










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.2 San José Jun. 2011
Effect of sucrose and methyl jasmonate on biomass and anthocyaninproduction in cell suspension culture of Melastoma malabathricum (Melastomaceae)
Koay Suan See, Arvind Bhatt & Chan Lai Keng
Plant Tissue and Cell Culture Laboratory, School of Biological Sciences, University Sains Malaysia, 11800 Penang, Malaysia; suansee9@hotmail.com, arvin_bhatt@rediffmail.com, lkchan@usm.my, merrilynchan@gmail.com
Dirección de correspondencia
Abstract
Melastoma malabathricum, belongs to the Melastomaceae family, is an important medicinal plant widely distributed from Madagascar to Australia, that is used in traditional remedies for the treatment of variousailments. Besides its medicinal properties, it has been identified as a potential source of anthocyanin production.The present study was carried out to investigate the effect of sucrose and methyl jasmonate and feeding time oncell biomass yield and anthocyanin production in cell suspension culture of M. malabathricum. Addition of differentconcentrations of sucrose into the cell culture of M. malabathricum influenced cell biomass and pigment accumulation. The addition of methyl jasmonate was found to have no effect on cell biomass but the presence of higher amount (12.5-50mg/L) had caused a reduction in anthocyanin production and accumulation. MS medium supplemented with 30g/L sucrose and 3.5 mg/L of MeJA added on cero day and 3rd day produced high fresh cell mass at the end of nine days of culture but did not support the production of anthocyanins. However, cells cultured in the medium supplemented with 45g/L sucrose without MeJA showed the highest pigment content (0.69±0.22Cv/g-FCM). The cells cultured in MS medium supplemented with 30 g/L sucrose with 3.5mg/L MeJA added on the 3rd and 6th day of culture, showed the lowest pigment content (0.37-0.40Cv/g-FCM). This study indicated that MeJA was not necessary but sucrose was needed for the enhancement of cell growth and anthocyanin production in M. malabathricum cell cultures. Rev. Biol. Trop. 59 (2): 597-606. Epub 2011 June 01.
Key words: Melastoma malabathricum, cell suspension culture, sucrose, methyl jasmonate, anthocyanins.
Resumen
Melastoma malabathricum pertenece a la familia de las melastomáceas, es una planta medicinal importante ampliamente distribuida desde Madagascar hasta Australia, que se utiliza en remedios tradicionales para el tratamiento de diversas dolencias. Además de sus propiedades medicinales, se ha identificado como una fuente potencial de producción de antocianinas. En esta investigación se estudió el efecto de la sucrosa, el metil jasmonato y el tiempo de ingestión en la producción de biomasa de las células y la producción de antocianinas, en el cultivo de células en suspensión de M. malabathricum. La adición de diferentes concentraciones de sucrosa al cultivo de células de M. malabathricum influencia la biomasa de las células y la acumulación de pigmento. La adición de metil jasmonato no tuvo ningún efecto sobre la biomasa celular, pero la presencia de una cantidad más alta (12.5-50mg/L) causó una reducción en la producción y acumulación de antocianinas. El medio MS complementado con sucrosa 30g/L y 3.5mg/L de MeJA en el día cero y el tercer día produjo una gran masa de células frescas al final de los nueve días de cultivo pero no se pudo mantener la producción de antocianinas. Sin embargo, las células cultivadas en el medio complementado con 45g/L de sucrosa sin MeJA mostró el mayor contenido de pigmento (0.69±0.22cv/g-fcm). Las células cultivadas en el medio MS complementado con 30 g/L de sucrosa y con 3.5 mg/l MeJA en el tercer y sexto día de cultivo, mostró el menor contenido de pigmentos (0.37-0.40cv/g-fcm). Este estudio indicó que MeJA no era necesario pero la sucrosa sí se necesitaba para mejorar el crecimiento celular y la producción de antocianinas en cultivos de células de M. malabathricum.
Palabras clave: Melastoma malabathricum, suspensión de cultivo celular, sucrosa, metil jasmonato, antocianinas.
Melastoma malabathricum L. (Melastomataceae) is one of the important medicinal plants widely distributed throughout Malaysia. Different plant parts of M. malabathricum are used in traditional remedies for the treatment of various ailments. The crude extract has been used in the treatment of diarrhea, postpartum treatment, hemorrhoids, to relieve toothache and also applied externally on inflamed wounds (Ahmad & Raji 1990, Burkill 1996). Some metabolites isolated from this plant have also been reported to have antiviral and cytotoxic activities, anti-helmethic and anti-spasmodic action, anti-nociceptive, anti-inflammatory and antipyretic properties, anti-oxidant and anti-cancer activities and anti-hypertensive activity (Osman et al. 2000, Lohezic-Le et al. 2002, Cheng et al. 2006, Susanti et al. 2006,Zakaria et al. 2006).
Generally, most of the plant species only produce small quantity of secondary metamolites. Cell suspension culture technique has been used as the alternative for the production of secondary metabolites including plant pigments. Elicitation has been shown to be the most efficient strategy that direct to the enhancement in anthocyanin production in plant cell cultures (Zhang & Furusaki 1999). The addition of methyl jasmonate (MeJA) or jasmonic acid (JA) in the culture medium as an elicitor enhanced the anthocyanin production of Tulipa gesneriana and Vaccinium pahalae (Saniewski et al. 1998, Fang et al. 1999). Methyl jasmonate (MJ) has successfully used as an elicitor in other plant species for enhancing the production of secondary metabolites in the cell cultures (Aoyagi et al. 2001, Kim et al. 2004, Thanh et al. 2005). Similarly, the manipulation in the components of the culture medium (e.g. carbon source, nitrogen and phosphate) also found to be effective for the production of secondary metabolites (Trejo-Tapia et al. 2001). Mori & Sakurai (1994) reported the enhancement of anthocyanin accumulation and cell growth in strawberry cell suspension cultures with the supplement of glucose, sucrose and fructose in the culture medium.
M. malabathricum has also been identified as a potential source for anthocyanin production (Janna et al. 2006). Anthocyanins are well known alternatives to synthetic food colours/ dyes and beneficial to health with potential physiological benefits (Mazza & Brouillard 1990, Downham & Collins 2000). However, until now there is no report available regarding the elicitor effect on cell biomass yield and anthocyanin production in cell suspension culture of M. malabathricum. Therefore, the present study was undertaken with an aim to investigate the effect of sucrose and methyl jasmonate acted as elicitors of cell biomass yield and anthocyanin production in cell suspension culture of M. malabathricum.
Material and Methods
Establishment of cell suspension cultures of M. malabathricum: The cell suspension culture of M. malabathricum was prepared by inoculating 0.75g (fresh weight) of nine-day old cells into liquid MS (Murashige & Skoog 1962) medium supplemented with 0.25mg/L BA and 0.5mg/L nAA, the cell proliferation medium (Chan et al. 2008). Sterile stainless steel sieve with 850μm pore size was used to filter the initial established cell cultures to obtain homogenous cell suspension culture. Individual cells or small cell aggregates that passed through the sieve (with diameter <850μm) were subsequently filtered through Whatman no. 1 filter paper using an air suction pump (Aspirator 3A-S-Eyela, Raikakikai Co. Ltd, Tokyo) to separate the cells from the medium. The cell cultures were placed on an orbital shaker (new Brunswick Scientific, Edison, NJ, USA) at 120rpm in a culture room maintained at 24±2oC under continuous illumination with cool white fluorescent tubes at a light intensity of 6.3-11.9mmol photons m-2sec-1. After nine days, the cells were harvested and used as plant materials for subsequent studies.
Elicitation with sucrose and methyl jasmonate: A cell biomass of 0.5g obtained from the cell suspension culture which was subcultured every nine days in liquid MS medium supplemented with 0.25mg/L BA+0.5mg/L NAA and exposed to continuous illumination was inoculated into each of the 100ml Erlenmeyer flasks containing 20ml liquid MS medium supplemented with different concentrations of sucrose (0, 15, 30, 45, 60, 75 and 90g/L). Eight replicates were used for each sucrose concentration treatment.
The same cell mass of 0.5g obtained through similar method was utilized to test the elicitation effect of methyl jasmonate (MeJA). Methyl jasmonate solution was prepared by dissolving 250mg of MeJA (Phyto Technology Laboratories) in ethanol: water (12: 13) to give a 50g/L stock solution. The MeJA solutions and the solvent were sterilized using a 0.22µm membrane filter under sterile condition. Different amount of 50g/L MeJA solution was treated into the cell proliferation medium to obtain final MeJA concentrations of 2, 5, 12.5, 20 and 50mg/L. Control cultures received 20µl of sterile filtered solvent. Eight replicates were used for each MeJA concentration treatment.
The cells were harvested at the end of nine days. Cell aggregates and the liquid medium were separated as previously mentioned and the fresh cell mass was determined after the cells were harvested. The pigment content and pigment production was determined from the harvested cells as stated below.
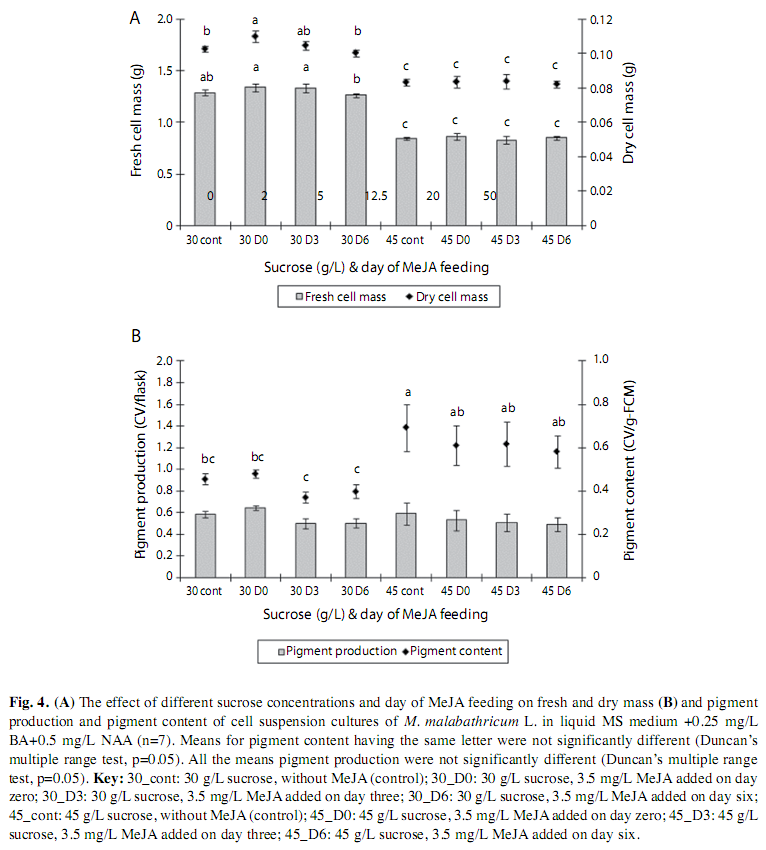
Effect of sucrose and day of elicitation of methyl jasmonate: Cell mass of 0.5g was inoculated into two different culture media: (i) liquid MS medium supplemented with 0.25mg/L BA+0.5mg/L nAA+30g/L sucrose (the normal sucrose content used in MS medium) and (ii) liquid MS medium supplemented with 0.25mg/L BA+0.5mg/L nAA+45g/L sucrose (best sucrose concentration that stimulated the highest pigment production). MeJA concentration of 3.5mg/L was added into the medium at day zero, day three and day six of culture. The addition of MeJA solution at the beginning of cell culture (0 day) was used as control. The cultures were maintained under continuous light with intensity of 6.3-11.9mmol photons m-2sec-1 and the cells were harvested at the end of nine days of culture. The cell mass, pigment content and production were then determined.
Quantification of pigment content and pigment production: The total anthocyanins, represented by pigment content and pigment production of each culture, was quantified based on its colour value (Cv)/g-DCM (dried cell mass) or Cv/flask, respectively. Eighty milligrams of freeze-dried cells were placed into a specimen bottle and 10ml of acidified methanol (methanol+1% HCl (Hydrogen chloride), v/v), the best extraction solvent, was added to the cells and stirred; the mixtures were left at 10oC for 24 hours. Optical absorbance (or optical density, OD) of the supernatant of each sample was determined at 535nm wavelength using the Pharmacia novaspec II Rapid spectrophotometer against the blank which consisted of the solvent. Pigment content, Cv/g-DCM, was adapted from Mori et al. (1993) as follow:
Pigment content = 0.1 x OD535 x dilution factor
(Cv/g-DCM)
Pigment production, Cv/flask, was calculated by multiplying the pigment content with the respective mean dried cell mass obtained at the end of each culture.
Data collection and analysis: Seven to eight replicates were used for each of the treatment study and each individual study was carried out using complete randomized design (CRD). The fresh cell biomass, the pigment content and pigment production was determined for each sample after nine days of culture. The data were analyzed using a one-way ANOVA followed by Duncan’s multiple range test for mean comparison at p≤0.05.
Results
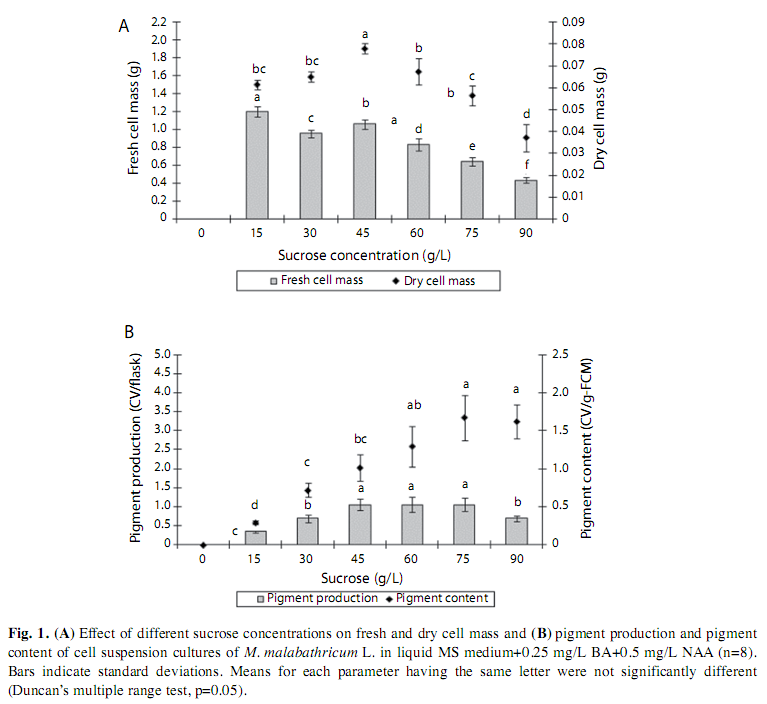
Effect of Sucrose: Different added concentration of sucrose influenced the cell biomass and anthocyanin accumulation in cell suspension cultures of M. malabathricum. However, control (without sucrose) caused cell death. Medium supplemented with 15g/L sucrose induced the highest fresh cell weight, (1.20±0.11g) which was significantly different from the cells cultured in medium supplemented with 45g/L sucrose. On the contrary, significantly higher dried cell weight was obtained from the cells cultured in the medium supplemented with 45g/L sucrose as compared to the cells cultured in the medium supplemented with 15g/L (Fig. 1A). However, when the sucrose level was further increased (60g/L or higher), there was a decreasing trend in cell growth.
The cells cultured in medium supplemented with 15g/L sucrose exhibited the lowest pigment content (0.29CV/g-FCM) and pigment production (0.35CV/flask). The cells cultured in medium supplemented with high sucrose concentration (90g/L) produced very high pigment content of 1.62±0.44CV/g-FCM. However, its cell biomass yield was very low (0.44±0.07g) and this had caused a low pigment production (0.69CV/flask). The amount of pigment produced in the cells cultured in medium supplemented with 45, 60 and 75g/L sucrose achieved high pigment production of 0.88-1.48CV/flask, each of which was significantly different from the other sets of cell cultures. Among these three treatment, culture supplemented with 45g/L sucrose showed the highest growth index (1.12), highest dried cell mass with fairly high pigment production and content, hence the addition of 45g/L sucrose into the cell culture of M. malabathricum could be used for enhancement of pigmentation (Fig. 1B).
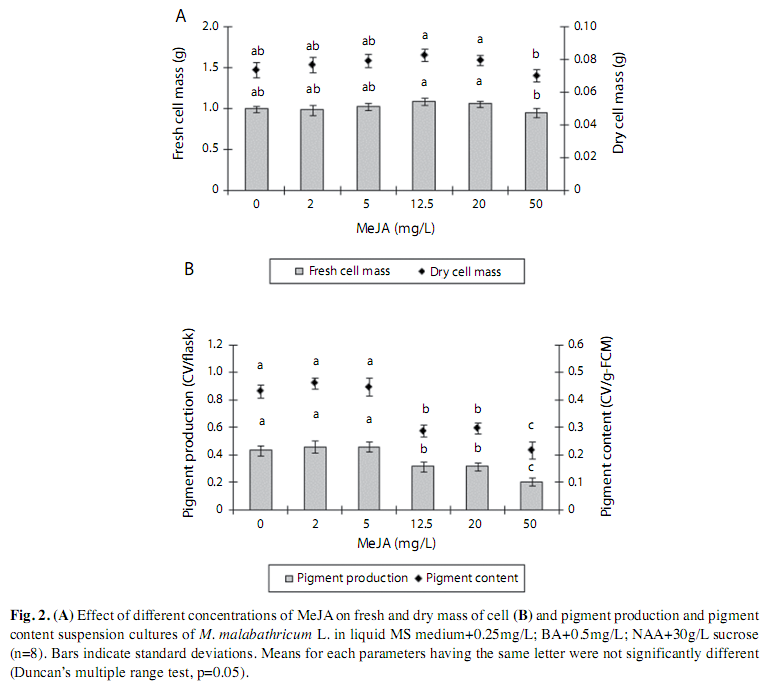
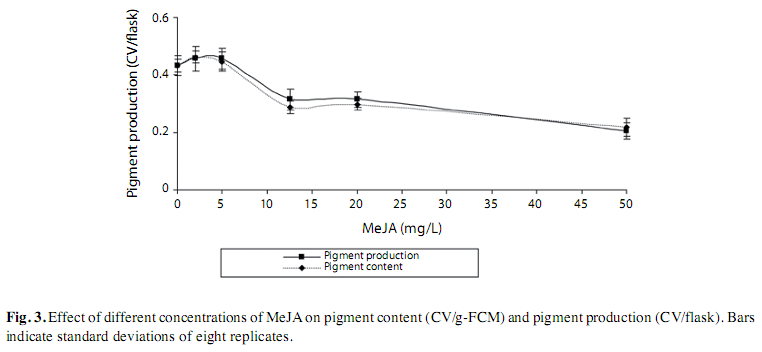
Elicitation with Methyl Jasmonate: The results showed that addition of MeJA at concentrations between 0 and 50mg/L at day zero did not influence the cell biomass at the end of nine days of culture (Fig. 2A). The control culture and the cells cultured in MS medium containing of low concentration of MeJA (2-5mg/L) showed significantly higher pigment content and pigment production as compared to those cultures incubated in MS medium supplemented with higher concentrations of MeJA (12.5-50mg/L). The highest concentration of MeJA (50mg/L) showed a significantly low anthocyanin accumulation (Fig. 2B). Additional result obtained indicated that a concentration of 3.5mg/L MeJA was found to be the best concentration that could induce the highest pigment production and was used in subsequent study (Fig. 3).
Effect of sucrose concentration and day of elicitation of methyl jasmonate: The results clearly showed that culture medium supplemented with 30g/L sucrose and 3.5mg/L of MeJA, added on different day did not affect the fresh and dried cell mass at the end of nine days of culture, and this was not significantly different from the cultures grown in absence of MeJA. Cell biomass was also not influenced by the addition of MeJA at different days into the culture media with 45g/L sucrose. However, the amount of cell biomass produced was significantly lower than using the culture medium containing 30g/L sucrose (Fig. 4A). Cell proliferation medium containing 30g/L or 45g/L sucrose with or without the addition of 3.5mg/L MeJA did not affect pigment production. However, cells cultured in the medium supplemented with 45g/L sucrose without MeJA showed the highest pigment content (0.69±0.22Cv/g-FCM) (Fig. 4B).
Discussion
Effect of Sucrose: The high sucrose content reduced water content in the cultured cells and the supply of sucrose as low as 15g/L, was found to be sufficient for cell growth and provide the energy carbon source for cell metabolism. However, higher sucrose level (60g/L or higher) showed the decreasing trend in cell growth of M. malabathricum. Similar trend was reported by Sato et al. (1996) that supplementing the culture medium of strawberry cell suspension culture with >0.09M sucrose resulted in a decrease in cell growth and attributed this to the inhibition of nutrient uptake due to an increase in the osmotic potential or the high viscosity of the medium. Zhong & Yoshida (1995), however, stated that in a medium in which all nutrients were present in excess, an increase in sugar concentration could result in a proportional increase in cell biomass. Experimenting on Perilla frutescens cell cultures, they reported that growth rates increased with increase in initial sucrose concentration (15, 30, 45 and 60g/L) in the medium for both cultures with initial inocula of 15 and 50 g-wet cells/L. Our result thus confirmed that higher concentration of sucrose supplemented into the culture medium reduce cell biomass due to the increase of osmotic potential which subsequently reduce the uptake of nutrient.
The cells culture supplemented with 15g/L sucrose exhibited the lowest pigment content and pigment production. However, the culture supplemented with 90g/L sucrose concentration produced high pigment content. On the other hand, this concentration (90g/L sucrose) reduces the cell biomass and pigment production. It is possible that the high level of sucrose could be related to osmotic stress conditions which caused cell death as explained by Rudge and Morris (1996) and hence lower anthocyanin production in M. malabathricum cell cultures.
The cell culture medium supplemented with 45, 60 and 75g/L sucrose show high pigment production. Therefore, we recommend that sucrose content in the culture medium should not be exceeded 75g/L. Zhong & Yoshida (1995) found that for cell suspension culture of Perilla frutescens, the medium that produced higher cell densities had higher sucrose levels as compared to the medium that produced higher pigment accumulation. This observation was the reverse for cell suspension culture of M. malabathricum, whereby the medium that enhanced pigment production had higher sucrose levels compared to the medium that produced the highest biomass production. A relatively higher concentration of sucrose was also reported to be favorable for rosmarinic acid production in Anchusa officinalis cell culture (De-Eknambul & Ellis 1985), the ajmalicine, serpentine, and tryptamine in Catharanthus roseus cell cultures (Merillon et al. 1984), the carotenoid production by carrot cells (Yun et al. 1990), the betacyanin accumulation in suspension cultures of Phytolacca americana as well as anthocyanin production by grape cells (Do & Cormier 1991), and Camptotheca acuminate cells (Pasqua et al. 2005). Product formation could be attributed to a certain level of osmotic stress caused by higher initial sucrose concentration (Zhong & Yoshida 1995). The positive effects of osmotic potential, provoked by high sucrose concentrations in medium, on the accumulation of anthocyanins was demonstrated in cell cultures of Vitis vinifera, where an increase in osmotic potential of the medium from -0.5 to 0.9MPa resulted in a significant increase in accumulation of anthocyanins in pigmented cells (Do & Cormier 1990).
Elicitation with methyl jasmonate: Addition of different concentrations of MeJA showed different trends in pigment content and pigment production. Lower concentrations of MeJA (2-5mg/L) showed significantly higher pigment content and pigment production as compared to the higher concentrations of MeJA (12.5-50mg/L). Our results were in consistent with Fang et al. (1999) who reported that addition of MeJA influenced the anthocyanin accumulation depending on its concentrations in Vaccinium pahalae. Similarly, addition of 200μM MeJA considerably increased the ginsenoside content in Panax ginseng and the increasing concentration (above 200μM) declined the accumulation of ginsenosides (Thanh et al. 2005). It was reported that the increased pigment accumulation caused by MeJA in Alkanna tinctoria was not due to increased biomass growth but to stimulation of the cells to its synthesis (Urbanek et al. 1996). For M. malabathricum cell culture, since the addition of MeJA did not increase the cell biomass but pigment production, it did suggest that MeJA stimulated the synthesis of anthocyanin production via induction of particular enzymes that catalyzes the synthesis of anthocyanin. As reported by Mizukami et al. (1993), Jasmonic acid and its derivatives were involved in a part of the signal transduction pathway that induced particular enzymes catalyzing biochemical reactions for the synthesis of secondary metabolites.
Effect of Sucrose concentration and day of elicitation of Methyl Jasmonate: Culture medium supplemented with 30g/L, 45g/L sucrose and 3.5mg/L of MeJA or without MeJA added on different day did not affect the fresh, dried cell mass and pigment production. However, cells cultured in the medium supplemented with 45g/L sucrose without MeJA showed the highest pigment content indicating that MeJA was not necessary for pigment production in cell culture of M. malabathricum. But the presence of sucrose was needed for both cell biomass and production of pigments. Our earlier result did show that when MeJA was added into the culture medium it did affect the production of pigment, but only very slightly. Hence when in combination of sucrose, the effect of MeJA in pigment production became negligible. That could also be accounted for the reason that the different feeding time did not influence the production and content of pigments. These findings were on contrary with Zhang et al. (2002) who found that addition of jasmonic acid at different feeding time influenced the anthocyanin accumulation in cell suspension cultures of Vitis vinifera.
Acknowledgments
We thank Universiti Sains Malaysia forresearch funding and facilities.
References
Ahmad, F. & H. Raji. 1990. Medicinal plants of the Community in Sabah. Universiti Kebangsaan, Malaysia. [ Links ]
Aoyagi, H., Y. Kobayashi, K. Yamada, M. Yokoyama, K. Kusakar & H. Tanaka. 2001. Efficient production of saikosaponins in Bupleurum falcatum root fragments combined with signal transducers. Appl. Microbiol. Biotechnol. 57: 482-488. [ Links ]
Burkill, I.H. 1996. A dictionary of the economic products of the Malaya Peninsula. Kuala Lumpur, Singapore. [ Links ]
Chan, L.K., S.S. Koay, P.H. Low & P.L. Boey. 2008. Effect of plant growth regulators and subculture frequency on callus culture and the establishment of Melastoma malabathricum cell suspension culture for the production of pigments. Biotechnology 7: 678-685. [ Links ]
Cheng, H., L.J. Yu, Q.Y. Hu, S.C. Chen & Y.P. Sun. 2006. Establishment of callus and cell suspension cultures of Corydalis saxicola Bunting, a rare medicinal plant. Z. naturforsch. C. Biosci. 61: 251-256. [ Links ]
De-Eknambul,W. & B.E. Ellis. 1985. Effects of macronutrients on growth and rosmarinic acid formation in cell suspension cultures of Anchusa officinalis. Plant. Cell. Rep. 4: 46-49. [ Links ]
Do, C.B. & F. Cormier. 1990. Accumulation of anthocyanin enhanced by a high osmotic potential in grape (Vitis vinifera L.) cell suspension. Plant. Cell. Rep. 9: 143-146. [ Links ]
Do, C.B. & F. Cormier. 1991. Effects of low nitrate and high sugar concentrations on anthocyanin content and composition of grape (Vitis vinifera L.) cell suspension. Plant Cell. Rep. 9: 500-504. [ Links ]
Downham, A. & P. Collins. 2000. Colouring our food in the last and next millennium. Int. J. Food. Sci. Technol. 35: 5-22. [ Links ]
Fang, Y., M.A.L. Smith & M.F. Pépin. 1999. Effects of exogenous methyl jasmonate in elicited anthocyanin producing cell cultures of ohelo (Vaccinium pahalae). In vitro Cell Dev. Biol. Plant. 35: 106-113. [ Links ]
Janna, O.A., A. Khairul, M. Maziah & Y. Mohd. 2006. Flower pigment analysis of Melastoma malabathricum. Afr. J. Biotechnol. 5: 170-174. [ Links ]
Kim, Y.S., E.J. Hahn, H.N. Murthy & K.Y. Paek. 2004. Adventitious root growth and ginsenoside accumulation in Panax ginseng cultures as affected by methyl jasmonate. Biotechnol. Lett. 26: 1619-1622. [ Links ]
Kubinsky, E. & G. Ifju. 1972. Influence of steaming on the properties of red oak. Part I. Structural and chemical changes. Wood Science 6: 87-94. [ Links ]
Lohezic-Le, F., A. Devehat, C. Bakhtiar, M.A. Bezivin & J. Boustie. 2002. Antiviral and cytotoxic activities of some indonesian plants. Fitoterapia 73: 400-405. [ Links ]
Mazza, G. & R. Brouillard. 1990. The mechanism of copigmentation of anthocyanins in aqueous solutions. Phytochemistry 29: 1097-1102. [ Links ]
Merillon, J.M., M. Rideau & J.C. Chenieux. 1984. Influence of sucrose on levels of ajmalicine, serpentine and tryptamine in Carharanthus roseus cells in vitro. Planta Med. 50: 497-501. [ Links ]
Mori, T., M. Sakurai, J. Shigeta, K. Yoshida & T. Kondo. 1993. Formation of anthocyanins from cells cultured from different parts of strawberry plants. J. Food. Sci. 58: 788-792. [ Links ]
Mori, T. & M. Sakurai. 1994. Production of anthocyanin from strawberry cell suspension cultures; effects of sugar and nitrogen. J. Food. Sci. 59: 588-593. [ Links ]
Mizukami, H., Y. Tabira & B.E. Ellis. 1993. Methyl jasmonate-induced rosmarinic acid biosynthesis in Lithospermum erythrorhizon cell suspension cultures. Plant. Cell. Rep. 12: 706-709. [ Links ]
Murashige, T. & F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plantarum 15: 473-497. [ Links ]
Osman, K., H.I. Faridah, Z.A. Ibrahim & A. Latif. 2000. A guide book to medicinal plants of Perlis. Jabatan Perhutanan negeri Perlis, Kangar, Perlis, Malaysia. [ Links ]
Pasqua, G., B. Monacelli, I. Mulinacci, S. Rinaldi, C. Giaccherini, M. Innocenti & F.F. Vinceri. 2005. The effect of growth regulators and sucrose on anthocyanin production in Camptotheca acuminate cell cultures. Plant Physiol. Biochem. 43: 293-298. [ Links ]
Rudge, K. & P. Morris. 1996. The effects of osmotic stress on growth an alkaloid accumulation of Catharanthus roseus, p 75-81. In P.A.H. Morris, A. Scragg, M. Stafford & W. Fowler (eds.). Secondary metabolism in plant cell cultures. Cambridge University, Cambridge, England. [ Links ]
Saniewski, M., A. Miszczak, L. Kawa-Misczak, E. Wegrzynowicz-Lesiak, K. Miyamoto & J. Ueda. 1998. Effects of methyl jasmonate on anthocyanin accumulation, ethylene production, and CO2 evolution in uncooled and cooled tulip bulbs. J. Plant. Growth Regul. 17: 33-37. [ Links ]
Sato, K., M. Nakayama & J. Shigeta. 1996. Culturing conditions affecting the production of anthocyanin in suspended cell cultures of strawberry. Plant Sci. 113: 91-98. [ Links ]
Susanti, D., H.S. Sirat, F. Ahmad, R.M. Ali, N. Aimi & M. Kitajima. 2006. Antioxidant and cytotoxic flavonoids from the flowers of Melastoma malabathricum L. Food. Chem. 103: 710-716. [ Links ]
Thanh, N.T., H.N. Murthy, K.W. Yu, E.J. Hahn & K.Y. Paek. 2005. Methyl jasmonate elicitation enhanced synthesis of ginsenoside by cell suspension cultures of Panax ginseng in 5-l balloon type bubble bioreactors. Appl. Microbiol. Biotechnol. 67: 197-201. [ Links ]
Trejo-Tapia, G., A. Jiménez-Aparicio, M. Rodríguez-Monroy, A. De Jesús-Sánchez & G. Gutiérrez-López. 2001. Influence of cobalt and other microelements on the production of betalains and the growth of suspension cultures of Beta vulgaris L. Plant Cell Tissue Organ Cult. 67: 19-23. [ Links ]
Urbanek, H., K. Bergier, M. Saniewski & J. Patykowski. 1996. Effect of jasmonates and exogenous polysaccharides on production of alkannin pigments in suspension cultures of Alkanna tinctoria. Plant Cell Rep. 15: 637-641. [ Links ]
Yun, J.W., J.H. Kim & Y.J. Yoo. 1990. Optimizations of carotenoid biosynthesis by controlling sucrose concentration. Biotechnol. Lett. 12: 905-910. [ Links ]
Zakaria, Z.A., R.N. Raden Mohd Nor, G. Hanan Kumar, Z.D. Abdul Ghani, M.R. Sulaiman, G. Rathna Devi, A.M. Mat Jais, M.N. Somchi & C.A. Fatimah. 2006. Antinociceptive, anti-inflammatory and antipyretic properties of M. malabathricum leaves aqueous extract in experimental animals. Can. J. Physiol. Pharmacol. 84: 1291-1299. [ Links ]
Zhang, W. & S. Furusaki. 1999. Production of anthocyanins by plant cell cultures. Biotechnol. Bioproc. Eng. 4: 231-252. [ Links ]
Zhang, W., C. Curtin, M. Kikuchi & C. Franco. 2002. Integration of jasmonic acid and light irradiation for enhancement of anthocyanin biosynthesis in Vitis vinifera suspension cultures. Plant Sci. 162: 459-468. [ Links ]
Zhong, J.J. & T. Yoshida. 1995. High-density cultivation of Perilla frutescens cell suspensions for anthocyanin production: Effects of sucrose concentration and inoculum size. Enzym. Microb. Tech. 17: 1073-1079. [ Links ]
Correspondencia a: Koay Suan See, Arvind Bhatt & Chan Lai Keng: Plant Tissue and Cell Culture Laboratory, School of Biological Sciences, University Sains Malaysia, 11800 Penang, Malaysia; suansee9@hotmail.com, arvin_bhatt@rediffmail.com, lkchan@usm.my, merrilynchan@gmail.com
Received 21-VII-2010. Corrected 20-X-2010. Accepted 22-XI-2010.


{kind=link}
{kind=link}
{kind=link}
{kind=link}