










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 suppl.1 San José Nov. 2009
Diversidad y distribución de palmas (Arecaceae) en tres fragmentos de bosque muy húmedo en Costa Rica
Virginia Aguilar Barquero & Fabiola Jiménez Hernández
Escuela de Biología, Universidad de Costa Rica, 11501-2060, San José, Costa Rica; vaguilarb@gmail.com; amenis86@gmail.com
Abstract: Palm (Arecaceae) diversity and distribution in three humid forest fragments in Costa Rica. To determine the existing differences between species composition of palms in forest fragments with different disturbance regimes, three forest fragments at "El Zota", Cariari de Guápiles, Costa Rica, were chosen according to their disturbance level: 1) Disturbed primary forest (B1-i), 2) Secondary forest (B2) and 3) Primary forest (B1). A field survey was carried out in nested plots, where palms were identified and counted according to three size categories: i) 0.15 m to 1.35 m ii) 1.35 m to 5 m and iii) more than 5 m in height. The diversity found in B1 and B1-i is very similar, meanwhile the B2 has the lowest diversity. Palm abundance and distribution were influenced by the level of disturbance of the fragment. The B1-i presents the higher diversity in the first size categories, while B1 has the highest abundance of palms over 5 m tall. Rev. Biol. Trop. 57 (Suppl. 1): 83-92. Epub 2009 November 30.
Key words: palms, forest fragments, humid forest, forest restoration, disturbance, palm diversity, Costa Rica.
La principal causa de la destrucción de grandes áreas boscosas en los trópicos es la transformación de estas zonas en terrenos de uso agrícola y ganadero (Clark et al. 1995, Scariot 1999, Wagner 2000). Esta situación provoca una paulatina pérdida de la capacidad productiva del ecosistema y un mayor empobrecimiento del sector campesino. Además, se presentan otros problemas indirectos, como la destrucción de los recursos hídricos y el daño en los ecosistemas costeros (Di Stefano 2004).
La fragmentación de hábitats puede provocar disminución de la complejidad, biodiversidad, capacidad de carga y productividad biológica de los ecosistemas, la pérdida de microhábitats, aislamiento de hábitats y cambios asociados a patrones de dispersión y migración (Scariot 1999, Montoya 2004). De igual manera, debido a la elevada mortalidad de árboles y daños que ocurren en los bordes, los fragmentos de bosque por lo general tienen claros y se encuentran en fases regenerativas. Estos efectos varían de acuerdo al tamaño, la forma y el grado de aislamiento del fragmento (Laurance et al. 1998).
Aunque se sabe que la fragmentación de las selvas tropicales afecta severamente a los árboles de gran porte, su efecto sobre otros estadios y formas de vida de las plantas es poco conocido (Benítez & Martínez 2003). Asimismo, existen pocos estudios que traten el efecto de la fragmentación del hábitat sobre la comunidad de palmas de bosques lluviosos (Laurance et al. 1998, Capers et al. 2005).
La familia Arecaceae se caracteriza por su diversidad y abundancia, con alrededor de 1 500 especies y 200 géneros en el mundo (Henderson et al. 1995). La gran abundancia y densidad de palmas en los bosques tropicales húmedos se encuentran en parte asociadas con los climas cálidos y húmedos que caracterizan estos ecosistemas (Ávalos et al. 2005, Bjorholm et al. 2005). Las palmas son una de las familias de plantas con ciclos de vida de mayor duración (Scariot 1999). Una de las etapas más críticas es la fase de establecimiento, que de acuerdo con Tomlinson (citado por Henderson 2002) consiste en un período largo durante los primeros estadios de desarrollo, en el cual el tallo alcanza el máximo diámetro. Después de esta etapa, la planta empieza a alargar más el tallo y alcanza potencial reproductivo.
Además, este grupo de plantas cumplen un papel ecológico importante, por encontrarse en todos los estratos del bosque y ser importantes para el mantenimiento de la estructura y la composición de éste (Scariot 1999). Asimismo, se les considera como posibles indicadores del nivel de alteración en bosques tropicales, debido a que la apertura de claros favorece la colonización de palmas (Svenning 1999a), cumpliendo un papel crucial en la regeneración y la dinámica general de los bosques (Fleischmann et al. 2005).
Este proyecto se realizó para cuantificar las diferencias existentes entre las comunidades de palmas en fragmentos de bosques con diferentes grados de alteración. Lo anterior con el objetivo de utilizar medidas de composición, abundancia y establecimiento de las especies de palmas como indicadores de la alteración en los fragmentos.
Materiales métodos
Área de estudio: El estudio se llevó a cabo en la finca "El Progreso" localizada al noreste de Costa Rica en la comunidad de El Zota (10o33.437 N, 83o44.177 W), a 45 m de elevación en la zona de Pococí, Limón. En esta estación se pueden encontrar distintos tipos de hábitats; entre ellos el bosque tropical lluvioso de bajura, pastizales, pantanos y áreas regeneradas de bosque secundario.
Se escogieron tres fragmentos de bosque de la finca y se clasificaron según su grado de alteración: 1) Bosque primario alterado (B1-i) de 5 ha, con 27 años de regeneración, limitado al NE, E y o por potreros y en el extremo So se encuentra conectado con el bosque primario. Además, toda el área del fragmento es atravesada por una quebrada la que tiene acceso el ganado, y existen senderos que se utilizan para agroecoturismo, por lo que el área se encuentra expuesta a estas alteraciones; 2) Bosque secundario (B2) de aproximadamente 3 ha, que fue utilizado como maizal y se abandonó hace alrededor de 15 años. Desde entonces se encuentra en regeneración y se han reforestado algunos sitios. Este bosque está delimitado por bananales y potreros y es recorrido por un sendero; 3) Bosque primario (B1) de aproximadamente 15 ha, del que se extrajo madera hace unos 25 años pero no se ha vuelto a practicar esta actividad. El B1 está rodeado por potreros y cuenta con algunos senderos para agroecoturismo. Sin embargo, en este sitio se encuentran las nacientes de agua de la finca, por esta razón casi no ha habido intervención antropogénica en los últimos 25 años (N. Chaves, com. pers). Por lo que se considera que este fragmento ha estado por un período de tiempo considerable en regeneración y por eso ha podido recuperar algunas características de bosque primario (palmas mayores de 5 m de altura y baja presencia de bejucos y maleza) en comparación con los demás fragmentos. Por ello se utilizó como un control, a pesar de que el fragmento no es estrictamente un bosque primario.
Determinación de diversidad y distribución de palmas: Se llevó a cabo un muestreo anidado en parcelas de 10x10 m, donde se contabilizaron e identificaron todas aquellas pal-mas mayores de 5 m de altura. Dentro de estas mismas parcelas se ubicó aleatoriamente una cuadrícula de 1 m2 para contabilizar e identificar las plántulas y juveniles mayores a 0.15 m pero menores de 1.35 m de altura. Además, en el centro de cada una de las parcelas de 10x10 m se hicieron cuatro parcelas de 5x2.5 m para las palmas mayores de 1.35 m pero menores de 5 m de altura. De esta manera en cada fragmento se realizaron un total de 78 parcelas: 13 de 10x10 m, 13 de 1 m2 y 52 de 5x2.5 m.
Sólo se consideraron las especies de palmas nativas de la finca. En todos los casos se excluyeron los puntos donde había pantanos, riachuelos, senderos, bordes o claros de bosque. Además, las parcelas se separaron 30 m entre sí y para reducir el efecto de borde se ubicaron a 10 m de los senderos y de los bordes de bosque.
Caracterización de fragmento: Se realizó una medición cualitativa de la cobertura vegetal del dosel por parcela anotando si ésta se encontraba completamente en sombra, parcialmente o si estaba expuesta al sol. En cada parcela se anotaron observaciones sobre el entorno (p. ej.: si la parcela se encontraba a poca distancia de un pantano) que servían para caracterizarlas. Además de esto, se midió la pendiente de cada parcela utilizando un clinómetro (Konustar, Konus optical & Sport Systems).
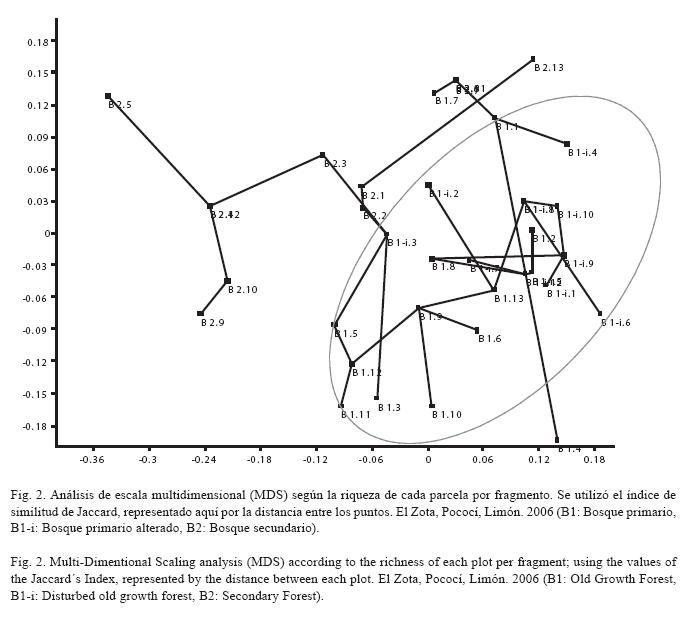
Análisis de datos: Para cuantificar la diversidad de cada uno de los fragmentos de bosque se calcularon índices de Shannon (H) y de equidad (J). Además, para evaluar la similitud entre los fragmentos de bosque y las parcelas según la riqueza de palmas se elaboró, con el programa estadístico PAST versión 1.83, un análisis de escala multidimensional (MDS) usando el coeficiente de similitud de Jaccard (Jaccard 1908), que ignora la ausencia compartida de especies para cada fragmento.
Se estimó el establecimiento de las distintas especies en cada uno de los fragmentos de bosque según la abundancia de palmas en cada una de las categorías de tamaño (entendiéndose por establecimiento al alcance del potencial reproductivo según el tamaño y la especie). Para esto se calcularon coeficientes de porcentaje de similitud Renkonens PS (Scariot 1999) para cada fragmento según la categoría de tamaño de las palmas.
Resultados
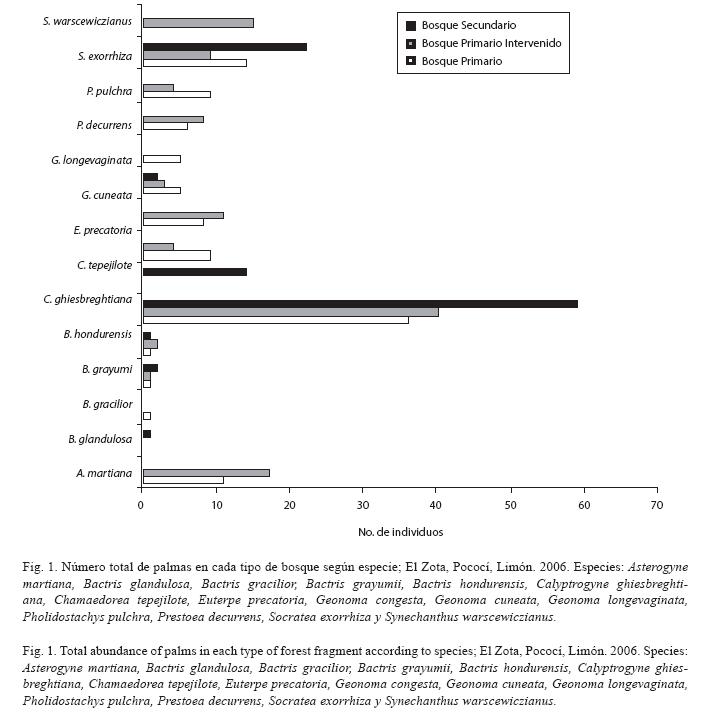
En total se observaron 392 palmas; de éstas, 147 se encontraron en el bosque primario, 127 en el bosque primario alterado y 118 en el bosque secundario. Durante el muestreo se hallaron 15 especies de palmas, siendo Socratea exorrhiza y Calyptrogyne ghiesbreghtiana las más comunes.
El bosque primario posee una diversidad de 84% (HB1= 3.10), que difiere muy poco del 82% de diversidad calculado para el bosque primario alterado (HB1-i= 2.82). El bosque secundario registra una diversidad de 41% (HB2= 1.16), que es la más baja de los tres fragmentos estudiados (Fig. 1). Según el MDS obtenido, se observa que la mayoría de las parcelas del B1 y el B1-i son muy similares ya que éstas se agrupan; mientras que muchas de las parcelas del B2 se alejan de esta conglomeración (Fig. 2). Esto evidencia que existe una mayor similitud entre los fragmentos B1 y B1-i, en cuanto que el B2 es el que más difiere de éstos.
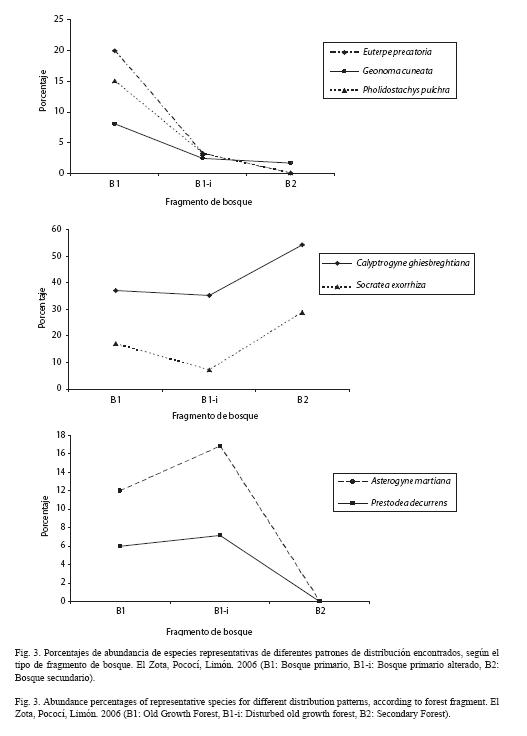
Se observaron tres patrones de distribución de las especies encontradas en la finca. Estos patrones varían según el grado de alteración de los fragmentos: 1) algunas especies disminuyen la abundancia conforme aumenta el grado de alteración del sitio; 2) algunas especies poseen mayor abundancia en B2 y B1, pero menor en el bosque primario alterado; 3) mientras que otras especies exhiben un patrón en el que la mayor abundancia se da en el B1-i (Fig. 3).
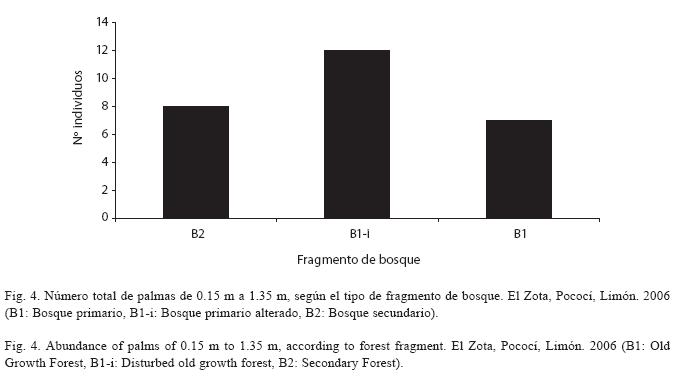
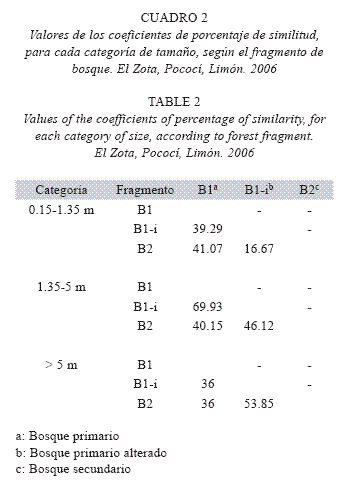
En cuanto a la diversidad de palmas de 0.15 m a 1.35 m, el B1-i presenta una mayor riqueza y abundancia, mientras que el B2 y el B1 son similares entre sí (Cuadro 1, Fig. 4). Según el resultado del coeficiente de porcentaje de similitud para las palmas con una altura de 0.15 a 1.35 m, la mayor diferencia se registra entre el bosque secundario y el bosque primario intervenido. La diversidad de palmas de 1.35 a 5 m, según el coeficiente de porcentaje de similitud, es muy similar en el B1 y el B1-i. Finalmente, se observó una mayor abundancia de palmas mayores a 5 m en el bosque primario. Para esta última categoría de tamaño, los coeficientes de porcentaje de similitud demuestran que el B1 es el fragmento que difiere más de los tres estudiados, mientras que los fragmentos B2 y B1-i son más similares entre sí (Cuadro 2).

Discusión
La abundancia de palmas observada en este estudio se debe, principalmente, a la alta precipitación anual de la zona atlántica de Costa Rica (entre 2 500 y 6 000 mm por año), que favorece la distribución y el establecimiento exitoso de estas plantas (Henderson et al. 1995). Las diferencias entre los fragmentos de bosque en cuanto a riqueza y abundancia de palmas se deben, en gran medida, a los diferentes grados de alteración de cada uno de los sitios. De acuerdo con Chazdon (1996), los patrones de abundancia y riqueza de un fragmento varían según las actividades humanas que anteriormente se practicaron en el sitio. Además, la composición, abundancia y diversidad de especies de plantas en un bosque varían de acuerdo con la etapa de sucesión. Se ha demostrado que conforme aumenta la edad de regeneración, más heterogéneo será el bosque (Solís-Corrales 2000). Por este motivo, el fragmento de bosque primario y el primario alterado presentan una mayor diversidad de palmas, mientras que el secundario es el más homogéneo. Además, las actividades propias de la plantación de maíz pudieron haber afectado en gran medida la regeneración del bosque secundario, ya que impedían la germinación de semillas y el crecimiento de plántulas. Lo anterior pudo generar un cambio en los patrones de distribución espacial de las plantas, que eventualmente ocasionó una disminución en la diversidad de palmas de la zona. Esto se puede evidenciar en la alta homogeneidad del B2, donde C. ghiesbreghtiana tiende a ser dominante.
Los remanentes de bosques primarios son importantes para la regeneración en sitios aledaños (Endress & Chinea 2001). La existencia de estos bosques crea un efecto de masa, ya que constituyen una fuente de propágulos para los sitios adyacentes con condiciones menos favorables (Svenning 1999a). La conexión y el efecto de masa existente entre el bosque primario y el bosque primario intervenido ocasionan que éstos posean porcentajes de diversidad muy semejantes, mientras que el bosque secundario, al estar a una mayor distancia del primario, tiene menor influencia del mismo.
En cuanto al primer patrón de distribución, en el cual algunas de las especies disminuyen su abundancia conforme aumenta el grado de alteración del sitio, se observa que Euterpe precatoria, Geonoma cuneata y Pholidostachys pulchra podrían ser especies con una mayor sensibilidad a los sitios más alterados. Sin embargo, estas especies se diferencian en las adaptaciones que poseen para colonizar ambientes menos alterados. Por ejemplo, E. precatoria se encuentra principalmente en el bosque primario de esta finca debido a la topografía plana y pantanosa que predomina en el mismo; Henderson et al. (1995) señalan que este tipo de hábitat es uno de los más comunes de E. precatoria en zonas bajas. Por su parte, G. cuneata es una palma propia de sotobosque y tolera la sombra (Capers et al. 2005); por esta razón al ser el bosque secundario más abierto y permitir una mayor entrada de luz al sotobosque, esta especie es menos abundante en el sitio.
El patrón de distribución de S. exorrhiza y C. ghiesbreghtiana es contrario al de especies con hábitos similares, como Prestoea decurrens y Asterogyne martiana, respectivamente. La colonización exitosa de Calyptrogyne en el terreno de la antigua plantación de maíz se dio desde un parche de bosque conservado en los alrededores; así, esta especie fue pionera en el sitio. Además, al ser una palma de sotobosque, las semillas de Calyptrogyne tienen una mayor probabilidad de supervivencia que otras especies, aunque caigan cerca de la planta madre, debido a que ésta tendrá un menor efecto de sombra sobre las plántulas (Ruokolainen & Vormisto 2000). Por otra parte, la distribución espacial de Asterogyne podría ser influenciada por la presencia de C. ghiesbreghtiana, ya que al ser especies de sotobosque, ambas compiten por recursos similares. A. martiana, al ser una especie tolerante a la sombra (Chazdon 1985), está mejor adaptada a fragmentos de regeneración avanzada porque tendrán una mayor cobertura de dosel y una menor penetración de luz en el sotobosque (Capers et al. 2005). No obstante, la mayor abundancia de esta palma se da en el bosque primario alterado; esto puede ser causa de un crecimiento más rápido de las plántulas en este sitio por la mayor disponibilidad de luz que se observó en comparación con el bosque primario. S. exorrhiza es más abundante en bosque secundario; una razón es que al desarrollar raíces fúlcreas en sus primeras etapas de desarrollo, el tallo se alarga más rápidamente, por lo que la fase crítica será más corta que la de otras palmas arbóreas. Asimismo, las raíces fúlcreas de esta palma facilitan la recuperación cuando el tallo ha sido dañado; por ej., por caída de ramas (Svenning 1999b, Ávalos et al. 2005). Lo anterior le confiere cierta ventaja sobre otras palmas con hábitos similares, como P. decurrens, y explica por qué Socratea se encuentra en todos los fragmentos estudiados.
La mayor abundancia de palmas de 0.15 m a 1.35 m en el bosque primario alterado, que comúnmente está expuesto a alteraciones tanto antropogénicas (mayoritariamente por extracción de semillas de palmas) como a la entrada de ganado y caída de árboles (N. Chaves, com. pers., 2006), concuerda con lo observado por Wright & Duber (2001), que encontraron una mayor supervivencia y reclutamiento de semillas en sitios con una exposición frecuente a alteraciones. En cambio, la menor abundancia de plántulas en B1 y B2 puede atribuirse a que ambos sitios esporádicamente se encuentran expuestos a alteraciones, ya que el bosque secundario se encuentra aislado y el bosque primario se encuentra en una etapa avanzada de regeneración. Sin embargo, la semejanza entre el B1 y el B1-i en cuanto a la diversidad de plántulas puede ser producto del efecto de masa (dada la cercanía del bosque primario) y a la conexión geográfica entre ambos fragmentos de bosque.
La similitud en diversidad de palmas de 1.35 a 5 m, entre el bosque primario y el bosque primario alterado, puede ser considerada como un indicador de condiciones de regeneración favorables del B1-i. Al respecto, Henderson (2002) señala que las palmas son plantas que comúnmente poseen una etapa crítica longeva, la cual corresponde a la fase de establecimiento. En el momento en que las palmas superan esta etapa, comienzan a crecer longitudinalmente. Así, las palmas con una altura de 1.35 m a 5 m han superado la fase crítica del ciclo de vida. Por tanto, una diversidad y abundancia considerables y similares de estas palmas en los bosques B1 y B1-i evidencian su establecimiento y una buena regeneración de estos bosques.
Los pocos años de regeneración del bosque secundario y las constantes alteraciones presentes en el bosque primario alterado, factores aunados a los largos ciclos de vida de las palmas (Scariot 1999), explican en gran medida la similitud entre estos dos fragmentos en diversidad de palmas de más de 5 m. Además, la mayor diversidad de palmas de esta categoría de tamaño en el B1 indica que, a pesar de que se extrajo madera en el pasado, el sitio no fue significativamente afectado por esa actividad, ya que aún mantiene un buen estado de madurez y establecimiento de palmas.
Las especies de palmas encontradas en el sitio de estudio se pueden utilizar como indicadores del grado de alteración de la zona, ya que éstas son propias de bosque y, por ello, su establecimiento en zonas deforestadas (p.e. pastizales y bosques secundarios en etapas iniciales de desarrollo) puede ser difícil. La diversidad de palmas observada en la finca indica que la regeneración se está dando exitosamente. Es importante enfatizar que este proceso de regeneración ha sido favorecido por la existencia de un fragmento de bosque primario de extensión considerable dentro de la finca, que actúa como un núcleo de dispersión de propágulos. El nivel de diversidad en el bosque secundario (41%) podría indicar que allí el proceso de regeneración es positivo. Además, se considera que si se mantienen las condiciones actuales, el ecosistema podrá llegar a recuperarse completamente.
Agradecimientos
A la familia Alpízar Chaves, especialmente a Nuria Chaves, por la hospitalidad y ayuda brindada en la identificación de palmas. A Carlos o. Morales, por el apoyo, la revisión y el interés en el trabajo. A José Francisco Di Stefano y Gerardo Ávalos, por el asesoramiento y el interés en el proyecto. A José Pablo Jiménez y Gerardo Umaña por la ayuda en el análisis estadístico.
Resumen
Para determinar las diferencias existentes entre la composición de especies de palmas en fragmentos de bosque con diversos grados de alteración, se escogieron tres fragmentos en "El Zota", Costa Rica, según el grado de alteración: 1) Bosque primario alterado (B1-i), 2) Bosque secundario (B2) y 3) Bosque primario (B1). Se hizo un muestreo con parcelas anidadas donde se identificaron y contabilizaron las palmas según tres categorías de tamaño: i) 0.15 m a 1.35 m, ii) 1.35 m a 5 m y iii) más de 5 m de alto. Se observó que la diversidad de B1 y B1-i es muy similar, mientras que B2 posee la menor diversidad. La abundancia y distribución de las palmas estaba influenciada por el nivel de alteración del fragmento. El B1-i presenta una mayor diversidad de la primera categoría de tamaño, mientras que B1 tiene mayor abundancia de palmas sobre 5 m de alto.
Palabras clave: Palmas, fragmentos de bosque, bosque húmedo, regeneración de bosques, alteración, diversidad de palmas, Costa Rica.
Recibido 09-III-2007. Corregido 06-X-2008. Aceptado 18-XI-2008.
Referencias
Ávalos, G., D. Salazar & A.L. Araya. 2005. Stilt Root Structure in the Neotropical Palms Iriartea deltoidea and Socratea exorrhiza. Biotropica 37: 44-53. [ Links ]
Benítez, J. & M. Martínez. 2003. impact of forest fragmentation on understory plant species richness in Amazonia. Conserv. Biol. 17: 389-400. [ Links ]
Bjorholm, S., J.C. Svenning, F. Skov & H. Balslev. 2005. Environmental and spatial controls of palm (Arecaceae) species richness across the Americas. Global Ecol. Biogeogr. 14: 423-429. [ Links ]
Capers, R.S., R.L. Chazdon, A. Redondo-Brenes & B. Vilchez-Alvarado. 2005. Succesional dynamics of woody seedling communities in wet tropical secondary forests. J. Ecol. 93: 1071-1084. [ Links ]
Chazdon, R.L. 1986. Leaf display, canopy structure and light interception of two understory palm species. Amer. J. Bot. 72: 1493-1502 [ Links ]
Chazdon, R.L. 1996. Spatial heterogeneity in tropical forest structure: canopy palms as landscape mosaics. Trends Ecol. Evol. (TREE) 11: 8-9. [ Links ]
Clark, D., D. Clark, R. Sandoval & M. Castro. 1995. Edaphic and human effects on landscape-scale distributions of the tropical rain forest palms. Ecology 76: 2581-2594. [ Links ]
Di Stefano, J.F. 2004. Establecimiento y crecimiento inicial de varias especies forestales en pastizales degradados en la cuenca del río Picagres, Puriscal, p. 33-50. In F. Montoya (ed.). investigación para la recuperación de áreas degradadas en la cuenca del río Picagres, Puriscal (1992-2004). SiRECo, San José, Costa Rica. [ Links ]
Fleischmann, K., P. Edwards, D. Ramseie & J. Kollamann. 2005. Stand structure, species diversity and regeneration of endemic palm forest on the Seychelles. Afr. J. Ecol. 43: 291-301. [ Links ]
Henderson, A., G. Galeano & R. Bernal. 1995. Field guide to the palms of the Americas. Princeton Univ., Princeton, EEUU. [ Links ]
Henderson, A. 2002. Evolution and Ecology of Palms. The New york Bot. Gard., Nueva york, EEUU. [ Links ]
Krebs, C.J. & A.J. Kenney. 2000. Programs for Ecological Methodology. Versión 5.2. Vancouver, Canada: DELPHi 4 [ Links ]
Jaccard, P. 1908. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaudoise Scie. Natur. 44: 223–270. [ Links ]
Laurance, W., L. Ferreira, J. De Merona & S. Laurance. 1998. Rain Forest fragmentation and the dynamics of Amazonian tree communities. Ecology 79: 20322040. [ Links ]
Montoya, F. 2004. Degradación y rehabilitación de ecosistemas terrestres: Estado de la cuestión, p. 10-15. In F. Montoya (ed.). investigación para la recuperación de áreas degradadas en la cuenca del río Picagres, Puriscal (1992-2004). SiRECo, San José, Costa Rica. [ Links ]
Rohlf, F.J. 1997. NTSyS-pc: numerical taxonomy and multivariate analysis system. Versión 2.0. Setauket, New york: Exeter Software, EEUU. [ Links ]
Ruokolainen, K. & J. Vormisto. 2000. The most widespread Amazonian palms tend to be tall and habitat generalists. Basic Appl. Ecol. 1: 97-108. [ Links ]
Scariot, A. 1999. Forest fragmentation effects on palm diversity in central Amazonia. J. Ecol. 87: 66-76. [ Links ]
Solís-Corrales, M. 2000. Análisis de crecimiento en tres bosques secundarios de diferentes edades en la zona de Boca Tapada, San Carlos, Costa Rica, durante el período 1995-1998. CoSEFoRMA, San José, Costa Rica. [ Links ]
Svenning, J. 1999a. Microhabitat specialization in a species-rich palm community in Amazonian Ecuador. J. Ecol. 87: 55-65. [ Links ]
Svenning, J. 1999b. Recruitment of tall arborescent palms in yasuní National Park, Amazonian Ecuador: Are large treefall gaps important? J. Trop. Ecol. 15: 355-366. [ Links ]
Wagner, U. 2000. Efectos de la corta selectiva sobre la composición florística y la estructura de los bosques húmedos de la vertiente atlántica de Costa Rica. Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ), Alemania. [ Links ]
Wrigth, S.J. & H.C. Duber. 2001. Poachers and Forest Fragmentation Alter Seed Dispersal, Seed Survival, and Seedling Recruitment in the Palm Attalea butyracea with implications for Tropical Tree Diversity. Biotropica 33: 583-595. [ Links ]

