











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.57 supl.1 San José Nov. 2009
The role of historical and local factors in determining species composition of the highland avifauna of Costa Rica and Western Panamá
Gilbert Barrantes
Escuela de biología, Universidad de Costa Rica, 11501-2060, San José, Costa Rica; gilbert.barrantes@gmail.com
Abstract: The formation of the mountain ranges of Costa Rica and western Panamá, as well as the cold climatic conditions that prevailed during the upper Pleistocene, played a crucial role in determining the bird species composition of the highlands in this region. Glacial conditions favored dispersal movements of bird species from the Andes, and from the Neartic region. Subsequent inter-glacial conditions reduced the connectivity between neotropical highlands (e.g., Talamanca-Andes), and between neotropical highlands and Neartic temperate region, isolating recently established populations from the ancestral populations, and promoting speciation. Within Costa Rica, the highland vegetation and the birds that occupied this vegetation possibly had a continuous distribution throughout all mountain ranges during glacial periods. This vegetation retreated to the summit of the mountains during inter-glacial periods, fragmenting the original continuous bird populations and forming "sky islands", which decrease in size from Talamanca mountains towards the northwestern ranges. The sizes of such islands of available habitat determine the number of highland birds present in each mountain range. Rev. Biol. Trop. 57 (Suppl. 1): 333-349. Epub 2009 November 30.
Key words: highland birds, biogeography, climate, endemism, Diglossa, Catharus, Piranga, Costa Rica.
Species composition in bird communities is influenced by both, historical and ecological factors (Vuilleumier & Simberloff 1980, Ricklefs 1987, Wiens 1989). The role of history in structuring communities has been frequently acknowledged, although few studies have specifically addressed its importance (Ricklefs & Cox 1972, Karr 1976a, Ricklefs 1978, Vuilleumier & Simberloff 1980, Lovette et al. 1998). More frequently, patterns of species occurrence have been attributed to past and contemporary interspecific interactions, such as competition and predation (MacArthur 1958, Diamond 1975, Cody 1978, Terborgh 1985, Martin 1992).
Regional species composition is likely the result of several factors acting in a more or less sequential order, e.g., colonization, speciation, and interspecific interactions (Wiens 1989). Species composition in a community is further modified by extinction, a continuos process that may be compensated by colonization and speciation (Mayr 1963, Wiens 1989). Colonization of a given location depends on speciesspecific dispersal ability and certain historical events (e.g., climatic change, formation of land bridges), that reduce or eliminate habitat discontinuities (Haffer 1970, 1974, 1987, Diamond 1984). Following colonization, speciation may later occur if isolation prevents gene flow between newly established and ancestral populations (Grant 1986, Hackett 1995). However, the importance of speciation in determining the species pool of a bird community has received little attention by community ecologists. On the contrary, interspecific interactions have frequently been considered to play a primary role in determining species membership in a bird assemblage (Lack 1947, Diamond 1975, Pulliam 1983). For example, competitive interactions have been proposed as a factor shaping bird communities through niche partitioning (Cody 1974, 1981). It is unlikely, however, that any single factor fully explains species composition in a bird community, and examination of present day communities is not complete without considering the role of history (Schluter & Ricklefs 1993).
Here, I examine how historical and ecological events have potentially interacted to determine species composition of the highland avifauna of Costa Rica and Western Panamá. This avifauna has a complex biogeographic origin and possesses the highest number of endemic species in Central America (Sánchez et al. 2004, Barrantes 2005). Highland bird assemblages in this region show discontinuous distributions due to the present day topography of highland sites and past historical events. I addressed two main questions in this study: (1) How did geological and vegetational changesassociated with Quaternary climatic fluctuations influenced the bird species composition in this region?; (2) How did local factors, specifically the area of highland forests (e.g., upper montane and subalpine forests), affected highland species composition and distribution among Costa Rican mountain ranges?
Materials and methods
Historical data: I used published information on geology, palaeoclimate, fossil record, and evolution of some groups of birds, to reconstruct possible dispersal and colonization events. These events are hypothesized to have had a major impact in determining species composition of the highland avifauna of Costa Rica and Western Panamá. I obtained geological information from Haffer (1974), Castillo (1984), Gómez (1986), and Coates & Obando (1996). Information on palaeoclimate and vegetation changes during the upper Pleistocene were described by Haffer (1974, 1987), Gómez (1986), Colinvaux (1996), and Webb & Rancy (1996). Changes in climate and expansion and contraction of highland forests on Talamanca mountains during late Quaternary were described by Hooghiemstra et al. (1992), and Islebe et al. (1995, 1996). Using information from Haffer (1974), Hooghiemstra et al. (1992), and Islebe et al. (1995, 1996) on vegetation patterns, I examined the role of vegetation changes on the current distribution of highland birds in Costa Rica. Using a digital elevation model (DEM) available at 30 arc-second scale for Costa Rica from USGS EROS web site and Geographic Information System software (Arc/INFO version 7.2, ArcView version 3.1, ESRITM, University of Missouri-St. Louis), I estimated the likely expansion and contraction of highland vegetation that occurred during the late Pleistocene and beginning of Holocene.
Dispersal events: I used information on fossil record (Olson 1985, Feduccia 1999), phylogenetic relationships (Hackett 1995, Burns 1998, Outlaw et al. 2003, Winker & Pruett 2006), and historical events (e.g., geological, vegetation, and climatic events) to reconstruct past dispersal events on the highlands of southern Central America (Haffer 1974, Hooghiemstra et al. 1992, Islebe et al. 1995, 1996).
Phylogenetic information on some extant taxa, including species present in Costa Rica and Panamá highlands, was used to infer the origin and dispersal routes of some bird lineages (Hackett 1995, Burns 1998, Outlaw et al. 2003, Winker & Pruett 2006). Although information on fossil record and pylogenetic relationships is incomplete, it still provides important information on the origin and expansion of ancient lineages.
Highland avifauna and current distribution:I relied on Stiles & Skutchs (1989) information to determine the distribution of highland bird species in the main mountain ranges of Costa Rica (Talamanca, Central Volcanic, Tilarán, and Guanacaste; Fig. 1). Highland species were defined by Wolf (1976), as those birds that occur above 2 600 m in the Talamanca mountain range (Appendix 1.1). The elevation of Costa Rican mountains decreases to the northwest (e.g, Tilarán and Guanacaste mountain ranges), where highest peaks lie between 1 500 and 2 000 m. However, climatic conditions have permitted the establishment of small fragments of the same highland vegetation present in the Talamancas at lower elevations on these more northerly mountains. Thus, highland habitats for birds
are present in all Costa Rican mountain ranges, although extension of these habitats decreases to the northwest.
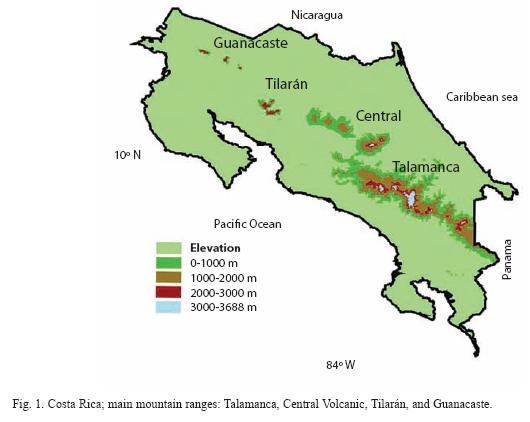
I described the effect of area of habitat available on species composition and distribution of highland, for non-endemic and endemic species (bird species exclusively restricted to Costa Rican and Western Panamá mountains), in different mountain ranges. The present area of highland vegetation (upper montane, subalpine, elfin, and paramo forests) of Costa Rican mountain ranges was calculated, using digitized maps (1:200,000, Lambert projection) and Geographic Information System software (ArcView, ESRITM, University of Missouri-St. Louis), to describe the effect of changes in area among mountain ranges on species richness of highland birds.
Results
Geological events
Early archipelagos: The recent formation of the Costa Rica-Panamá isthmus was the culmination of a long and intensive interaction of tectonic plates and volcanism (Haffer 1974, Castillo 1984, Gómez 1986, Coates & Obando 1996). The formation of the isthmus likely initiated during the upper Jurassic (Castillo 1984), with the collision of the Pacific and Caribbean plates to the South American plate (Coates & Obando 1996). These tectonic dynamics, and an intensive submarine volcanism that extended from the upper Jurassic throughout most of the Cretaceous, resulted in a progressive uplift of an incipient archipelago between South and North America (Castillo 1984, Gómez 1986). At the end of the Cretaceous, the majority of the islands disappeared under the ocean, possibly as a result of erosion (Castillo 1984). A new uplifting event, however, raised an archipelago in this ocean gap by the end of the Paleocenebeginning of the Eocene, approximately 50 million years ago (mya), (Castillo 1984).
Costa Rica - Chiriquí highlands: Duringthe Middle and Late Oligocene (ca. 30 mya) an orogenic folding event caused by subduction of the Cocos ridge beneath the Caribbean Plate developed into the incipient Talamanca mountain range (Castillo 1984, Coates & Obando 1996). Uplifting continued until the Upper Miocene, 12 - 7 mya (Castillo 1984). The Miocene Talamancas consisted of a group of low-hilly islands separated from each other by some shallow marine connections (Coates & Obando 1996). The tectonic activity during the Pliocene possibly culminated with the formation of the first continental land bridge between South and North America about 3.5 - 3.1 mya (Haffer 1974, Castillo 1984, Coates & Obando 1996).
The end of the Pliocene-beginning of the Pleistocene epoch marked the start of the last tectonic and volcanic events that gave to the highlands of Panamá and Costa Rica their present form. The last uplift of Talamanca mountains was initiated at the beginning of the Pleistocene (ca. 2 mya). This tectonic process was accompanied by a very intensive volcanic activity that eventually resulted in the formation of Tilarán, Central Volcanic, and Guanacaste mountain ranges in Costa Rica (Castillo 1984) and the Chiriquí mountain range in Panamá (Gómez 1986). These tectonic and volcanic activities extended throughout much of the Pleistocene epoch.
Historical climatic changes
Tertiary climate: In general, it is accepted that from 80 to 40 mya, previous to the Eocene, the average global temperature was higher and more stable than at present (Haffer 1974, Gómez 1986). Thus, even during the early Tertiary, a uniform and generally warm climate reached farther north and south than the present tropics (Haffer 1974). The climate began to shift to lower temperatures during the Eocene (53 mya) and Oligocene (33 mya) (Emiliani 1956, Tanner 1968, Haffer 1974, Gómez 1986). Reduction in temperature during the Tertiary affected mainly the polar regions with little, if any, effect on the Neotropical zone (Dorf 1959, Haffer 1974, Tanner 1968).
Quaternary climate and vegetation changes: Climatically, the Quaternary was characterized by the alternation of cool and generally drier periods with warm and wet periods (Emiliani 1956, Weyl 1957, Berggren et al. 1967, Tanner 1968, Haffer 1974, Webb & Rancy 1996). Cool and relatively dry climatic conditions prevailed during the late Plioceneearly Pleistocene. As glaciers advanced at high latitudes, in the northern hemisphere, the tropics became cooler and drier (Webb & Rancy 1996). During the early Pleistocene, the recently emerged lowland of the Isthmus of Costa Rica-Panamá likely held no forest, or forest was very reduced in its distribution (Haffer 1974, Webb & Rancy 1996), and the low altitude of the Talamanca mountains precluded formation of glaciers in the region (Haffer 1974, 1987).
During the middle Pleistocene climate progressively turned more tropical as temperature and relative humidity increased (Haffer 1974, Gómez 1986, Webb & Rancy 1996). The wet and warm conditions that prevailed during this interglacial period resulted in the expansion of South American tropical rain forests into Central America (Webb & Rancy 1996). Thus, tropical forests became the dominant vegetation in the lowlands and mountains on the southern extreme of Middle America (Gómez 1986). A shift in climatic conditions, however, occurred at the end of the Pleistocene (Weyl 1957, Haffer 1974, Gómez 1986, Webb & Rancy 1996). Lower temperature and relative humidity likely caused a drastic reduction in the extent of lowland wet forests in tropical regions (Weyl 1957, Haffer 1974, 1987). As a consequence, arid or semiarid vegetation apparently substituted rain forests in large extensions of the Neotropics (Haffer 1974, Gómez 1986, Webb & Rancy 1996). These changes in vegetation also affected the distribution of the avifauna (Haffer 1974, 1987). The climatic conditions prevailing by the end of the Pleistocene led to the formation of extensive glaciers on high tropical mountains (e.g., the Andes; Haffer 1974). Less extensive glaciers also occurred on the highest peaks of Talamanca mountain range (Weyl 1957, Haffer 1974, Gómez 1986). In these high tropical mountains (e.g., the Andes, the Talamancas), the declining temperature compressed downward the altitudinal life zones along the mountain slopes as much as 700 m (Haffer 1974, Gómez 1986). Thus, subalpine and montane forests had a more continuous distribution than today (Haffer 1974, Gómez 1986). The cold phase at the end of the Pleistocene was characterized by the alternation of relatively short humid-warm periods followed by dry-cold periods. Such climatic fluctuations affected the extension of highland forests, and consequently, the connectivity between Neotropical mountain ranges and between tropical and temperate regions, particularly during the last 100 000 years of the Pleistocene epoch (Haffer 1974, Bermingham et al. 1992, Colinvaux 1996).
Post-Pleistocene climate: The climate at the end of the Pleistocene became gradually wetter and warmer (Haffer 1974, 1987, Gómez 1986, Webb & Rancy 1996). This shift in climatic conditions marked the beginning of the present interglacial period. When climatic conditions turned more tropical after the Pleistocene, lowland forests re-expanded and replaced the arid and semiarid vegetation that had covered large tracts in Central and South America. On the mountain ranges, the prevailing climatic conditions forced highland vegetation to retreat towards the summit of mountain ranges (Haffer 1974, Gómez 1986, Hooghiemstra et al. 1992, Islebe et al. 1995, 1996). Consequently, paramo, puna, subalpine, and upper montane forests have a more discontinuous distribution at present than during the upper Pleistocene, and in some cases these habitats are truly "sky islands" (Mayr & Diamond 1976, Vuilleumier & Simberloff 1980). This process of expansion- contraction of highland vegetation likely occurred several times during the upper Pleistocene (Haffer 1974, Hooghiemstra et al. 1992, Islebe et al. 1995, 1996, Colinvaux 1996).
Dispersal events
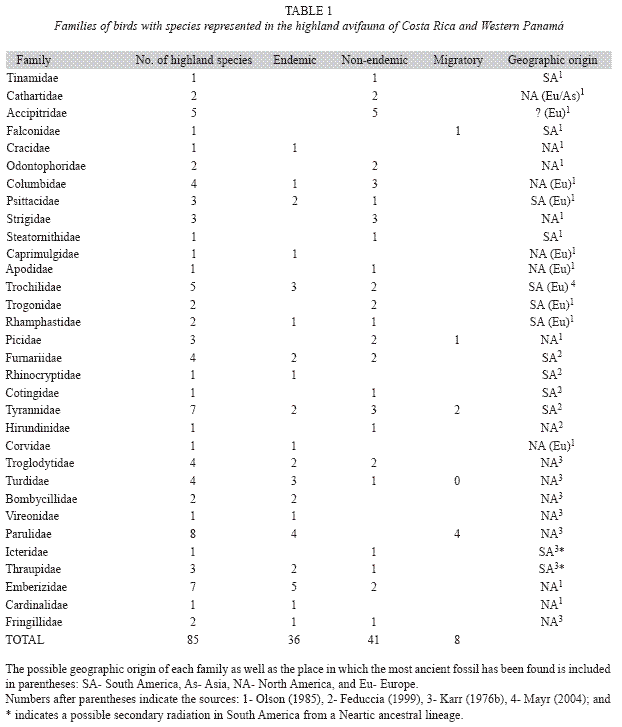
The Costa Rican highland avifauna consists of 85 species, from which 37 are endemic to the region (Table 1). These species have affinities to at least three different geographic regions (Appendix 1), including those closely related to South American lineages, those with clear Neartic derivation, and those that may have originated from lowland Central American ancestors (Stiles 1983).
Knowledge on the geological events that gave origin to the southern part of Middle America and historical climatic changes provide a basis for hypotheses regarding the main dispersal events that gave origin to this highland avifauna (Haffer 1970, 1974, Stiles 1983, Gómez 1986, Webb & Rancy 1996, Feduccia 1999). The scarce and incomplete fossil record of birds prevents a more precise determination of the time period in which these dispersal events occurred (Olson 1985, Feduccia 1999). Nonetheless, it is possible to hypothesize about three main events of dispersal: (1) at the end of the Pliocene-beginning of the Pleistocene, (2) at the Middle Pleistocene, and, (3) at the end of the Pleistocene.
End of Pliocene-beginning of Pleistocene dispersal: The first, important, dispersal of birds into the emerging lands of southern Central America likely happened during the upper Pliocene- lower Pleistocene (ca. 3 mya), with the formation of the first land bridge between South and North America (Haffer 1970, 1974, Rich & Rich 1983, Gómez 1986, Webb & Rancy 1996). Unfortunately, no fossil evidence exists of those groups that may have colonized the emerging lands. Considering that the climatic conditions during such time were cooler and drier than at present, Webb & Rancy (1996) suggested that savanna type vegetation dominated the landscape of this part of Central America. This argument is supported by the savanna-type mammal fauna that arrived to the southern extreme of Middle America and continued moving into South America when the land bridge was finally formed (Rich & Rich 1983, Webb & Rancy 1996). It is also possible that some bird groups, such as corvids and woodpeckers, well adapted to an open or semiopen landscape, arrived from North America to these latitudes during this period. The Costa Rica-Panamá mountain ranges were far from being completely developed at the Pliocenebeginning of Pleistocene time, so that the early arrivals did not become an integral part of the highland avifauna. Yet, some of these Nearctic groups may have undergone secondary radiations in Central and South American forests (e.g., New World jays, forest woodpeckers respectively), and these new lineages likely colonized the southern Central American highlands, as proposed by Howell (1969).
Middle Pleistocene dispersal: The tropical climate that prevailed during most of the middle Pleistocene resulted in the longest interglacial period of the Quaternary (Haffer 1974, Webb & Rancy 1996). During the middle Pleistocene, when elevation of mountains (e.g., Talamanca, Andes) were still increasing, and global climate turned warmer and wetter, South American tropical rain forest birds likely extended their range into the recently emerged terrain of Costa Rica-Panamá (Webb & Rancy 1996) (Table 1). Dispersers from tropical South American were likely constituted by lowland neotropical (e.g., tinamous, manakins) and pantropical lineages (e.g., parrots, trogons). Furthermore, groups that had undergone secondary radiations in South America may have also arrived in the same period (e.g., forest woodpeckers, thrushes).
High montane areas were relatively well isolated in the neotropics during the middle Pleistocene (Haffer 1974); however, it is possible that some South American highland and Neartic (ancestral) species colonized the high mountains of Costa Rica and Western Panamá. Dispersal among high montane "islands" through (or over) lowland forests has been reported for the montane avifauna of New Guinea (Mayr & Diamond 1976). Also, the presence of solitares (Myadestes spp.), and trogons (Priotelus spp.) on the Antillean islands(Raffaele et al. 1998), confirm the dispersal capabilities of even "poor fliers".
Dispersal during the Pleistocene glacial periods: The cold climatic conditions that dominated at the end of the Pleistocene (especially the last 100 000 yrs of this epoch), caused a reduction in lowland tropical forest and expansion of more temperate-like vegetation (i.e., paramo, subalpine forest). Hence, the climate and vegetation that prevailed at the cold phases of this epoch again favored the dispersal of Neartic bird groups into the highlands of southern Middle America. The genus Junco and ancestors of Ptilogonatinae (Bombycillidae), are possible examples of this Neartic component that arrived during the upper Pleistocene (Miller 1941, Sibley & Ahlquist 1990). Some South American groups that were already adapted to highland environments in the Andes also are hypothesized to have reached the highlands of Costa Rica and Western Panamá during this cold period. For example, the genera Diglossa and Scytalopus, both of them diverse groups of the high Andes, as well as Margarornis and Pseudocolaptes are examples of groups that likely extended their distribution from the South America highlands during cool and dry glacial phases (Hackett 1995). Finally, some highland birds may have also evolved from middle elevation, resident species that became adapted to the cold environments of the Costa Rica-Panamá mountains.
Distribution and phylogenetic evidence for the origin of the highland avifauna of Costa Rica – Panamá
Considering the 32 families of birds represented in the highlands of Costa Rica and Western Panama, approximately 50% were present in South and 50% in North American, previously to the formation of Costa Rica-Panamá isthmus. Most South American lineages consist of non-passerine pantropical groups and suboscine bird groups, while North American lineages consist of widely distributed Holarctic groups and oscine families. Oscine birds likely invaded North America through Beringia during the late Tertiary, Oligocene/early Miocene (Feduccia 1999, Barker et al. 2004). Yet, most species in this group likely remained isolated in the North American continent until the Great Faunal American Interchange occurred during the late Pliocene (Feduccia 1999, Webb & Rancy 1996), when South and North America were finally connected by a landbridge (Coates & Obando 1996). Some immigrant groups underwent large secondary radiations in both continents after the Great Interchange. In South America, groups such as tanagers, oropendulas and relatives, and forest woodpeckers (e.g., Celeus, Veniliornis) diversified extensively (Howell 1969, Feduccia 1999). Similarly, North American flycatchers, the most diverse group in South America, underwent an extensive speciation (Fitzpatrick 1985). The rapid speciation of numerous resident and immigrant groups that occurred on both continents (South America and North America) during the late Pliocene and Pleistocene was likely the result of drastic climatic fluctuations and mountain uplifts (Haffer 1974, 1987). These changes in climate and geomorphology created new habitats, when changes in vegetation occur, and consequently new opportunities for speciation (Mayr 1963). Expansion of montane forests also increased the probability of some groups of birds (including recently evolved species) to gradually disperse into Costa Rica-Panamá highlands (Haffer 1967, Mayr & Diamond 1976).
The scarce phylogenetic information supports the complex biogeographic origin of Costa Rica-Panamá highland avifauna. Some phylogenetic hypotheses include information on highland species: Diglossa flowerpiercers (Hackett 1995), Catharus thrushes (Outlaw et al. 2003, Winker & Pruett 2006), and Piranga genus (Burns 1998). Although phylogenies give relatively accurate information on the evolutionary history of these groups, extinctions and absence of species in some clades canobscure their cladogenesis and lead to erroneous interpretation of patterns of evolution.
Flowerpiercers: Diglossa plumbea is an endemic member of the highland avifauna of Costa Rica and Western Panamá. The genus is South American in origin and likely dispersed into Central America during a glacial period of the Pleistocene (Fig. 2A, Hackett 1995). D. plumbea form part of the baritula complex which includes two more species, D. baritula, an endemic species to the mountains of Mexico, Guatemala, and Honduras, and the basal member of the complex, D. sittoides. Diglossa sittoides is found in the highland regions of South America (Hackett 1995). Allozyme distance data indicate that divergence among D. sittoides, D. plumbea, and D. baritula occurred during the Pleistocene (during the last two million years, Hackett 1995). This information suggests that Central American ancestral populations became isolated from South American populations, and subsequently Central American populations gave rise to two species, D. baritula and D. plumbea.
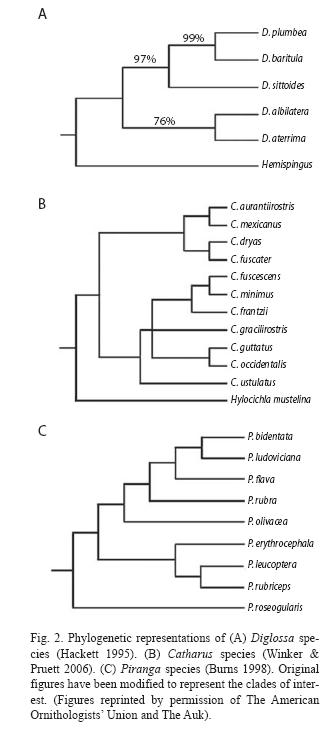
Catharus thruses: C. gracilirostris is another endemic species of Costa Rica-Panamá highlands. The distribution of the genus (12 species) is primarily North and Middle American. Five out of the twelve species are migratory, breeding in North America and wintering in Middle and South America (Peterson & Chalif 1973, Ridgely & Tudor 1989, Stiles & Skutch 1989). The phylogenetic work on the genus Catharus presented by Outlaw et al. (2003) and Winker & Pruett (2006) indicates that the ancestor of C. gracilirostris was, more likely, a low-middle land, resident species (Fig. 2B), suggesting that C. gracilirostris evolved by adaptation of a possibly middle elevation ancestor to the prevailing conditions during the glacial periods in the upper Pleistocene.
Piranga: P. bidentata is a resident, nonendemicspecies of the highlands from Costa Rica and Western Panamá to Mexico (Sibley & Monroe 1990). The geographic distribution of this genus and likely its origin is centered in Mexico and Central America (Burns 1998). Using maximum likelihood approach to reconstruct the phylogeny of Piranga, Burns (1998) argued that P. bidentata has recently evolved possibly from a latitudinal migratory lineage (Fig. 2C), since this species is terminal in a clade that embraces most of the migratory species.
These examples show the evolutionary and biogeographical complexity of the highland avifauna of this region. It is apparent that the conformation of this highland avifauna was mediated by the climatic changes that occurred during the last part of the Pleistocene, but the processes of adaptation and speciation were extremely complex
Highland avifauna in Costa Rica
The climatic changes (increase in temperature and relative humidity) that occurred during the Pleistocene interglacial periods had two main consequences for the highland avifauna of Costa Rica and Western Panamá. First, these changes severed connections between montane avifaunas of Costa Rica-Western Panamá and South America, when montane forests retreated to higher elevations. Second, within Costa Rica, the highland avifauna became fragmented along the different mountain ranges (Talamanca, Central Volcanic, Tilarán, and Guanacaste).
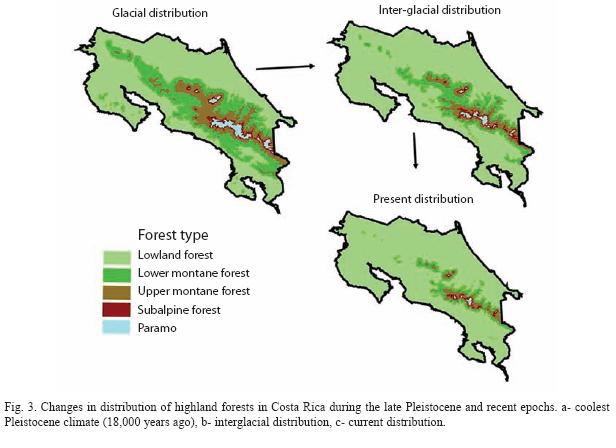
During the Pleistocene glacial phases, subalpine forest in Talamanca mountains descended to 2 200 m, 700 m below its current limit (Fig. 3A). Likewise, upper montane forest likely extended until 1 400 m and lower montane to ca. 500 m (Gómez 1986, Haffer 1987). Similar shifts in vegetation are expected to have occurred in all Costa Rican mountain ranges during glacial periods. Hence, subalpine and upper montane forests were likely connected, or nearly so, throughout all mountain ranges during glacial periods (Fig. 3A). Therefore, the distribution of highland birds were more likely continuous along Talamanca, Central Volcanic, Tilarán, and perhaps, Guanacaste mountains. Colonization and establishment of highland birds in Guanacaste mountain range, however, may have been restricted by the intense activity of its volcanos during the Pleistocene and beginning of the Holocene epochs (Gómez 1986), and only later would highland birds have reached this mountain range.
The tropical climate that prevailed during the Pleistocene interglacial periods caused the contraction of highland forests (Fig. 3B-C). This shift in vegetation likely caused the fragmentation of highland bird populations. Therefore, the distribution of several highland birds consisted of isolated populations restricted to the highest portion of different mountain ranges. These changes in habitat extension and fragmentation likely resulted in the present day nested subset of highland bird species distributions.
All highland species, including nonendemic and endemic species, are present in the Talamanca mountain range. However, the number of non-endemic and endemic (Fig. 4) species decreases in a nested fashion from Talamanca to Guanacaste mountains. A possible explanation for this decline in species richness from Talamanca to Guanacaste mountains is the reduction of potential habitat available (mainly paramo, subalpine and upper montane forests) along these mountain ranges (Fig. 3).
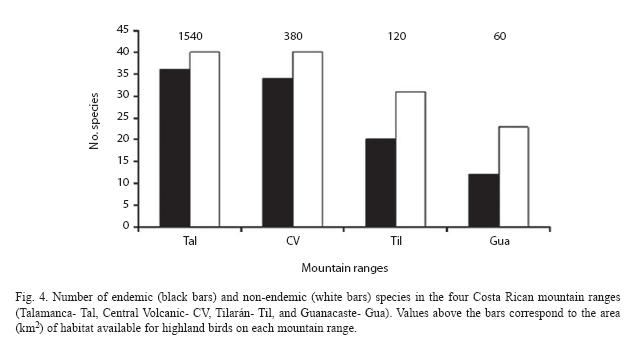
Much of the complex composition of Costa Rica and Western Panamá highland avifauna is explained by the geographical location of Costa Rica-Panamá (between South and North America), the geological events that culminated with the formation of the first landbridge between South and North America, the formation of Talamanca-Chiriquí mountain range, and the climatic changes during the late Tertiary and Quaternary. Particularly, the climatic changes permitted diverse groups of birds to reach the isolated Talamanca-Chiriquí mountains, when montane and temperate vegetation descended to lowlands during cool phases of the Pleistocene (Haffer 1974, Mayr & Diamond 1976, Hacket 1995).
However, it is notable the absence of the rich South American paramo avifauna on Talamanca-Chiriqui mountains (Vuilleumier & Simberloff 1980, Fjeldså & Krabbe 1990). The following arguments are possible explanations to the absence of the Andean paramo avifauna in Central America: 1- Even during the coldest Pleistocene periods, the paramo was very fragmented and isolated to allow South American paramo species to reach highlands in Costa Rica and Western Panamá. 2- Although the climatic changes during the glacial periods were dramatic, such changes could not had been sufficient to allow birds adapted to the coldest conditions on the Andes, to overcome the differences in climatic conditions predominant over the large, lowland extensions that separated the Andes from the Talamancas.
The composition of the assemblage of highland birds of Costa Rica and Western Panamá has resulted from the combination of historical events and area of available habitat (highland forests). The biogeographic complexity of this highland avifauna began with the formation of the landbridge that connected South and North America during the late Pliocene-beginning of the Pleistocene. The formation of this connection was possibly followed by the first great avifaunal interchange between both continents. Quaternary climatic changes promoted a massive movement of South American birds into Central America, when South American tropical forests expanded into the recently formed Costa Rica and Panamá. During the upper Pleistocene, temperatures began to decline, and this climatic shift resulted in the last glacial period. Changes in vegetation associated with such climatic changes allowed Neartic and high Andean birds to reach to the highlands of Costa Rica and Western Panamá.
Within the region, climatic changes promoted the expansion and contraction of suitable habitats for highland birds. When climate became warmer during the interglacial periods, highland forests retreated to the summit of high mountains resulting in fragmentation of the original populations. Area of these isolates seemed to have played an important role in determining the pattern of species richness and composition within the region (Costa Rica and Western Panamá highlands). These highland forest fragments include a particular composition of plants (Vargas & Sánchez 2005), and possibly of arthropods (Kappelle 2005) that serve as food resources to highland birds (Barrantes & Loiselle 2002, Barrantes 2005). Hence, in addition to the area of potential available habitat, changes in food resources associated to changes in mountains elevation may also be important in explaining the geographic pattern of species richness, particularly for endemic highland birds. In summary, historical and contemporary factors have to be considered to understand the underlying mechanisms that determine the biogeographic patterns in a given organismal assemblage (Prance 1987, Brown 1987, Monge-Nájera 1994).
Acknowledgments
I thank Robert E. Ricklefs, John Blake, Bette Loiselle, and J.A. Vargas for valuablecomments on the manuscript.
Resumen
La formación de las cordilleras en Costa Rica y oeste de Panamá, así como las frías condiciones climáticas que prevalecieron durante la última parte del Pleistoceno, jugaron un papel muy importante en determinar la composición de las especies de aves en las tierras altas de esta región. Las condiciones glaciales favorecieron la dispersión de aves desde los Andes y desde la región Neártica. Las subsecuentes condiciones inter-glaciales redujeron la conectividad entre las zonas altas neotropicales (e.g., Talamanca-Andes) y entre las zonas altas neotropicales y la región templada Neártica, aislando así las poblaciones recientemente establecidas, de las poblaciones ancestrales, fomentando la especiación. Dentro de Costa Rica, la vegetación de zonas altas así como las aves que ocupaban esta vegetación, posiblemente tuvieron una distribución continua a lo largo de todas las cordilleras durante os periodos glaciales. Esta vegetación quedó restringida a la cumbre de las montañas durante los periodos inter-glaciales, fragmentando las poblaciones originales continuas de aves, lo que formó islas en las cumbres de las montañas, las cuales decrecen en tamaño desde la cordillera de Talamanca hacia las cordilleras en el noroeste del país. El tamaño de esas islas con habitat disponible determina el número de especies de aves de zonas altas presente en cada cordillera.
Palabras clave: aves de tierras altas, biogeografía, distribución, endemismo, Costa Rica.
Received 30-VI-2007. Corrected 09-X-2008. Accepted 18-XI-2008.
References
Barker, F.K., A. Cibois, P. Schikler, J. Feinstein & J. Cracraft. 2004. Phylogeny and diversification of the largest avian radiation. Proc. Nat. Acad. Sci. 101: 1104-1045. [ Links ]
Barrantes, G. 2005. Historia natural de las aves del páramo costarricense, p. 521-532. In M. Kappelle & S. Horn (eds.). Páramos de Costa Rica. INBio, Heredia, Costa Rica. [ Links ]
Barrantes, G. & B.A. Loiselle. 2002. Reproduction, habitat use, and natural history of Phainoptila melanoxantha. Ornitol. Neotrop. 13: 121-136. [ Links ]
Berggren, W.A., J.D. Phillips, A. Bertels & D. Walls. 1967. Late Pliocene-Pleistocene stratigraphy in deep-sea cores from the south-central North Atlantic. Nature 216: 253-254. [ Links ]
Bermingham, E., S. Rohwer, S. Freeman & C. Wood. 1992. Vicariance biogeography in the Pleistocene and speciation in North American wood warblers: a test of Mengels model. Proc. Natl. Acad. Sci. 89:6624-6628. [ Links ]
Brown, K.S. 1987. Quaternary history of tropical America, p. 66-99. In T.C. Whitmore & G.T. Prance (eds.). Biogeography and Quaternary history in tropical America. Clarendon, Oxford, USA. [ Links ]
Burns, K. 1998. Molecular phylogenetics of the genus Piranga: implications for biogeography and the evolution of morphology and behavior. Auk 115: 621-634. [ Links ]
Castillo, M.R. 1984. Geología de Costa Rica. Universidad de Costa Rica, San José, Costa Rica. [ Links ]
Coates, A.G. & J.A. Obando. 1996. The geologic evolution of the Central American isthmus, p. 21-56. In J.B.C. Jackson, A.F. Budd & A.G. Coates (eds.). Evolution and environment in tropical America. University of Chicago, Chicago, USA. [ Links ]
Cody, M.L. 1981. Habitat selection in birds: the roles of vegetation structure, competition and productivity. BioScience 31: 107-111. [ Links ]
Cody, M.L. 1978. Habitat selection and interspecific territoriality among the sylviid warblers of England and Sweden. Ecol. Monogr. 48: 351-396. [ Links ]
Cody, M.L. 1974. Competition and structure of bird communities. Princeton University, Princeton, USA. [ Links ]
Colinvaux, P.A. 1996. Quaternary environmental history and forest diversity in the neotropics, p. 359-405. In J.B.C. Jackson, A.F. Budd & A.G. Coates (eds.). Evolution and environment in tropical America. University of Chicago, Illinois, USA. [ Links ]
Diamond, J.M. 1984. Distribution of New Zealand birds on real and virtual islands. New Zeal. J. Ecol. 7: 37-55. [ Links ]
Diamond, J.M. 1975. Assemblage of species communities, p. 342-444. In M.L. Cody & J.M. Diamond (eds.). Ecology and evolution of communities. Harvard University, Massachusetts, USA. [ Links ]
Dorf, E. 1959. Climatic changes of the past and present. Contrib. Mus. Paleontol. U. Mich. 13: 181-210. [ Links ]
Emiliani, C. 1956. Oligocene and Miocene temperatures of the equatorial and Subtropical Atlantic Ocean. J. Geol. 64: 281-288. [ Links ]
Feduccia, A. 1999. The origin and evolution of birds. Yale University, London, England. [ Links ]
Fitzpatrick, J.W. 1985. Form, foraging, and adaptive radiation in the Tyrannidae. Ornith. Monogr. 36: 447-470. [ Links ]
Fjeldså, J. & N. Krabbe. 1990. Birds of the high Andes. Zoological Museum of Copenhagen and Apollo Books, Svendborg, Denmark. [ Links ]
Gómez, L.D. 1986. Vegetación de Costa Rica. Apuntes para una biogeografía costarricense. Universidad Estatal a Distancia, San José, Costa Rica. [ Links ] Grant, P.R. 1986. Ecology and evolution of Darwins finches. Princeton University Press, Princeton, USA. [ Links ]
Hackett, S.J. 1995. Molecular systematics and zoogeography of flowerpiercers in the Diglossa baritula complex. Auk 112: 156-170. [ Links ]
Haffer, J. 1987. Quaternary history of tropical America, p. 1-18. In T.C. Whitmore & G.T. Prance (eds.). Biogeography and Quaternary history in tropical America. Clarendon, Oxford, England. [ Links ]
Haffer, J. 1974. Avian speciation in tropical South America. Nut. Ornith. Club No. 14. [ Links ]
Haffer, J. 1970. Geologic-climatic history and zoogeographic significance of the Uraba region in northwestern Colombia. Caldasia 10: 603-636. [ Links ]
Hooghiemstra, H., A.M. Cleef, G. Noldus & M. Kappelle. 1992. Upper Quaternary vegetation dynamics and palaeoclimatology of the La Chonta bog area (Cordillera de Talamanca, Costa Rica). J. Quaternary Sci. 7: 205-225. [ Links ]
Howell, T.R. 1969. Avian distribution in Central America. Auk 86: 293-326 [ Links ]
Islebe, G.A., H. Hooghiemstra & R. Van T. Veer. 1996. Holocene vegetation and water level history in two bogs of the Cordillera de Talamanca, Costa Rica. Vegetatio 124: 155-171. [ Links ]
Islebe, G.A., H. Hooghiemstra & K. Van Der Borg. 1995. A cooling event during the younger Dryas Chron in Costa Rica. Palaeogeogr. Palaeocl. 117: 73-80. [ Links ]
Kappelle, M. 2005. Insectos de los páramos de Costa Rica, p. 493-499. In M. Kappelle & S. Horn (eds.). Páramos de Costa Rica. INBio, Heredia, Costa Rica. [ Links ]
Karr, J.R. 1976a. Within- and between–habitat avian diversity in Africa and neotropical lowland habitats. Ecol. Monogr. 46: 457-481. [ Links ]
Karr, J. R. 1976b. Birds of Panamá: Biogeography and ecological dynamics, p. 77-93. In W.G. DArcy & M. Correa (eds.). The botany and natural history of Panamá. Braun-Brunfield Inc., Michigan, USA. [ Links ]
Lack, D. 1947. Darwins finches. Cambridge University, Cambridge, USA. [ Links ]
Lovette, I., E. Bermingham, G. Seutin, & R.E. Ricklefs. 1998. Evolutionary differentiation in the three endemic West Indian warblers. Auk 115: 890-903. [ Links ]
MacArthur, R.H. 1958. Population ecology of some warblers in northeastern coniferous forests. Ecology 39: 599-619. [ Links ]
Martin, J.L. 1992. Expansion in an insular bird community: an autoecological perspective. J. Biogeogr. 19: 375-381. [ Links ]
Mayr, E. 1963. Animal species and evolution. Harvard University, Cambridge, USA. [ Links ]
Mayr, G. 2004. Old world fossil record of modern-typehummingbirds. Science 304: 861-864. [ Links ]
Mayr, E. & J.M. Diamond. 1976. Birds on islands in the sky: origin of the montane avifauna of northern Melanesia. P. Natl. Acad. Sci. USA 73: 1765-1769. [ Links ]
Miller, A. 1941. Speciation in the avian genus Junco. University of California, California, USA. [ Links ]
Monge-Nájera, J. 1994. Ecological Biogeography in the Phylum Onychophora. Biogeographica 70: 111-123. [ Links ]
Olson, S.L. 1984. The fossil record of birds, p. 80-238. In D.S. Farmer, J.R. king & K.C. Parkes (eds.). Avian biology. Volume VI. Academic, New York, USA. [ Links ]
Outlaw, D.C., G. Voelker, B. Mila & D.J. Girman. 2003. Evolution of long-distance migration and historical biogeography of Catharus thrushes: a molecular phylogenetic approach. Auk 120: 299-310. [ Links ]
Peterson, R.T. & E.L. Chalif. 1973. A field guide to Mexican birds. Houghton Mifflin Company, Boston, USA. [ Links ]
Prance, G.T. 1987. Quaternary history of tropical America, p. 46-65. In T.C. Whitmore & G.T. Prance (eds.). Biogeography and Quaternary history in tropical America. Clarendon, Oxford, USA. [ Links ]
Pulliam, H.R. 1983. Ecological community theory and the coexistence of sparrows. Ecology 64: 45-52. [ Links ]
Raffaele, H., J. Wiley, O. Garrido, A. Keith & J. Raffaele. 1998. A guide to the birds of the West Indies. Princeton University, Princeton, New Jersey, USA. [ Links ]
Rich, P.V. & T.H. Rich. 1983. The Central American dispersal route: biotic history and paleogeography, p. 12-34. In D.H. Janzen (ed.). Costa Rican natural history. University of Chicago, Illinois, USA. [ Links ]
Ricklefs, R.E. 1987. Community diversity: relative roles of local and regional processes. Science 235:167-171. [ Links ]
Ricklefs, R.E. 1978. Stage of taxon cycle, habitat distribution, and population density in the avifauna of the West Indies. Oikos 29: 60-66. [ Links ]
Ricklefs, R.E. & G.W. Cox. 1972. Taxon cycles in the West Indian avifauna. Am. Nat. 106: 195-219. [ Links ]
Sánchez, J.E., G. Barrantes & F. Durán. 2004. Distribución, ecología y conservación de las aves de la cuenca del río Savegre, Costa Rica. Brenesia 61: 63-93. [ Links ]
Schluter, D. & R.E. Ricklefs. 1993. Species diversity: an introduction, p. 1-10. In R.E. Ricklefs, & D.Schluter (eds.). Species diversity in ecological communities. Historical and geographical perspectives. University of Chicago, Illinois, USA. [ Links ]
Sibley, C.G. & J.E. Ahlquist. 1990. Phylogeny classification of birds. A study in molecular evolution. Yale University, London, England. [ Links ]
Sibley, C.G. & B.L. Monroe, Jr. 1990. Distribution and taxonomy of birds of the world. Yale University, London, England. [ Links ]
Stiles, F.G. 1983. Birds. Introduction, p. 502-530. In D.H.Janzen (ed.). Costa Rican natural history. University of Chicago, Illinois, USA. [ Links ]
Stiles, F.G. & A.F. Skutch. 1989. A guide of the birds of Costa Rica. Cornell University, New York, USA. [ Links ]
Tanner, W.F. 1968. Tertiary sea-level symposium – Introduction. Palaeogeogr. Palaeocl. 5: 7-14. [ Links ]
Terborgh, J. 1985. Habitat selection in Amazonian birds, p. 311-338. In M.L. Cody (ed.). Habitat selection in birds. Academic, New York, USA. [ Links ]
Vargas, G. & J.J. Sánchez. 2005. Plantas con flores de los páramos de Costa Rica y Panama: el paramo istmico, p. 399-435. In M. Kappelle & S. Horn (eds.). Páramos de Costa Rica. INBio, Heredia, Costa Rica. [ Links ]
Vuilleumier, F. & D. Simberloff. 1980. Ecology versus history as determinants of patchy and insular distributions in high Andean birds. Evol. Biol. 12: 235-379. [ Links ]
Webb, S.D. & A. Rancy. 1996. Late Cenozoic evolution of the neotropical mammal fauna, p. 335-358. In J.B.C. Jackson, A.F. Budd & A.G. Coates (eds.). Evolution and environment in tropical America. University of Chicago, Illinois, USA. [ Links ]
Weyl, R. 1957. Contribución a la geología de la Cordillera de Talamanca de Costa Rica. Instituto Geográfico de Costa Rica, San José, Costa Rica. [ Links ]
Wiens, J.A. 1989. The ecology of bird communities. Foundations and patterns. Cambridge University, Cambridge, USA. [ Links ]
Winker, K. & C.L. Pruett. 2006. Seasonal migration and morphological convergence in the genus Catharus (Turdidae). Auk 123: 1052-1068. [ Links ]
Wolf, L. 1976. Avifauna of the Cerro de la Muerte region Costa Rica. Am. Mus. Nov. No 2606. [ Links ]


{kind=link}