










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.58 suppl.3 San José Oct. 2010
Restoration and monitoring of a vessel grounding on a shallow reef in the Florida Keys
Joe Schittone
NOAA 1305 E. West Hwy, SSMC4, NMS/6, 11th Floor, Silver Spring, Maryland. USA 20910;joe.schittone@noaa.gov
Abstract
Key words: coral restoration, vessel grounding, coral monitoring, coral density, coral biodiversity, coral sizeclass frequency distribution.
Resumen
Besides the restoration itself, the Act also provides that damages are to be collected to enable monitoring of the recovery of a restoration. The monitoring program tracks patterns of biological recovery and determines the success of restoration measures of the site over time. In this instance in order to evaluate success, reference habitat adjacent to the restoration site was concurrently monitored to compare the condition of restored and "natural" coral reef areas.
Reef Injury: On October 25, 1989, the M/V Alec Owen Maitland, a 47m oil field supply vessel, ran aground in a reef coral community south of Carysfort Reef, in 2-3m of water. Additional injury occurred as the result of initial attempts to "power off" the reef by putting the propellers in reverse, and then revving up the vessels´ engines. The grounding totally destroyed 681m2 of living corals and partially destroyed 930m2 of coral reef framework (NOAA undated).
The grounding of the ship and subsequent attempts to free it resulted in significant injury to the reef substrate and resident marine organisms. Efforts by the vessel to extricate itself caused "blowholes" (gouges caused by high-revving propellers) in the reef´s surface. Approximately 79% of the total stony coral cover, as well as numerous sponges and sea fans at the site, were destroyed by the grounding and/or removal attempts. The injuries ranged from superficial scraping of the reef surface and toppling of large coral heads, to complete crushing of coral and severe cracking of the reef framework structure.
The company responsible for the grounding and NOAA agreed to a settlement in December 1991. Before a restoration could be implemented however, a very large hurricane, Hurricane Andrew, severely impacted the Maitland site. The storm passed less than 100km to the north of the site, with winds exceeding 200kph. After the hurricane´s passage, it was found that the blowholes had been "excavated," i.e., expanded and merged together (CSA 1993).
NOAA believed the hurricane´s effects exacerbated the grounding injuries and that additional storm-generated injuries would occur in the future absent restoration efforts. Thus, restoration was undertaken in 1995. Activities were planned by Office of National Marine Sanctuaries headquarters, and Florida Keys Sanctuary staff. The NOAA personnel collaborated with marine engineers from the firm of Olsen Associates, Inc.; work was implemented by Team Land and Development, Inc., along with Sanctuary staff.
Materials and methods
Reef Restoration: The objectives of the Maitland site restoration were to fill in the blowhole and to stabilize the damaged reef framework. Engineering design for the site was particularly challenging because of the scope of the damage, the site´s shallow-water, high energy environment, and the proximity to sensitive resources. Structural repairs to the site included the placement of 40 pre-cast concrete and limestone boulder units to fill in the grounding crater, filling approximately 800m2 in total. After placement, they were sealed with approximately 45m3 of underwater pumped concrete. Before setting, the concrete was embedded with locally quarried limestone rocks.The units were designed in six sizes to accommodate the crater´s complex and varying geometry. Each unit weighed approximately 9.5-10tons above water. The units were designed to withstand wave and current forces anticipated in a once-in-25-year storm event. The structure has withstood the passage of several close hurricanes (Georges in 1998 and at least seven more recent storms). Overall, the restoration was intended to "re-create a stable foundation which closely emulates the adjacent natural seabed and which would foster future recruitment of the local biota" (Bodge 1996).
Restoration Monitoring: Between August 5 and 10, 2004, the Maitland restoration site was monitored using SCUBA. Another monitoring event occurred on June 7 and July 26, 2005 and the latest monitoring visit occurred between July 27 and September 14, 2007. Between the August 2004 and the June 2005 monitoring evernts, the eyes of three powerful hurricanes passed within 250km of the restoration site. These were Charley in late August, and Jeanne and Frances in September 2004. Each sustained winds approximating 175-195kph at the time of closest approach to the restoration site.
In the 2005 season, hurricane Dennis made its closest approach in July, at similar distances and wind speeds as noted for the 2004 storms. In addition, in August and subsequently, hurricanes Katrina, Rita, and Wilma passed within 175km of the site. At the time of closest approach, Katrina and Rita had winds of about 130kph. Wilma had winds of approximately 205kph, but was over land (SW Florida) at the time. No monitoring of the site was conducted between the 2005 and the 2007 monitoring, and thus the possible confounding effects of these hurricanes, if any, are speculative. However, no visually perceptible damage of the restoration structures was observed.
Field Methods: In 2007, within both the restoration and adjacent Reference sampling area, 21 one m2 quadrats were surveyed for variables of interest as described in the Biological Classifications section. Within each survey area, a random number generator corresponded to a digital grid of uniquely identified 1 m2 cells overlaid on the grounding site map. Transect lines were used from landmarks to determine cell locations in the field as best as possible.
Data Analysis: Regarding the density analyses: for the Gorgonians, square root transformation was necessary to enable t tests; for the Millepora and Scleractinians, no transformations were needed. All three Orders evidenced sufficient homogeneity of variance that no corrections were applied.
For all years, common biodiversity indices were calculated for the Scleractinian populations. Additionally, size-class frequency distributions are shown for Porites astreoides, the only species with a sample size sufficient for such calculations.
Additionally, data on substrate attachment in the restoration area was collected to permit Chi-square (χ2) "Goodness of Fit" Test between actual and expected settlement occurrences. Due to the low proportion of Millepora, the test was conducted with Yates´ continuity correction.
Inter-annual comparisons are made for Scleractinians and Gorgonians. For Scleractinians, no transformations were necessary; analysis proceeded by way of a single-factor ANOVA, followed by Bonferroni´s Multiple Comparison Test. For the Gorgonians, normality could not be achieved after attempted transformation; analysis was by way of a Kruskal-Wallis Test, followed by Dunn´s Multiple Comparison Test.
Results
Density: Scleractinian densities in the Restored area greatly exceeded those in the Reference area. For the three Orders surveyed in 2007, the data yielded the densities shown in Fig. 1. For the Gorgonians, the statistical test used (see Data Analysis) indicated that the difference between the Restoration and the Reference populations were very significantly different (p=0.0015). The same test yielded non-significant results (p=0.8875) for Millepora. For Scleractinians, the statistical test utilized resulted in highly significant differences between the two areas (p<0.0001).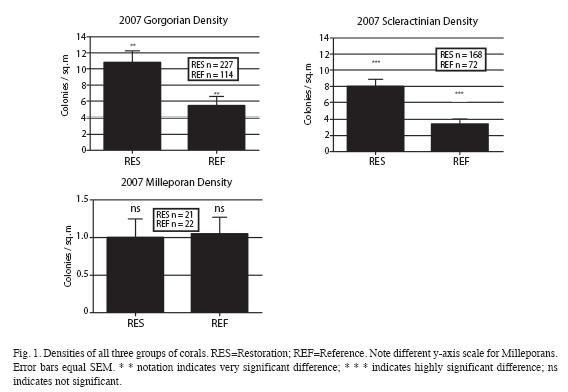
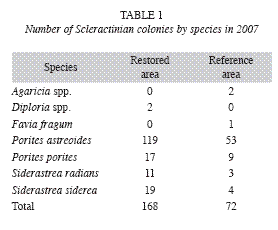
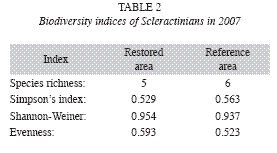
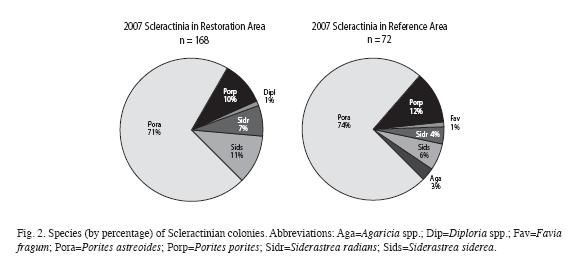
Biodiversity: Table 1 and Fig. 2 show a comparison of the biodiversity of Scleractinian colonies among the Restored and Reference sampling areas within the Maitland restoration site. Table 2 lists the results of a number of standard biodiversity indices performed for the Scleractinian colony populations.
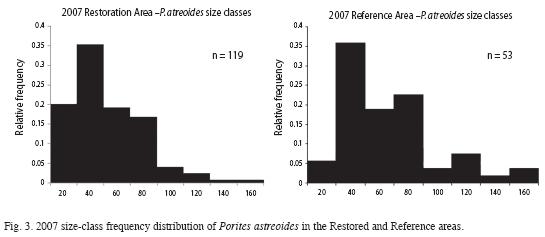
Size Distribution: In 2007 size-class frequency distributions were ascertained for the only coral with sufficient numbers to make such calculations meaningful, Porites astreoides. The graphs depicting the Restored and Reference population distributions are shown in Fig. 3.
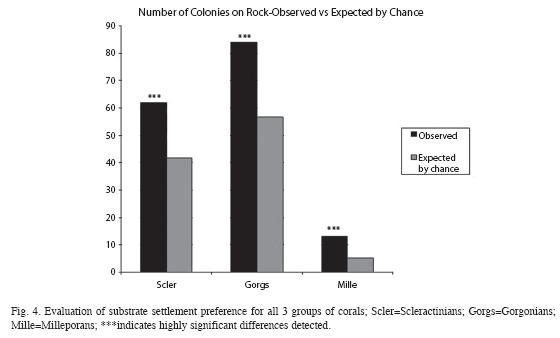
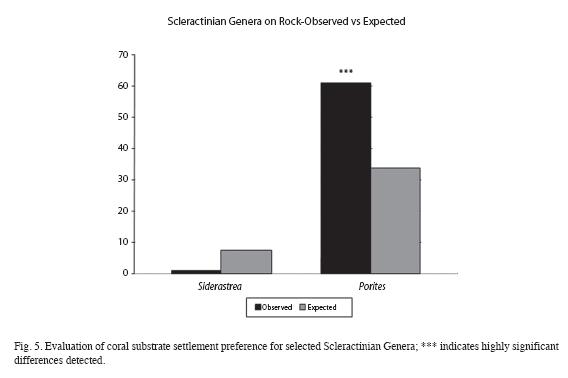
For Scleractinians, the evaluation was broken down (Fig. 5) into the Genera that comprised the majority of colonies, those being Siderastrea and Porites. (Note: this included virtually all Scleractinians; one P. astreoides settlement wasn´t categorized, and two Diploria spp. that settled on cement. P. astreoides made up 87% of total Porites, and S. siderea made up 63% of total Siderastrea).
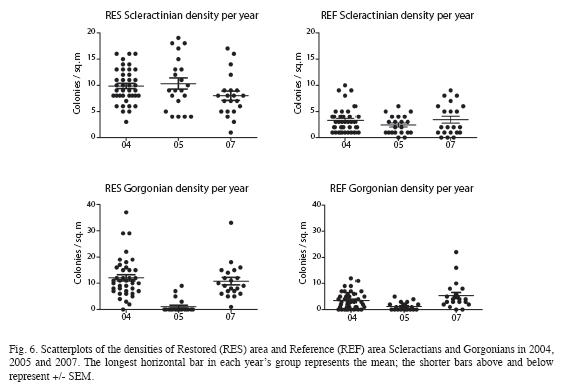
Inter-annual Comparisons: In addition to examining data for 2007, some inter-annual comparisons with previous years´ monitoring were made. In the analysis which follows, only Scleractinian and Gorgonian data will be presented (Fig. 6), as Millepora populations were too low for meaningful analysis.
For Scleractinains in the Restored area, densities were very similar for 2004 and 2005. While they appeared to slightly decrease in 2007, overall ANOVA revealed no significant differences, (p=0.1228). Likewise, in the Reference area differences were also non-significant (p=0.3226).
Discussion
The development of coral communities is well-known to be a long-term process, so NOAA cannot make definite conclusions about the success of the Maitland restoration at this stage. Additionally, coral recruitment in the Florida Keys is well-known to be comparatively low (Shearer and Coffroth 2006). Taking these locally limited circumstances into account, in general the results of this monitoring program are comparable with the results reported by other researchers (Moulding 2005).
Further, the density and composition of coral recruits at this stage provides a good indication that the stability offered by the restoration is providing suitable substrate. However, the scope of the monitoring program is somewhat limited. It tracks only certain aspects of coral restoration, namely: density, biodiversity, settlement preference, and size distribution. The effort is circumscribed by the amount of damages collected from the party responsible for the grounding. Given this inherent limitation, it was thought more important to focus on coral recovery than overall community structure (Miller et al. 2009).
As the above density data reveals, either very or highly significant differences were shown for Gorgonian and Scleractinian densities respectively (Fig. 1). The much greater densities in the Restored area give rise to some interesting questions. Are the restoration units really such great recruitment attractors, or do the densities only appear high in relation to mature, stable areas? Or, has young-adult colony mortality not yet had a chance to play a role as a structuring factor? (Recall that the monitoring protocol does not track actual recruitment and mortality, only resultant overall density. As noted, the monitoring permitted by available recovered funds does not permit tracking individual colonies, and the discrimination of population structure that might result.).
Regarding biodiversity, Tables 1, 2 and Fig. 2 reveal only slight differences by 2007 and the two areas may be fairly said to have converged by this time, at least as regards common biodiversity indicators. As the mentioned tables and charts show, large broadcasting corals (Siderastrea siderea) were disproportionately present on the restoration site.
Just why this should be so remains somewhat of a mystery; as mentioned, the issue of generally low recruitment for these corals in the Florida Keys is well known. Why then does it appear much higher in the Restored area? Certainly, one should be conservative when it comes to making predictions based upon demographics of these corals, based upon the relatively low numbers involved. However, if any reader has any thoughts or insights regarding this issue, contact with the author would be appreciated.
Thus, with the exception of the very smallest class (≤20mm), where the Restored area still had a 3.5-fold advantage (20.2 vs 5.7%), the distribution in the two zones almost mirrored each other. Besides the smallest class, the only notable exception was in the very largest (≥120mm) classes where - unsurprisingly - the Reference area retained a large advantage. Future monitoring could reveal whether the two areas eventually totally converge in this regard.
The inter-annual comparisons among the Scleractinian and Gorgonian densities (Fig. 6) were revealing. For the Scleractinians, very little difference in the Restored and Reference areas across years was apparent. There was some slight (though non-significant) decrease in density in 2007 as compared to either of two previous years. The real story to be told is that, considering the respective zones across years, the Restored area maintained approximately a three-fold advantage in population over the Reference area. Does this provide any inference regarding the most oft-cited coral health metric, percent cover? It must be remembered that the individual colonies (at least of P. astreoides, as well as certain other species1) were growing during this period (see above discussion re size-class distribution). While percent cover was not evaluated, it is likely that it would have increased.
The Gorgonians proved equally interesting. From 2004 to 2005 they suffered a precipitous decrease. As a first suspicion, one might implicate disease brought about by Aspergillosis sydowii as the cause of the rapid Gorgonian decline. Aspergillosis is a fungal pathogen responsible for considerable Gorgonian decline in the Florida Keys in the last two decades. However, at least in this case, physical ablation appears a more probable cause. The passage of the three violent hurricanes in the near vicinity between the 2004 and 2005 monitoring events has already been noted.
Further support for the proposition of a physically destructive causative factor is provided by the fact that colony densities experienced complete recovery to 2004 levels by the time of the 2007 monitoring event. Perhaps most noteworthy is that both before and after the population crash, the Restored area Gorgonians evidenced a 2.5 to 3-fold population advantage over the Reference area, proving similar to the Scleractinians is this regard.
Lastly, among the most interesting findings, the data provide some evidence that brooder-spawning Porites larvae exhibit an active settlement preference for the limestone rock substrate. This is as opposed to the cement matrix in which the rock is embedded; see Figs. 4 and 5, and the accompanying text. This finding is generally consistent with the work of Miller and Barimo (2001) regarding early settlement preference results (obtained 3 years post-settlement) conducted at this site.
Nonetheless, it should be prominently noted that nothing can be said regarding the mechanism of the preference. Are the larvae drawn to the rock because of chemotaxis due to something in the limestone, or simply because it presents more rugosity in its surface area, providing more cryptic settlement opportunities (Babcock & Mundy 1996)? (Elevation may also play a role, as the rocks project surface area above the surrounding flat, horizontal concrete.) Or, are they "put off" by chemicals leaching out of the cement, differences in pH, etc.? Most intriguing is the fact that the pattern seems to be exactly reversed for the broadcaster-spawning Siderastrea. There would appear to be something about cement that is attractive to that Genus.
Certainly it is well known that restoration substrate differences can result in variation of density of coral recruitment (Burt et al. 2009). One possible answer in this case is may be that one surface or the other is subject to greater colonization by macroalgae and/or cyanobacteria. If so, perhaps settlement by one Genus is more inhibited by the presence of such organisms (Kuffner et al 2006). Even if coral settlement takes place, it is conceivable that the mortality rate of relatively new settlers is differentially affected by the inhibiting taxa (Vermeji et al. 2009). Again, if any reader has any thoughts along these lines, contact and communication would be appreciated. This phenomenon is suggestive of future research possibilities.
Acknowledgments
Appreciation is expressed to Eileen Alicea and Gonzalo Cid, who provided the translation of the abstract. The Damage Assessment and Restoration Program of the Florida Keys National Marine Sanctuary performed the fieldwork providing the basis for the article.
Referencias
Bodge, K. 1996. Engineering Summary Report: Structural Restoration of the M/V Alec Owen Maitland and M/V Elpis Vessel Grounding Sites: Florida Keys National Marine Sanctuary. Report prepared for: Office of Ocean Resources Conservation and Assessment, Silver Spring, Maryland, USA. [ Links ]
Burt, J., A. Bartholomew, A. Bauman, A. Saif & P. Sale. 2009. Coral recruitment and early benthic community development on several materials used in the construction of artificial reefs and breakwaters. J. Exp. Mar. Biol. Ecol. 373: 72-78. [ Links ]
Continental Shelf Associates, Inc. (CSA). 1993. Coral Reef Structural Restoration Planning Field Survey Report for the M/V Alec Owen Maitland and M/V Elpis Grounding Sites in the Florida Keys National Marine Sanctuary. Report prepared for: National Oceanic and Atmospheric Administration. Silver Spring, Maryland, USA. [ Links ]
Edmunds, P. & R. Elahi. 2007. The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecol. Monographs 77: 3-18. [ Links ]
Gittings, S. 1991. Mitigtion and Recovery Enhancement at the Grounding site of the M/V Alec Owen Maitland, Key Largo National Marine Sanctuary. Report prepared for: Office of Ocean Resources Conservation and Assessment. Silver Spring, Maryland, USA. [ Links ]
GraphPad Software, Inc. 2003, 2007. InStat Biostatistics, version 3.0; Prism 5 for Windows, version 5.0. La Jolla, California, USA. [ Links ]
Kuffner I., L. Walters, M. Becerro, V. Paul & K. Beach. 2006. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 323: 107-117. [ Links ]
Miller, M. & J. Barimo. 2001. Assessment of juvenile coral populations at two reef restoration sites in the Florida Keys National Marine Sanctuary: Indicators of success? Bull. Mar. Sci. 69: 395-405. [ Links ]
Miller, M., A. Valdivia, K. Kramer, B. Mason, D. Williams & L. Johnston. 2009. Alternate benthic assemblages on reef restoration structures and cascading effects on coral settlement. Mar. Ecol. Prog. Ser. 387: 147-56. [ Links ]
Moulding, A. 2005. Coral recruitment patterns in the Florida Keys. Rev. Biol. Trop. 53: 75-82. [ Links ]
National Oceanic and Atmospheric Administration (NOAA), undated. Coral Reef Restoration Key Facts-M/V Alec Owen Maitland Grounding. Prepared by (then) Sanctuaries and Reserve Division (now Office of National Marine Sanctuaries). Silver Spring, Maryland, USA. [ Links ]
National Oceanic and Atmospheric Administration (NOAA), undated. Reef Restoration Action Project. Prepared by (then) Sanctuaries and Reserve Division (now Office of National Marine Sanctuaries). Silver Spring, Maryland, USA. [ Links ]
Shearer, T. & M.A. Coffroth. 2006. Genetic identification of Caribbean scleractinian coral recruits at the Flower Garden Banks and the Florida Keys. Mar. Ecol. Prog. Ser. 306: 133-142. [ Links ]
Vermeij, M., J. Smith, C. Smith, R. Thurber & S. Sandin. 2009. Survival and settlement success of coral planulae: independent and synergistic effects of macroalgae and microbes. Oceologia 159: 325-336. [ Links ]
Internet Reference
http://apps.isiknowledge.com/full_record.do?product=WOS&search_mode=GeneralSearch&qid=1&SID=1B@on9ie8LN4KgPMpM8&page=1&doc=15 - address000263424800008-4
1. An interesting, though far from statistically meaningful footnote: To evaluate the growth of a coral with a different life history strategy than P.astreoides, size-class frequency distributions were constructed for Siderastrea siderea, a frameworkbuilding, broadcasting coral. S. siderea also proved to be the second most abundant coral during each of the three monitoring years, and as a proportion of total Scleractinians, was consistent at about 9.5% of restoration area colonies. Absolute numbers of colonies were so low as to make a frequency distribution less than robust, and it is for this reason though the size classes utilized for S. siderea were smaller (10, 20, 30, 40, & 50mm). The smallest two size classes started out as 62% in 2004, but by 2007 had dropped to 37%, with the larger classes gaining reciprocally. Since the density of the species was almost exactly the same across years (at approx. 0.9col./m2) the colonies´growth represents an absolute increase in their percentage benthic cover.
Correspondencia: Joe Schittone. NOAA 1305 E. West Hwy, SSMC4, NMS/6, 11th Floor, Silver Spring, Maryland. USA 20910;joe.schittone@noaa.gov
Received 30-VII-2009. Corrected 05-II-2010. Accepted 12-VII-2010.

