimentales y naturales del agente de biocontrol Nosema locustae (Protozoa. Microspora) en especies de tucuras y langostas (Orthoptera: Acridoidea) de la Argentina. Rev. Soc. Entomol. Argent. 62: 15-22. [ Links ]
Lange, C.E., M.M. Cigliano & M.L. De Wysiecki. 2005. Los acridoideos de importancia económica en la Argentina, p. 93-135. In L. Barrientos Lozano & P. Almaguer (eds.). Manejo integrado de la langosta centroamericana y acridoideos plaga en América Latina. Instituto Tecnológico de Ciudad Victoria, Tamaulipas, México. [ Links ]
Michel, A.A. & H.R. Terán. 2006. Cría en cautiverio de Baeacris punctulatus (Orthoptera,Acrididae, Melanoplinae). Acta Zool. Lilloana 50: 131-134. [ Links ]
Mariottini, Y., M.L. De Wysiecki & C.E. Lange. 2008. Fecundidad y desarrollo postembrionario de Baeacris pseudopunctulatus Ronderos (Orthoptera: Acrididae: Melanoplinae) bajo condiciones controladas. Rev. Soc. Entomol. Argent. 67: 135-138. [ Links ]
Mariottini, Y. 2009. Biología y ecología de Acridios (Orthoptera: Acridoidea). Tesis Doctorado, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina. [ Links ]
O’Neill, K.M., B.E. Olson, M.G. Rolston, R. Wallander, D.P. Larson & C.E. Seibert. 2003. Effects of livestock grazing on rangeland grasshopper (Orthoptera:Acrididae) abundance. Agr. Ecosyst. Environ. 97: 51-64. [ Links ]
Otte, D. 1995. Orthoptera species File 4. Grasshoppers Acridomorpha C. Orthopterist´s Society and The Academy of Natural Sciences of Philadelphia, Philadelphia, EEUU. [ Links ]
Pfadt, R.E. 1994. Migratory grasshopper Melanoplus sanguinipes (Fabricius). Wyoming Agricultural Experiment Station Bulletin 912, Species Fact Sheet. (disponible en línea: www.sdvc.uwyo.edu/grasshopper/mesa.htm). [ Links ]
Pfadt, R.E. 2002. Field guide to common western grasshoppers. University of Wyoming, Laramie, EEUU. [ Links ]
Putnam, L.G. 1962. The damage potential of some grasshoppers on the Native Grassland of British Columbia. Can. J. Plant. Sci. 42: 596-601. [ Links ]
Putnam, L.G. 1963. The progress of nymphal development in pest grasshoppers Acrididae) of western Canada. Can. Entomol. 95: 1210-1216. [ Links ]
Ragonese, E.A. 1985. Forrajeras nativas. Reunión Técnica en Honor al Ing. Hermán Serrano. INTA EEA Pergamino. [ Links ]
Ronderos, R.A., C.S. Carbonell & A. Mesa. 1968. Revisión de las especies del género Dichroplus Stal, del grupo elongatus (Orthoptera, Acrididae, Catantopinae). Rev. Mus. La Plata, Secc. Zool. 10: 271-325. [ Links ]
Sánchez, N.E. & M.L. De Wysiecki. 1990. Quantitative evaluation of feeding activity of the grasshopper Dichroplus pratensis (Orthoptera: Acrididae) in a natural grassland of La Pampa, Argentina. Environ. Entomol. 19: 1392-1395. [ Links ]
Sánchez, N.E., P. Pessacq, M. Theiller & G. Almandoz. 2001. Aspectos biológicos y demografía de Dichroplus schulzi Bruner (Orthoptera: Acrididae), en laboratorio. Rev. Soc. Entomol. Argent. 60: 73-78. [ Links ]
Turk, S.Z. & M. Barrera. 1979. Acridios del NOA. III. Estudio bio-ecológico sobre siete especies del género Dichroplus Stal (Orthoptera, Acrididae). Acta Zool.Lilloana 35: 785-805. [ Links ]
Vickery, V.R. 1989. The biogeography of Canadian Grylloptera and Orthoptera. Can. Entomol. 121: 389-424. [ Links ]
Correspondencia a: Christian Bardi, Yanina Mariottini & Maria L. De Wysiecki. Centro de Estudios Parasitológicos y de Vectores (CEPAVE) (CCT La Plata - CONICET - UNLP), Calle 2 Nº 584, CP 1900, La Plata, Buenos Aires, Argentina; bardi_c@yahoo.com.ar, ymariottini@cepave.edu.ar, mlw@cepave.edu.ar
Carlos E. Lange. Centro de Estudios Parasitológicos y de Vectores (CEPAVE) (CCT La Plata - CONICET - UNLP), Calle 2 Nº 584, CP 1900, La Plata, Buenos Aires, Argentina / Comisión de Investigaciones Científicas (CIC) de la provincia de Buenos Aires; carlosl@cepave.edu.ar
Recibido 25-X-2010. Corregido 20-II-2011. Aceptado 29-III-2011.


{kind=link}
{kind=link}
{kind=link}