










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.58 n.2 San José Jun. 2010
Dinámica de las propiedades del suelo en bosques de Rhizophora mangle L. (Rhizophoraceae) en Isla de Margarita, Venezuela
Luz Esther Sánchez-Arias1, Jorge Paolini2 & Jon Paul Rodríguez2
1. Centro de Estudios Botánicos y Agroforestales, Instituto Venezolano de Investigaciones Científicas Regional Zulia, Maracaibo, Venezuela; lsanchez@ivic.gob.ve
2. Centro de Ecología, Instituto Venezolano de Investigaciones Científicas, Apdo. 20632, Caracas 1020-A, Venezuela; jpaolini@ivic.gob.ve; jonpaul@ivic.gob.ve
Abstract
Dynamics of soil properties in forests of Rhizophora mangle L. (Rhizophoraceae) in Margarita Island, Venezuela. Biochemical and microbiological properties of soils can provide information related to ecosystems environmental status. With the aim to determine the response of microbial biomass, and enzymatic and microbial activity in hypersaline (IS≥55ups) and saline (IS<55ups) mangrove soils exposed to interstitial salinities (IS) greater than 36ups, these properties were measured in mono-specific forests of Rhizophora mangle at Laguna de la Restinga National Park (Margarita Island, Venezuela). During three seasons (dry, wet and transition), a total of 120 soil samples were collected from ten (5 hypersaline and 5 saline) randomly-selected sites of 1000m2 each. Four soil samples (400-500g) per plot were randomly collected with a corer at a depth of 10cm using a 1m2 quadrat; each sample consisted in the combination of 8 sub-samples (50-60g ea.). Physical, chemical, enzymatic, biochemical and microbiological properties of soil samples were determined using standard laboratory protocols. The response of microbial biomass and microbial and enzymatic activity was analyzed taking into account spatial and climatic factors and interstitial salinity. Microbial biomass was linked to each locality conditions, and was not sensitive to seasonal or salinity differences. Microbial activity remained functionally active during the study period and presented variable responses. Dehydrogenase activity proved to be a good indicator for flooded and anoxic environments, and arginine ammonification resulted to be the more sensitive microbial activity to changes in salinity. Regarding enzyme activities, spatial variability was the most widespread response. We did not find a unique general pattern between enzymatic activities and spatio-temporal variation; and only the enzyme phosphatase was negatively affected by salinity. We conclude that microbial populations of mangrove soils and their activities have functional adaptations to flooded and highly-saline environments typical of a negative estuary, subjected to drastic changes due to weather and water dynamics. Future studies are needed to determine the relation between the "health" of mangrove forest and microbial populations, and their activities in mangroves soils. Rev. Biol. Trop. 58 (2): 547-564. Epub 2010 June 02.
Key words: enzymes, Laguna de La Restinga, mangrove forest, microbial activity, Rhizophora mangle, salinity, soils.
Resumen
Propiedades bioquímicas y microbiológicas de los suelos pueden proporcionar información relativa al estado de los ecosistemas. Se determinaron las propiedades microbianas (biomasa y actividad) y bioquímicas (actividad enzimática) en suelos de bosques mono-específicos de Rhizophora mangle en Laguna de la Restinga (Isla Margarita, Venezuela), con el objeto de cuantificar cambios en áreas expuestas a salinidades intersticiales (SI) superiores a 36.0ups, clasificadas como salinas (IS<55.0 ups) e hipersalinas (SI≥55.0 ups). Se ubicaron al azar 5 localidades salinas y 5 hipersalinas, y durante tres épocas climáticas se tomaron 4 muestras de suelo por localidad (120 muestras en total). Se utilizó análisis de varianza por permutaciones multivariadas (PERMANOVA) para el análisis de los resultados. La biomasa microbiana se vio afectada por las condiciones particulares de cada localidad, y no fue sensible a los cambios de época climática o salinidad. La actividad microbiana se mantuvo funcionalmente activa. Se destacó el diacetato de fluoresceina (DAF) y la enzima (ß-glucosidasa), como probables indicadores tempranos de degradación. La deshidrogenasa resultó ser un buen indicador de ambientes inundados y anóxicos. Se concluye que las poblaciones microbianas muestran actividades que sugieren su adaptación funcional a ambientes altamente salinos e inundables, sujetos a cambios drásticos debido a las dinámicas climáticas e hídricas propias de un estuario negativo.
Palabras clave: actividad microbiana, bosque de mangle, enzimas, Laguna de La Restinga, Rhizophora mangle, salinidad, suelos.Los manglares conforman áreas extensas de bosques en las costas de zonas tropicales y subtropicales del planeta, adaptados a las condiciones de salinidad e inundación propias de un humedal costero (Spalding et al. 1997, Yap 2000, FAO 2007). El papel de la actividad microbiana en los ecosistemas de manglar ha sido descrito por varios autores. Sherman et al. (1998) indican que la relación planta/suelo/ microorganismos contribuye a los patrones de distribución de las especies de manglar a lo largo de la zona intermareal debido a la competencia intra-específica favorecida por comunidades microbianas. Vásquez et al. (2000) reportan la existencia de comunidades de microorganismos solubilizadores de fósforo en la rizósfera de Avicennia germinans y Laguncularia racemosa en una laguna costera semiárida y sugieren que estos organismos pueden servir como un mecanismo para solubilizar el fosfato de calcio. Holguín et al. (2001) indican que la dinámica del hierro, fósforo y azufre está íntimamente ligada a las bacterias sulfatoreductoras (los primeros descomponedores en los suelos anóxicos de manglar). Paolini & Sánchez (2008), muestran una alta actividad microbiana y enzimática en suelos rizófericos de Rhizophora mangle, que es superior a la de bosques tropicales continentales.
De acuerdo con Ashton et al. (1999), el ecosistema de manglar sustenta una amplia y variada diversidad microbiana. Esto se debe posiblemente a la rápida descomposición de la hojarasca y a la importancia del detrito como base de la cadena alimenticia en los ambientes marino costeros del manglar (Lacerda et al. 2002). La calidad del suelo de los bosques depende de numerosas variables físicas, químicas, biológicas y bioquímicas. Dentro de estas variables, las microbiológicas (biomasa y actividad) y las bioquímicas (actividad enzimática) son las más sensibles y por lo tanto pueden proveer información de los cambios ocasionados por el estrés ambiental (Vanhala & Ahtiainen 1994). Si se tiene en cuenta que la actividad microbiológica es esencial para mantener la calidad de los suelos (Pascual et al. 2000), la biomasa microbiana y su actividad son aspectos que deben ser considerados para determinar el estado de un sistema, o para entender cómo la actividad humana altera los ciclos biogeoquímicos al comparar zonas naturales y degradadas (Bastida et al. 2006). Las enzimas del suelo, por su parte, actúan sobre sustratos orgánicos haciendo disponible los nutrientes para las plantas, relacionándose así con los ciclos de nitrógeno (ureasa-proteasa), fósforo (fosfatasa) y carbono (β-glucosidasa) (Dinesh et al. 1998, Dinesh et al. 2004), entre otros.
Si bien existen investigaciones donde se identifican las diferentes comunidades microbianas en suelos de manglar (Sherman et al.1998, Holguín et al. 2001), llama poderosamente la atención la ausencia de trabajos relacionados con la actividad microbiana y enzimática de los suelos, donde sólo se registran dos trabajos sobre el tema en manglares de la India (Dinesh et al. 1998, 2004). Este trabajo estudia la actividad microbiana y enzimática de suelos de manglar y la respuesta de la biomasa microbiana a los cambios espacio-temporales de salinidad en un estuario negativo, donde la evaporación es superior al aporte de agua dulce. Además ofrece un marco de referencia para posteriores estudios relacionados con procesos de degradación de bosques de manglar debidos a procesos de salinización.
Materiales y métodos
Descripción del sitio: Este estudio fue llevado a cabo en bosques de manglar asociados a un estuario negativo durante el período comprendido entre agosto de 2005 y mayo de 2007. Se seleccionaron zonas de manglar ubicadas en el sistema lagunar costero del parque nacional Laguna de la Restinga, en la Isla de Margarita, Venezuela (10˚58’15"-11˚05’22" N y 64˚01’32"-64˚17’09" W). Catalogado por su importancia ecológica como un sitio Ramsar, el parque nacional se localiza en un ambiente árido, con una precipitación anual comprendida entre 200-600mm (Lentino & Bruni 1994). Los meses más secos corresponden a marzo-mayo y los más lluviosos a julio-septiembre; el período de transición lluvia-sequía va de diciembre a febrero, donde las lluvias disminuyen gradualmente en intensidad. La Laguna de la Restinga comprende un área 25km2 y carece de fuentes permanentes de agua dulce (Lentino & Bruni 1994). Presenta una salinidad promedio de 40±2ups y una distribución espacial de la vegetación de manglar muy definida (Fig. 1). En la zona sur y central, donde el agua fluye con regularidad, se encuentra Rhizophora mangle representado por árboles de gran porte. En las áreas internas, predomina Rhizophora mangle, pero de menor porte, y en la zona de canales se observan bosques uniespecíficos de Avicennia germinans. La tercera zona corresponde a los extremos (oriental y occidental), donde no hay mangle (Monente 1978).

Caracterización de la vegetación: A través de un sistema de información geográfica (SIG) entre agosto de 2005 y enero de 2006, se identificaron al azar 10 localidades de estudio, conformadas por bosques de Rhizophora mangle: 5 salinas y 5 hipersalinas. En cada localidad, se delimitaron parcelas de 1000m2 y se contó el número total de individuos (Cintrón & Novelli 1984). La altura de los árboles fue estimada usando una base de referencia (altura de una persona: 1.80m) y el diámetro a la altura del pecho (DAP) se midió a 1.30m del suelo. La abundancia (%) de árboles de diferentes tallas fue clasificada según las categorías diamétricas forestales: brinzal (DAP<5cm); latizo (DAP 5-15cm) y fustes (DAP>15cm) (Sánchez-Páez et al. 2000).
Muestreo: La toma de muestras de suelo y aguas intersticiales se realizó en bajamar. Las mediciones del agua intersticial se hicieron in situ. En cada localidad, se midió la temperatura, oxígeno disuelto y pH del agua intersticial introduciendo un tubo de PVC en el suelo y luego una sonda multiparamétrica (Sanchez- Arias, 2008). Se tomó una muestra del agua intersticial mediante el uso de una inyectadora de 10ml y una manguera de plástico que se introdujo en el tubo, con la muestra obtenida se midió in situ la salinidad utilizando refractómetro de campo. Adicionalmente, el nivel freático se midió con una reglilla, teniendo como punto de referencia la superficie del suelo. Para cada parcela de 1000m2 se tomaron cuatro muestras de suelo al azar utilizando una cuadrícula de 1m2. Cada muestra de suelo consistió en la combinación de 8 sub-muestras/ cuadrícula (50-60g. cada una) tomadas con un núcleo de 1 pulgada de ancho a una profundidad de 0-10cm (total 400-500g de suelo). Las muestras de suelo se colocaron en bolsas plásticas, se trasportaron al laboratorio y se mantuvieron almacenadas a 4oC. Se realizaron muestreos en tres temporadas: seca (abril-mayo 2006), lluviosa (septiembre- octubre 2006) y de transición a seca (enero-febrero 2007). En total, se tomaron 120 muestras de suelo (10 localidades, 4 muestras por parcela y 3 temporadas de muestreo).
Análisis de laboratorio: Se realizaron en las muestras de suelo y se determinaron las variables fisicoquímicas relacionadas con los microorganismos del suelo: salinidad (pasta saturada, US Department of Agriculture 1950), pH (FONAIAP 1990), carbono orgánico total (Anderson & Ingram 1993) y fósforo total se realizó por digestión húmeda por el método del peróxido y determinación por el método colorimétrico azul de molibdeno (Murphy & Riley 1962) de acuerdo a protocolos estándar. Igualmente, se realizaron análisis bioquímicos y microbiológicos. Se determinaron las propiedades bioquímicas generales: la respiración basal del suelo (Alef & Nanipieri 1995), la biomasa microbiana por respiración inducida por sustrato (Anderson & Domsch 1978). También se determinaron las propiedades bioquímicas específicas: fosfatasa ácida relacionada con el ciclo de P (Tabatabai & Bremner 1969), ureasa (Kandeler & Gerber 1988) y caseinasa (Ladd & Butler 1972), relacionadas con el ciclo N, y β-glucosidasa, relacionada con el ciclo C (Tabatabai 1994). Tambiιn se determinaron enzimas relacionadas con la actividad microbiológica: deshidrogenasa (Casida et al. 1964), hidrólisis del diacetato de fluoresceína (Schnürer y Roswall 1982) y amonificación de arginina (Alef & Kleiner 1986). Cada análisis se realizó por quintuplicado (3 muestras y dos blancos).Todos los resultados se expresan como suelo seco.
Para establecer las relaciones entre clima, variación espacial y salinidad, y las variables de suelo, se utilizó un análisis de varianza multifactorial con interacción basada en permutaciones multivariadas (PERMANOVA) adecuado para sistemas abiertos y dinámicos (Anderson 2005). El PERMANOVA permite evaluar hipótesis en sistemas ecológicos complejos con variaciones naturales temporales y espaciales; tiene su origen en la necesidad de los ecólogos de un método robusto, interpretable con referencia experimental, multivariado y no paramétrico, que distribuya la variación en cualquier medida de distancia para cualquier diseño multifactorial (Anderson 2005). Para asegurar que las observaciones son intercambiables, las muestras fueron escogidas al azar y en igual número; se determinaron las medias, desviación estándar y coeficientes de variación empleando MS Excel, exportándose los datos posteriormente al software de PERMANOVA. Los factores fijos del PERMANOVA fueron la época climática (seca, lluvia, transición) y el tipo de salinidad (salina: S.I.<55ups, hipersalina: S.I.≥55ups), mientras que el factor al azar, anidado al factor salinidad, fue la variación espacial dada por localidad. El diseño contempló las 4 réplicas para cada una de las diez localidades (5 salinas y 5 hipersalinas), tomadas durante 3 épocas climáticas (n=120). Con el fin de determinar la estructuración del conjunto de datos multivariados, se utilizó el programa PRIMER a partir de varianzas permutadas y se aplicó la prueba de similitud SIMPER (Clarke & Warwick 2001) para determinar que parámetros eran comunes entre los diferentes bosques.
Resultados
Características generales de las localidades: Los bosques de Rhizophora mangle en el área de estudio presentaron fustes de diámetro promedio 22.3±7.2cm, latizos con un diámetro promedio de 9.2±3.6cm y brinzales con valores promedio de 2.0±1.3cm. En todas las localidades, la categoría diamétrica predominante fue la de latizos y las alturas del dosel promedio variaron entre 3.2 y 10.0m (Cuadro 1). El oxígeno disuelto en el agua intersticial no superó el valor de 0.7±0.02mg/l y el pH de las aguas intersticiales fue 6.5±0.1. La temperatura del agua intersticial presentó la mayor variación (26.5-27.0ºC) entre las diferencias épocas, pero sus diferencias no fueron significativas (p>0.05). Durante el periodo de muestreo, las localidades de estudio presentaron variación en los niveles de altura de sus aguas intersticiales (nivel freático). Es así como en la época seca el nivel medio de las aguas intersticiales estuvo a -15±2cm de la superficie mientras que en época de lluvia fue -5±0.5cm y en época de transición -25±3cm. Tanto para las mareas como para el nivel freático, los mayores descensos de agua se dieron en la época de transición (20-30cm) (Fig. 2).

Parámetros fisicoquímicos
Salinidad: En general, la salinidad intersticial y la del suelo disminuyeron en la época de lluvia (Cuadro 2), pero al realizar el análisis por localidad no se observa una tendencia uniforme (Fig. 3). La salinidad intersticial varió significativamente por efecto de la interrelación entre las épocas climáticas y las características de cada localidad. (p=0.002). En cuanto a la salinidad del suelo entre localidades, se observa que la interacción del efecto de las épocas climáticas (EP), el tipo de salinidad (TSI) y las localidades (LO), no explica de forma significativa la variación presentada en la salinidad del suelo (p=0.105).
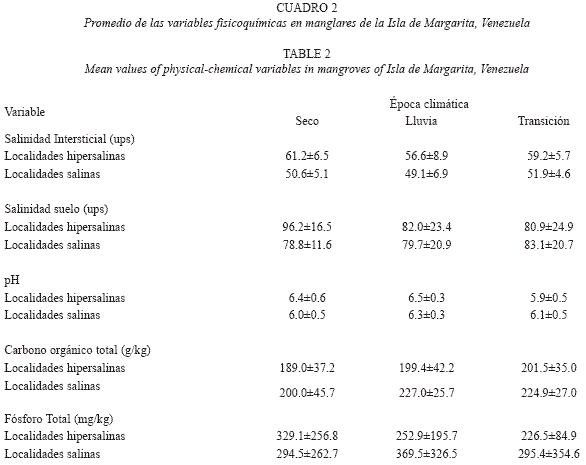
pH: El pH en los suelos con vegetación de Rhizophora mangle fue ligeramente ácido, con un valor promedio de 6.25±0.55, siendo ligeramente menos ácido en las parcelas hipersalinas (Cuadro 2). No existen diferencias significativas debidas a la interacción entre épocas climáticas, salinidad y localidades (p=0.37).
Carbono orgánico total (COT): El promedio del contenido de carbono orgánico total estuvo entre 200.0±45 y 227.0±25.7g/kg para las localidades salinas y entre 189.0± 7.2 y 201.5±35.0g/kg para las localidades hipersalinas (Cuadro 2). No hubo diferencias significativas debidas a la variación temporal, ni a la salinidad (p= 0.32), pero si se detectaron diferencias debidas a la variación espacial ligadas a cada localidad (p=0.002) (Cuadro 2).
Fósforo total (FT): Las localidades salinas presentaron mayores contenidos de fósforo total que las hipersalinas, 398.0±312.0mg/kg y 325.6±197.0mg/kg, respectivamente (Cuadro 2). Sin embargo, la salinidad, épocas climáticas y localidades no tuvieron efecto significativo sobre la variación del fósforo total contenido en el suelo (p=0.84).
Parámetros microbiológicos y bioquímicos generales: Los valores encontrados muestran diferentes respuestas de acuerdo al tipo actividad microbiana, localidades, tipo de salinidad en las localidades (salinas/hipersalinas) y efecto sinérgico debidos a las época climática (Cuadro 3).

Carbono microbiano: Los mayores valores se registran en las localidades hipersalinas (287.0±174.5 a 333.5±308.3mg Cmic/kg suelo) (Cuadro 3). En la poca de transición, se presenta un incremento respecto a las otras épocas, tanto para las parcelas salinas (264.4±124.6mg Cmic/kg suelo) como para las hipersalinas (333.5±308.3mg Cmic/kg suelo). Sin embargo, es notoria la variabilidad registrada en todas las localidades, presentándose coeficientes de variación superiores al 50%. Al realizar el análisis se indica que los cambios registrados en el carbono microbiano son afectados por las características de cada localidad ligada a la variación espacial y a la salinidad (p=0.0002).
Respiración basal: En general, los mayores valores se registraron en las localidades hipersalinas, acentuándose la diferencia durante la época de transición (Cuadro 3). Se observa un descenso durante la época seca en todas las localidades, siendo las respuestas más marcadas en las localidades hipersalinas (Cuadro 3). El análisis PERMANOVA indica que la respuesta de la respiración basal está afectada significativamente por la interrelación entre los factores considerados en este estudio y no por su efecto individual (p=0.0268).
Amonificación de la arginina: Esta actividad presentó una alta variabilidad tanto en las localidades salinas como hipersalinas, especialmente durante la época de lluvia, El máximo valor se observó en la época de transición para las parcelas salinas (Cuadro 3). Independiente de la localidad, el efecto conjunto entre la salinidad y la época climática determinan la respuesta de esta actividad microbiana en el suelo de manglar (p=0.008).
Hidrólisis del DAF: Los valores estuvieron entre 216.7±97.6 y 439.8±188.2μg fluoresceνna/g suelo·h para las localidades hipersalinas y entre 229.2±92.0 y 516.0±181.6μg fluoresceνna/g suelo·h para las localidades salinas. Se registraron diferencias entre las localidades salinas e hipersalinas, las cuales se mantuvieron durante todo el período de estudio. La época de lluvia presentó los mayores valores para todas las localidades (Cuadro 3). Independiente de la localidad, la interacción de las épocas climáticas sobre la salinidad determina la respuesta de esta actividad. (p=0.0002).
Actividad deshidrogenásica: Presentó los mayores valores durante la época de lluvia tanto para las localidades salinas (1 261.5±1 037.0μg TFF/g suelo·24h) como para las hipersalinas (1 255.8±1 196.5μg TFF/g suelo·24 h). En general, se puede decir que la ιpoca seca presenta la menor variabilidad si se compara con las otras temporadas (Cuadro 3). Independiente de la localidad, la acción conjunta de la salinidad y las diferentes épocas climáticas ejerce un efecto significativo sobre la actividad de la deshidrogenasa (p=0.0008).
Parámetros bioquímicos específicos: Los parámetros bioquímicos específicos del suelo en las zonas con bosques de Rhizophora mangle presentaron una alta variabilidad. En general se observó un descenso en la respuesta de los mismos durante la época seca, mientras que la respuesta ante la lluvia y el período de transición dependió de cada tipo de enzima (Cuadro 4). La ureasa, la fosfatasa y la caseinasa presentaron sus mayores valores en la época de lluvia, mientras que la glucosidasa presentó una mayor actividad en la época de transición (Cuadro 4). A pesar de estas variaciones, al aplicar el análisis multifactorial para la ureasa (p=0.0002), fosfatasa (p=0.0002), ß-glucosidasa (p=0.001) y caseinasa (p=0.0002), se observó que independiente de la salinidad y del clima, son las características específicas de las localidades las que determinan las diferencias encontradas.

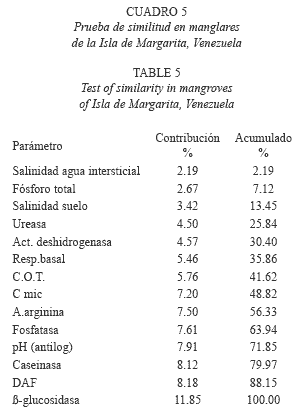
Variables más comunes entre los bosques Rhizophora mangle: Al analizar la similaridad de las variables fisicoquímicas y bioquímicas estudiadas bajo la prueba SIMPER de similaridad (Cuadro 5), las variables más comunes entre los bosques y que representan el 69.60% de la información están relacionadas de forma directa con la biogeoquímica del manglar fueron: actividad enzimática de ß-glucosidasa (11.85%), actividad de DAF (8.18%), actividad de caseinasa (8.12%), pH (7.91%), fosfatasa (7.61%), amonificación de la arginina (7.50%) y carbono microbiano (C mic) (7.20%), estas tres últimas con contribuciones muy similares.
Discusión
El período de muestreo (abril 2006 a febrero 2007) se caracterizó por presentar menores valores de precipitación que el registro histórico (1972-1996), especialmente para noviembre de 2006 (10.0mm o 1/3 del histórico) (Sánchez- Arias 2008).
La Laguna de la Restinga presenta bosques de bajo porte que corresponden a ambientes áridos e hipersalinos, sin embargo llama la atención que se presenten promedios mayores de altura en algunas localidades hipersalinas al comparar con las localidades salinas (Cuadro 2), lo que puede indicar que la salinidad no ha sido el único factor limitante en el desarrollo sino que pueden existir relaciones a nivel de las características de cada sitio que marquen las diferencias. Por otro lado, al analizar las salinidades del agua intersticial y su relación con el estado del bosque de Rhizophora mangle en las localidades hipersalinas, se encontró que aunque la salinidad del agua intersticial era superior a los 55ups, no se evidenciaban signos de estrés como defoliación, clorosis masiva, o árboles muertos que pusieran en evidencia un riesgo en la persistencia del arbolado, a pesar de ser considera esta especie como una de las más suceptibles a la hipersalinización (Lüttge et al. 2002). Por lo tanto, según los valores de salinidad es posible inferir que un proceso de salinización ya se haya iniciado en las zonas con vegetación de la Laguna de La Restinga, pero que el equilibrio aún se mantenga o que la salinidad del agua intersticial no sea el único factor limitante en la permanencia del bosque. A este respecto (Marchand et al. (2004) indican que Rhizophora mangle requiere de pulsos ocasionales de agua dulce para mantenerse en ambientes hipersalinos. Durante el período de estudio se registraron pulsos de agua menos salina, incluso la salinidad del agua intersticial disminuyó en un 10% (Cuadro 2) y en algunos casos se evidenció una menor salinidad de los suelos en las épocas de lluvia, que podrían ser suficientes para mantener la vegetación. Dado que la variación del nivel freático se relacionó con el ritmo mareal, podrían estar ocurriendo procesos de lavado de sales, así como transporte de materia orgánica y nutrientes. Lo anterior, coincide con el incremento de la salinidad en el agua intersticial en otras épocas del año. Además, si se examinan los contenidos de sales de las localidades de forma individual, se observa variabilidad (Fig. 3). La explicación puede estar vinculada con la hidrodinámica, tratada específicamente como la forma en que afectan las mareas a los diferentes suelos de las localidades. Otro factor que puede afectar el contenido de sales en el suelo es la microtopografía de cada localidad, ocasionando que durante el período seco y de transición (cuando las bajamares son mayores) la precipitación de las sales varíe espacialmente (Ridd & Stieglitz 2002). Esta variación espacial debida a la microtopografía fue observada en diferentes parámetros del suelo de manglar por Rivera- Monroy et al. (2004).
Ya que los bosques de Rhizophora mangle de las localidades con vegetación presentan una densidad de fustes y latizos vivos superiores a 60% (Cuadro 1), se podría sugerir que bajo las condiciones dadas en el área de estudio, no ha habido limitaciones de los elementos esenciales que afecten la sobrevivencia del bosque. Se puede sugerir que esto puede ser debido a que la mineralización de la materia orgánica está determinada por una comunidad de microorganismos del suelo (como los solubilizadores de fosfato), adaptados a la hidrodinámica y a los valores de salinidad registrados, los cuales también dependen de un pH óptimo, así como del aporte dado por la acumulación y descomposición de la hojarasca (Sánchez-Páez et al. 2001). Sin embargo, sólo cuando se profundice el conocimiento sobre las características bioquímicas y biológicas del suelo de manglar se podrá afirmar lo anterior.
Los valores de pH encontrados en los suelos de manglar de la Laguna de La Restinga (Cuadro 2), están dentro del intervalo reportado por otros autores en Venezuela (Pannier & Pannier 1985), Brasil (Ferreira et al. 2007) y Australia (Boto & Wellington 1984). Los valores ligeramente ácidos de pH registrados en el presente estudio pueden ser debidos a las condiciones anóxicas que promocionan procesos de reducción (Breemen & Buurman 1998), a la actividad microbiana que influye en el descenso del pH o por efecto mismo de la planta a través de exudados o producción de CO2 (Hall 2002). También se ha determinado que el pH del suelo depende en gran medida de las características específicas de cada suelo y de su hidrodinámica. Por ejemplo, cuando la marea lava un suelo salino puede eliminar iones Cl-o introducir iones CO3-2 y HCO3-, los cuales tenderían a aumentar el pH (Chen & Twilley 1999, Muhammad et al. 2008).
La ausencia de diferencias en COT para las épocas climáticas y para la salinidad, se puede explicar por el hecho de que los suelos fueron orgánicos de tipo fibrist, y porque el COT es un parámetro que responde de manera lenta a los cambios, y por lo tanto, se requiere de mediciones a lo largo de períodos prolongados para detectar su variación. Igualmente, el COT incluye compuestos, sin distinguir si son lábiles o recalcitrantes. Gran parte de la materia orgánica del suelo fibrist de los bosques de manglar es derivada de las raíces las cuales pueden permanecer sin descomponerse por largo tiempo (Marchand et al. 2003). La variación espacial encontrada se puede deber a las características propias de cada localidad (microtopografía), al aporte de material orgánico autóctono o alóctono (Ellison, 1998, Marchand et al. 2004), a la productividad del bosque (Lacerda et al. 1995), a condiciones de anoxia (Bellanger et al. 2004), y a la presencia de fauna o algas (Marchand et al. 2003).
Los resultados obtenidos en cuanto al FT (Cuadro 2) están dentro de los valores reportados por Chacón et al. (2008) para bosques tropicales no salinos con períodos de inundación superiores a cinco meses (318 a 373mg P/kg), pero por debajo de los reportados por Alongi et al. (1993) en bosques de manglar de Nueva Guinea (fósforo total de 780 a 830mg P/kg) y los reportados por Alongi & Carvalho (2008) en bosques mixtos de manglar ubicados en áreas áridas (503mg P/kg). Tanto en bosques inundables como en las zonas de manglar (Alongi et al 1993, Alongi & Carvalho 2008, Chacón et al. 2008), se menciona una limitación de fósforo, por lo que es de esperarse que el fósforo también sea limitante en el presente estudio. A pesar de esto, en La Restinga se registran bosques donde 60% de los árboles tienen DAP>10cm (Cuadro 4). Podría esperarse que en las localidades de estudio (salinas o hipersalinas) existan poblaciones microbianas halotolerantes, con mecanismos para responder de forma eficiente a las necesidades nutricionales del bosque de manglar y a las condiciones salinas imperantes (Zahran et al. 1992). Es posible también que el fósforo total esté relacionado no sólo con los contenidos de COT encontrados en este estudio, sino que además esté relacionado con los aportes alóctonos durante la época de lluvias. Los coeficientes de variación superiores a 50% (Cuadro 2) reflejan que dentro de los bosques de un mismo tipo existe una gran variabilidad en el contenido de nutrientes debido a la geomorfología, la amplitud mareal y la época climática (Alongi et al. 1993).
En cuanto a la biomasa microbiana, es posible que los suelos analizados en este estudio posean poblaciones halotolerantes estables que estén adaptadas a las condiciones locales y que a su vez, la vegetación brinde ambientes propicios para el crecimiento microbiano, (Paolini & Sánchez-Arias 2008). Si se observa el Cuadro 2, los mayores valores se dan en las parcelas hipersalinas, pero al mismo tiempo estas presentan la mayor variabilidad. El PERMANOVA indica que la diferencia encontrada no depende sólo del clima sino de la interrelación entre éste y las características de las localidades (variación espacial), Poret-Peterson et al. (2007) y Pezeshki (2001) han registrado cambios en la biomasa microbiana en ambientes costeros y humedales debido a la variación del ambiente.
En cuanto a la actividad microbiana, el descenso observado en la respiración basal durante la época seca constituye una respuesta típica que se registra en ecosistemas terrestres afectados por salinidad, que se puede atribuir a que gran parte de la población microbiana muere o permanece inactiva por las condiciones desfavorables debidas al estrés osmótico ocasionado por las sales y la baja disponibilidad del agua (Frankenberger & Bingham 1982, Yuan et al. 2007). Por otro lado, una mayor respiración basal puede indicar que los microorganismos están mineralizando con eficiencia la materia orgánica del suelo, es una respuesta de las poblaciones ante cambios ambientales o estrés.
Los resultados obtenidos para la amonificación de la arginina sugieren que existe una población de hongos y bacterias que utilizan este aminoácido en los diferentes bosques, y que su variabilidad se puede deber a que es un ecosistema abierto que sufre cambios producidos por las épocas climáticas y la salinidad (Frankenberger & Bingham 1982, García & Hernández 2000). En los ecosistemas terrestres, la salinidad conduce a una menor proliferación de hongos y actinomicetos relacionados de forma directa con la amonificación de la arginina, lo que podría estar ocurriendo durante la época seca en las zonas de manglar.
La hidrólisis de DAF mide la actividad heterotrófica del suelo (Kassen & Kleiner 2001), la cual resultó afectada significativamente por las diferentes épocas climáticas y la salinidad intersticial de los suelos (Cuadro 3). Esto contrasta con los estudios de Bandick & Dick (1999) y Dumontet et al. (2001), quienes afirman que DAF es relativamente estable al clima en ambientes tropicales. Se registraron diferencias significativas entre localidades salinas e hipersalinas, lo que coincide con los resultados obtenidos para amonificación de la arginina. Estos pueden estar asociados con diferentes poblaciones heterotróficas que se encuentran distribuidas en el total de las localidades pero que son susceptibles a los cambios climáticos y a la salinidad. Los valores obtenidos están muy por encima de los valores encontrados en sistemas salinos ubicados en zonas áridas (de 9.4±1.5 a 57.7±4.8μg fluoresceína/g suelo· h) (Yuan et al. 2007), lo que puede estar asociado con una mayor productividad del ecosistema de manglar.
La actividad de la deshidrogenasa refleja la totalidad de la actividad oxidativa de la biomasa microbiana y ha sido relacionada con poblaciones de microorganismos anaeróbicos. Los valores registrados en este estudio son menores a los reportados por Dinesh et al. (2004) en bosques de manglar de la India (3 115,92±745,2μg TFF/g · 24h), y estos pueden explicarse a travιs de las diferencias en salinidad de ambos estudios. Así, Dinesh et al. 2004 registran valores menores de 15ups de salinidad intersticial, mientras que este estudio registra valores que superan los 40ups. Los trabajos de Frankenberger & Bingham (1982) y Tripathi et al. (2007) apoyan esta afirmación, al señalar que dicha actividad es afectada negativamente por la salinidad. Por otra parte, los valores obtenidos en este estudio superan los valores encontrados por Tripathi et al. (2007), quienes reportan valores comprendidos entre 144 y 32μg TFF/g· suelo 24h para los suelos de αreas costeras de la India bajo cultivo de arroz. Esto puede explicarse por las condiciones particulares de este estudio (inundación, bajos contenidos de oxígeno disuelto, altos contenidos de materia orgánica) y por el trabajo de Rossel et al. (1997), que indica que esta enzima intracelular está asociada con los microorganismos proliferantes anaeróbicos y no es estabilizada por los coloides inorgánicos (arcillas) y orgánicos (sustancias húmicas) del suelo. De hecho, algunos estudios demuestran que las poblaciones de organismos anaerobios son mayores a los aerobios en suelos de manglar (El-Tarabily 2002, Holguín & Bashan 2007). La variabilidad puede estar asociada a la incidencia de las mareas y a la microtopografía de cada zona. En la época de lluvia, la deshidrogenasa se incrementa, lo que podría entenderse como una respuesta a la dilución de sales, a un incremento de materia orgánica y al aumento de los períodos de inundación.
La variabilidad registrada en las actividades enzimáticas coincide con el trabajo de Bergstrom et al. (1998), quienes concluyen que una explicación de esta variabilidad es que la actividad enzimática está ligada a la distribución de las propiedades del suelo, como humedad y contenido de carbono orgánico, y que estas propiedades no se distribuyen uniformemente. Adicionalmente, Amador & Jones (1997) indican que la variabilidad también es afectada por la distribución de la biomasa radicular. Si tenemos en cuenta lo anterior, aunado al hecho de que el humedal de manglar es afectado continuamente por la mareas, se puede afirmar que esta variabilidad es una de las características de las zonas de manglar. De todas las enzimas estudiadas, la ß-glucosidasa y la caseinasa y fueron las que presentaron las menores variaciones. La primera es una fuente importante de energía para los microorganismos, implicada en la degradación enzimática de la celulosa (Turner et al. 2002) y la segunda es una proteasa que interviene en la ruptura de los enlaces peptídicos de las proteínas y está relacionada con el ciclo del nitrógeno (Ladd & Butler 1972).
Los valores más comunes de acuerdo al análisis de similitud entre los bosques (SIMPER), correspondieron a actividades microbianas y enzimáticas, y a parámetros físicoquímicos relacionados con las mismas. La enzima más común entre los bosques fue la ß-glucosidasa, una hidrolasa de origen mayoritariamente fúngico (Eivazi & Tabatabai 1990) que está relacionada con el ciclo del carbono y se cree que es fuente de energía importante para las poblaciones microbianas (Tabatabai 1994). Esta hidrolasa, participa en la ruptura de carbohidratos presentes en el suelo o en los residuos de plantas en descomposición y juega un papel importante en la degradación de los materiales celulósicos (Hayano & Tubaki 1985, Sarathchandra & Perrot, 1984). Cundell et al. (1979) describieron la importancia de los hongos en la degradación de las hojas senescentes de Rhizophora mangle, recalcado recientemente por Holguín et al. (2001). Le sigue en importancia la actividad microbiana DAF, que mide actividad heterotrófica y ha sido correlacionada con poblaciones microbianas anaeróbicas (Kassen & Kleiner 2001), las cuales son típicas de ambientes inundables. El tercer lugar corresponde a la caseinasa, indicadora de la actividad bacteriana y de la mineralización potencial del nitrógeno (Alef & Nannipieri 1995).
De acuerdo a los parámetros comunes se puede inferir que los suelos de las zonas de manglar de la Laguna de la Restinga, presentan una diversidad de microorganismos que mantienen su funcionalidad y que están adaptados a las condiciones críticas y fluctuantes (Cuadros 2 y 3). La variabilidad temporal encontrada en la actividad microbiana puede indicar que, dependiendo de las condiciones físicas, químicas y biológicas, algunas comunidades dominen y posteriormente sean remplazadas por otras cuando cambien las condiciones, todo en un equilibrio dinámico. Esta estrategia de cambio basada en una mayor diversidad microbiana ha sido relacionada con una mayor capacidad de respuesta ante los factores de estrés (Caldwell 2005).
Las poblaciones microbianas de los bosques de Rhizophora mangle de la Laguna de la Restinga, muestran actividades que sugieren su adaptación a estos ambientes altamente salinos, inundables y con bajos contenidos de oxígeno, sujeta a cambios drásticos debido a las dinámicas climática e hídrica propias de un estuario negativo. La salinización de los suelos de manglar es uno de los principales efectos causados por la alteración de la dinámica hídrica en ambientes áridos y semiáridos. Este estudio sugiere que las variables microbianas y bioquímicas son sensibles a los cambios y por lo tanto pueden proveer información de efectos a corto plazo, lo cual puede servir como herramienta de diagnóstico en el momento de evaluar el estado de degradación de un bosque de manglar. Igualmente, se destaca la actividad microbiana, el diacetato de fluoresceina (DAF) y la enzimática (ß-glucosidasa) como probables indicadores tempranos de degradación, por ser comunes a todos los bosques estudiados, al estar involucrados en los ciclos del carbono, ser fuente de energía, representar comunidades heterotróficas y anaeróbicas y ser sensibles a la salinidad.
Igualmente, la actividad deshidrogenásica es un posible indicador de procesos de estancamiento de las aguas. Los resultados obtenidos para COT y fósforo total, permiten inferir que bajo las condiciones dadas en este estudio, dichos parámetros no muestran una alta sensibilidad para detectar cambios en el suelo por procesos de salinización. Por otro lado, la salinidad intersticial no es un buen indicador de un proceso de salinización a diferencia de la salinidad del suelo. Se requieren estudios adicionales que determinen las relaciones entre las poblaciones microbianas del suelo y su actividad, y el estado de salud de los manglares.
Agradecimientos
A Diana Benzo, Yully Murillo, Henry Ramos, Alberto Oropeza, Nelda Dezzeo, Noemí Chacón, Gisela Cuenca y Milagros Lovera por el apoyo brindado durante los análisis de laboratorio. A Augusto Ruiz, Sergio Zambrano y Rodrigo Lazo por su apoyo con el sistema de información geográfica y a Edlin Guerra por su asesoría con los programas estadísticos. Al personal de PROVITA y su equipo de biomonitores, por su constante respaldo durante los trabajos de campo - especialmente Douglas y Robert. Al personal de Inparques que labora en el Parque Nacional "Laguna de La Restinga". A PROVITA (a través del Neotropical Migratory Bird Conservation Act del U.S. Fish & Wildlife Service) y al Instituto Venezolano de Investigaciones Científicas (IVIC) por su apoyo financiero y la beca otorgada a Luz Esther Sánchez para realizar estudios de postgrado.
Referencias
Alef, K & D. Kleiner. 1986. Arginine ammonification a simple method to estimate microbial activity potential in soils. Soil. Biol. Biochem. 18: 233-235. [ Links ]
Alef, K & P. Nannipieri. 1995. Soil respiration. Methods in applied soil microbiology and biochemistry. Academic, Londres, Inglaterra. [ Links ]
Alongi, D.M & N.A Carvalho. 2008. The effect of smallscale logging on stand characteristics and soil biogeochemistry in mangrove forests of Timor Leste. Forest Ecol. Manage. 255: 1359-1356 [ Links ]
Alongi, D.M., P. Christoffersen & F. Tirendi. 1993. The influence of Foresttype on microbial-nutrient relationships in tropical mangrove sediments. J. Exp. Mar. Biol. Ecol. 171: 201-223. [ Links ]
Amador, J.A & R.D Jones. 1997. Response of carbon mineralization to combined changes in soil moisture and carbon-phosphorus ratio in a low phosphorus histosol. Soil Sci. 162: 275-282. [ Links ]
Anderson, M.J. 2005. PERMANOVA: a FORTRAN computer program for permutational multivariate analysis of variance. Department of Statistics, University of Auckland, Auckland, Nueva Zelanda. [ Links ]
Anderson, J.P.E & I. Ingram. 1993. Tropical soil biology and fertility. A handbook of methods. CAB International, Walingford, Reino Unido. [ Links ]
Anderson, J.P.E & K. Domsch. 1978. A physiological method for the quantitative measurement of microbial biomass in soil. Soil. Biol. Biochem. 10: 215-221. [ Links ]
Ashton, E.C., P. Hogarth & R. Ormond. 1999. Breakdown of mangroves leaf litter in a managed mangrove Forestin Peninsular Malasya. Hydrobiologia 413: 77-88. [ Links ]
Bandick, A & R. Dick. 1999. Field management effects on soil enzymes activities. Soil. Biol. Biochem. 31: 1471-1479. [ Links ]
Bastida, F., J.L. Moreno, T. Hernández & C. García. 2006. Microbiological degradation index for soil in semiarid climate. Soil Biol. Biochem. 38: 3463-3473. [ Links ]
Bellanger, B., S. Huon, P. Steinmann, F. Chabaux, F. Velasquez, V. Valles, K. Arn, N. Clauer & A. Mariotti. 2004. Oxic-anoxic conditions in the water column of a tropical freshwater reservoir (Peña-Larga dam, NW Venezuela). App. Geochem. 19: 1295-1314. [ Links ]
Bergstrom, D.W., C.M. Monreal & D.J. King. 1998. Sensitivity of soil enzyme activities to conservation practices. Soil Sci. Soc. Am. J. 62: 1286-1295. [ Links ]
Boto, K & J. Wellington. 1984. Soil characteristics and nutrient status in a northern Australian mangrove forest. Estuaries 7: 61-69. [ Links ]
Breemen, N.V & P. Buurman. 1998. Soil formation. Kluwer, Dordrecht, Holanda. [ Links ]
Caldwell, B. 2005. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 49: 637- 644. [ Links ]
Casida, L., D. Klein & T. Santoro. 1964. Soil dehydrogenase activity. Soil Sci. 98: 371-376. [ Links ]
Chacón, N., N. Dezzeo, M. Rangel & S. Flores. 2008. Seasonal changes in soil phosphorus dynamics and roots mass along a flooded tropical gradient in the lower Orinoco river, Venezuela. Biogeochemistry 87: 157-168. [ Links ]
Chen, R & R. Twilley. 1999. A simulation model of organic matter and nutrient accumulation in mangrove wetland soils. Biogeochemistry 44: 93-118. [ Links ]
Cintrón, G.M & Y.S. Novelli. 1984. Methods for studying mangrove structure, p. 91-113 In: S.C. Snedaker, J.G. Snedaker (eds.). The Mangrove Ecosystem: Research Methods. UNESCO, Paris, Isla de Francia, Francia. [ Links ]
Clarke, K & R. Warwick. 2001. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. PRIMER-E, Plymouth, Devon, Inglaterra. [ Links ]
Cundell, A.M., M. Brown, R. Stanford & R. Mitchell. 1979. Microbial degradation of Rhizophora mangle leaves inmersed in the sea. Estuar. Coast. Mar. Sci. 9: 281-286. [ Links ]
Dinesh, R., B.R. Shome & A.K. Bandyopadhyay. 1998. Enzyme activities and their relations to relevant soil properties. Curr. Sci. India 75: 510-512. [ Links ]
Dinesh, R., S.G. Chaudhuri, A.N. Ganeshamurthy & S.C. Pramanik. 2004. Biochemical properties of soils of undisturbed and disturbed mangroves Forestof South Andaman (India). Wetlands Ecol.Manag. 12: 309-320. [ Links ]
Dumontet, S., A. Mazzatura, C. Casucci & P. Perucci. 2001. Effectiveness of microbial indexes in discriminating interactive effects of tillage and crop rotations in a Vertic Ustorthents. Biol. Fert. Soils 34: 411-416. [ Links ]
Eivazi, F & M. Tabatabai. 1990. Factors affecting glucosidase and galactosidase activities in soils. Soil Biol. Biochem. 22: 891-897. [ Links ]
El-Tarabily, K.A. 2002. Total microbial activity and microbial composition of a mangrove sediment are reduced by oil pollution at a site in the Arabian Gulf. Can. J. Microbiol. 48: 176-182. [ Links ]
Ellison, J. 1998. Impacts of sediment burial of mangroves. Mar. Pollut. Bull. 37: 420-426. [ Links ]
FAO. 2007. The world´s mangroves 1980-2005. FAO Forestry Paper No. 153. Roma, Lazio, Italia. [ Links ]
Fondo Nacional de Investigaciones Agropecuarias (FONAIAP). 1990. Manual de métodos, procedimientos y referencias (Análisis de suelos para diagnósticos de fertilidad). Serie D No 26. Ministerio de Agricultura y Cría, Maracay, Vargas, Venezuela. [ Links ]
Frankenberger, W.T & F.T. Bingham. 1982. Influence of salinity on soil enzyme activities. Soil Sci. Soc. Am. J. 46: 1173-1177. [ Links ]
García, C & M.T. Hernández. 2000. Investigación y perspectivas de la enzimología de suelos en España. Consejo Superior de Investigaciones Científicas, CSIC-CEBAS, Murcia, España. [ Links ]
Ferreira, T.O., X.L. Otero, P. Vidal-Torrado & F. Macías. 2007. Effects of bioturbation by root and crab activity on iron and sulfur biogeochemistry in mangrove substrate. Geoderma 142: 36-46 [ Links ]
Hall, J.L. 2002. Cellular mechanism for heavy metal detoxification and tolerance. J. Exp. Bot. 53: 1-11. [ Links ]
Hayano, K & K. Tubaki .1985. Origin and properties of β-glucosidase activity of tomato-field soil. Soil Biol. Biochem. 17: 553-557. [ Links ]
Holguín, G & Y. Bashan. 2007. La importancia de los manglares y su microbiología para el sostenimiento de las pesquerías costeras, p. 239-252. In R. Ferrera Cerrato y A. Alarcón (eds.) Microbiología agrícola: hongos, bacterias, micro y macrofauna, control biológico, planta organismo. Trillas, México, México. [ Links ]
Holguín, G., P. Vazquez & Y. Bashan. 2001. The role of sediment microorganism in the productivity, conservation, and rehabilitation of mangrove ecosystem: an overview. Biol. Fertil. Soils 33: 265-278. [ Links ]
Kandeler, E & H. Gerber. 1988. Short-term assay of urease activity using colorimetric determination of ammonium. Biol. Fert. Soil 6: 68-72. [ Links ]
Kassen, A & D. Kleiner. 2001. Estimation of anaerobic microbial activities in soils by arginine ammonification and glucose-dependent CO2-production. Soil Biol. Biochem. 19: 683-686. [ Links ]
Lacerda, L.D., C.G. Rezende & D.M José. 1995. Mineral concentration of mangrove trees. Biotropica 17: 260-262. [ Links ]
Lacerda L.D., J.E. Conde, B. Kjerve, R. Álvarez-León, C. Alarcón & J. Polanía. 2002. American Mangroves, p. 1-62. In L.D. Lacerda (ed.). Mangrove Ecosystem function and management. Springer, Berlin. Alemania. [ Links ]
Ladd, J.N & J.H. Butler. 1972. Short-term assay of soils proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 4: 19-30. [ Links ]
Lentino, M & A.R. Bruni. 1994. Humedales costeros de Venezuela: situación ambiental. Sociedad Conservacionista Audubon de Venezuela. Caracas, Venezuela, p.113-135 In L. Lüttge (ed.). Mangroves. Salinity: Environment - Plants- Molecules. Kluwer, Dordrecht, Holanda. [ Links ]
Lüttge, U. 2002. Mangroves, p. 113-135. In A. Lauchli & U. Lüttge (eds.). Mangroves. Salinity: Environment - Plants - Molecules. Kluwer, Dordrecht, Holanda. [ Links ]
Marchand, C., E. Lallier-Verge & F. Baltzer. 2003. The composition of sedimentary organic matter in relation to the dynamic features of a mangrove-fringed coast in French Guiana. Estuar. Coast. Shelf Sci. 56: 119-130. [ Links ]
Marchand, C., F. Baltzer, E. Lallier-Vergès & P. Albéric. 2004. Pore water chemistry in mangrove sediments: relationship with species composition and developmental stages (French Guiana). Mar. Geol. 208: 361-381. [ Links ]
Monente, J.A. 1978. Estudio químico físico de la laguna de La Restinga. Mem. Soc. Ven. Cienc. Nat. La Salle 110: 227-307. [ Links ]
Muhammad, S., T. Müller & R.G. Joergensen. 2008. Relationships between soil biological and other soil properties in saline and alkaline arable soils from the Pakistani Punjab. J. Arid Environ. 72: 448-457. [ Links ]
Murphy, J. & J.P. Riley. 1962. A modified single solution method for the determination of phosphate in natural waters. Anal Chim. Acta 27: 31-39. [ Links ]
Pannier, F & R.F. Pannier. 1985. La vegetación de manglares de la cuenca del Lago de Maracaibo. Mem. Soc. Ven. Cienc. Nat. La Salle 60: 327-346. [ Links ]
Paolini, J & L.E. Sanchez-Arias. 2008. Comparative biochemical study of the rhizosphere of Rhizophora mangle and its associated species Cyperus sp. in the Ciénaga de Soledad (Colombia). Tasks Veg. Sci. 43: 79-84. [ Links ]
Pascual, J.A., C. Garcia, J.L. Hernandez, J. Moreno & M. Ros. 2000. Soil microbial activity as a biomarker of degradation and remediation processes. Soil Biol. Biochem. 32: 1877-1883. [ Links ]
Pezeshki, R.L. 2001. Wetland plant responses to soil flooding. Environ. Exp. Bot. 46: 299-312. [ Links ]
Poret-Peterson, A.T., B. Ji, E. Engelhaupt & J. Gulledge. 2007. Soil microbial biomass along a hydrologic gradient in a subsiding coastal bottomland forest: Implications for future subsidence and sea-level rise. Soil Biol. Biochem. 39: 641-645. [ Links ]
Ridd, P.V. & T. Stieglitz. 2002. Dry season salinity changes in arid estuaries fringed by mangroves and saltflats. Estuar. Coast. Shelf Sci. 54: 1039-1049. [ Links ]
Rivera-Monroy, V.H., R.R. Twilley, E. Medina, E.B. Moser, L. Botero, A.M. Francisco & E. Bullard. 2004. Spacial variability of soil nutrients in disturbed riverine mangroves Forestat diferents stages of regeneration in the San Juan river estuary, Venezuela. Estuaries 27: 44-57. [ Links ]
Rossel, D., J. Tarradellas, G. Bitton & J.L. Morel. 1997. Use of enzymes in ecotoxicology: a case for dehydrogenase and hydrolytic enzymes, p. 179-192. In J. Tarradellas, G. Bitton & D. Rossel (eds). Soil Ecotoxicology. CRC Lewis, Boca Raton, Florida, EEUU. [ Links ]
Sarathchandra, S. & K. Perrot. 1984. Assay of ß-glucosidase activity in soils. Soil Sci. 138: 15-19. [ Links ]
Sánchez-Arias, L.E. 2008. Efecto de la salinidad sobre la actividad microbiana y enzimática en suelos de manglar del Parque Nacional "Laguna de La Restinga" (Nueva Esparta, Venezuela). Ph.D tesis. Instituto Venezolano de Investigaciones Científicas, Centro de Estudios Avanzados, Caracas, Venezuela. [ Links ]
Sánchez-Páez, H., G. Ulloa-Delgado & R. Álvarez-León. 2000. Hacia la recuperación de los manglares del Caribe de Colombia. Impresos Panamericana, Santafé de Bogotá, D.C. Colombia. [ Links ]
Spalding, M., F. Blasco, C. Field. 1997. World Mangrove Atlas. International Society for Mangrove Ecosystems, Okinawa, Japón. [ Links ]
Schnürer, J. & T. Roswall. 1982. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 43: 1256- 1261. [ Links ]
Sherman, R.E., T.J. Fahey & R.W. Howarth. 1998. Soilplant interactions in a neotropical mangrove forest: iron, phosphorus and sulfur dynamics. Oecologia 115: 553-563. [ Links ]
Tabatabai, M.A. 1994. Soil Enzymes, p. 775-883. In R.W. Weaver, S. Angle & P. Bottomley (eds.). Methods of Soil Analysis: Microbiological and biochemical methods Book Series: 5. Soil Science Society of America, Madison, Wisconsin, EEUU. [ Links ]
Tabatabai, M.A. & J.M. Bremmer. 1969. Use of p-nitrophenylphospfate for assay of soil phosphatase activity. Soil Biol. Biochem. 1: 301-307. [ Links ]
Tripathi, S., A. Chakraborty, K. Chakrabarti & B.K. Bandyopadhya. 2007. Enzyme activities and microbial biomass in coastal soils of India. Soil Biol. Biochem. 39: 2840-2848. [ Links ]
Turner, B.L., R. Baxter & B.A. Whitton. 2002. Seasonal phosphatase activity in three characteristic soils of English uplands polluted by long-term atmospheric nitrogen deposition. Environ. Pollut. 120: 313-317. [ Links ]
US Department of Agriculture. 1950. Agriculture Handbook No. 60. Diagnosis and improvement of saline and alkali soils. U.S. Department of Agriculture, EEUU. [ Links ]
Vanhala, P.T & J.H. Ahtiainen. 1994. Soil respiration, ATP content and photobacterium toxicity test as indicators of metal pollution in soil. Environ. Tox. Water Qual. 9: 115-121. [ Links ]
Vásquez, P., G. Holguín, M. Puente, A. Lopez-Cortes & Y. Bashan. 2000. Phosphate-solubilizing microorganisms associated with the rizosphere of mangroves in a semiarid coastal lagoon. Biol. Fert. Soil 30: 460-468. [ Links ]
Yap, H. 2000. The case for restoration of tropical coastal ecosystems. Ocean Coast. Manage. 43: 841-851 [ Links ]
Yuan, B.C., Z.Z. Li, H. Liu, M. Gao & Y. Zhang. 2007. Microbial biomass and activity in salt affected soils under arid conditions. Appl. Soil Ecol. 35: 319-328. [ Links ]
Zahran, H., A.M. Moharram & H.A. Mohammad. 1992. Some ecological and physiological studies on bacteria isolated from salt-affected soils of Egypt. J. Basic Microbiol. 32: 405-413. [ Links ]


{kind=link}