











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.57 no.1-2 San José mar./jun. 2009
Gilberto Avalos1, Miryam P. Damborsky1, María E. Bar1, Elena B. Oscherov1 & E. Porcel2
1. Cátedra Biología de los artrópodos, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, Av. Libertad 5470, W 3404 AAS, Corrientes, Argentina; gilbertoa@exa.unne.edu.ar, mdambor@exa.unne.edu.ar, mebar@exa.unne.edu.ar, ebosche@exa.unne.edu.ar
2. Cátedra de estadística descriptiva, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, Av. Libertad 5470, W 3404 AAS, Corrientes, Argentina; eporcel@arnet.com.ar
Abstract: Composition of the Araneae (Arachnida) fauna of the provincial Iberá Reserve, Corrientes, Argentina. A survey of the spider community composition and diversity was carried out in grasslands and woods in three localities: Colonia Pellegrini, Paraje Galarza and Estancia Rincón (iberá province Reserve). Pit fall traps, leaf litter sifting, foliage beating, hand collecting and sweep nets were used. Shannons diversity index, evenness, Berger-Parkers dominance index, ß and 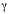 diversity were calculated, and a checklist of spider fauna was compiled. Species richness was estimated by Chao 1, Chao 2, first and second order Jack-knife. A total of 4 138 spiders grouped into 150 species from 33 families of Araneomorphae and two species from two families of Mygalomorphae were collected. Five species are new records for Argentina and eleven for Corrientes province. Araneidae was the most abundant family (39.8%), followed by Salticidae (10.9%), Anyphaenidae (7.9%), Tetragnathidae (7.4%), and Lycosidae (5.5%). The other families represented less than 5% of the total catch. The web-builder guild had the highest number of specimens and the highest richness index. The abundance, observed richness, Shannon diversity and evenness indexes were highest in Colonia Pellegrini woodland and Paraje Galarza grassland. Alpha diversity represented 89% of the gamma; the remaining 11% corresponded to ß diversity. According to the indexes, between 67% and 97% of the existing spider fauna was represented in the collected specimens from iberá. Rev. Biol. Trop. 57 (1-2): 339-351. Epub 2009 June 30.
diversity were calculated, and a checklist of spider fauna was compiled. Species richness was estimated by Chao 1, Chao 2, first and second order Jack-knife. A total of 4 138 spiders grouped into 150 species from 33 families of Araneomorphae and two species from two families of Mygalomorphae were collected. Five species are new records for Argentina and eleven for Corrientes province. Araneidae was the most abundant family (39.8%), followed by Salticidae (10.9%), Anyphaenidae (7.9%), Tetragnathidae (7.4%), and Lycosidae (5.5%). The other families represented less than 5% of the total catch. The web-builder guild had the highest number of specimens and the highest richness index. The abundance, observed richness, Shannon diversity and evenness indexes were highest in Colonia Pellegrini woodland and Paraje Galarza grassland. Alpha diversity represented 89% of the gamma; the remaining 11% corresponded to ß diversity. According to the indexes, between 67% and 97% of the existing spider fauna was represented in the collected specimens from iberá. Rev. Biol. Trop. 57 (1-2): 339-351. Epub 2009 June 30.
Key words: Araneae, woods, grassland, diversity, Iberá Reserve, Corrientes.
Las arañas comprenden uno de los grupos más numerosos del reino animal, con más de 40 000 especies descritas (Platnick 2008). Tienen una amplia aceptación en los estudios ecológicos como indicadores de calidad ambiental (Clausen 1986, Maelfait et al. 1990). Como depredadores cumplen una destacada función en la regulación de las poblaciones de artrópodos, influyen en la densidad de la fauna de detritívoros y afectan los procesos de descomposición (Wise 1993, 2002).
En contraste con lo que ocurre en áreas templadas, los estudios sobre diversidad de los ecosistemas tropicales son más bien recientes (Russell-Smith 1999), aunque en los bosques tropicales se espera que se concentre alrededor del 80% de la araneofauna aún no conocida (Coddington y Levi 1991).
En las últimas décadas se destacan las investigaciones ecológicas en los bosques tropicales de América Latina, como las efectuadas en la Amazonia central y en la Amazonia Peruana y en diferentes tipos de bosques de Colombia (Höfer et al. 1990, Silva y Coddington 1996, Flórez 2000).
En Argentina el conocimiento taxonómico de algunas familias de arañas es amplio. Sin embargo, las investigaciones referidas a aspectos ecológicos de comunidades de arañas asociadas a áreas naturales son escasas. Al respecto se puede mencionar los estudios efectuados en un área natural protegida de la provincia de Formosa (Corronca y Abdala 1994), en bosques degradados del Chaco Húmedo (Avalos et al. 2007), y en el Parque nacional Mburucuyá, provincia de Corrientes (Rubio et al. 2008).
El Sistema del Iberá, situado en el centro – norte de la provincia de Corrientes, abarca una superficie aproximada de 12 300 km2 y constituye un área de contacto entre grandes provincias fitogeográficas (Carnevali 1994). Es uno de los sistemas de humedales más extensos e importantes de América neotropical, fue declarado Reserva provincial en 1983, y Sitio Ramsar en enero del 2002. Debido a su extensión, ubicación geográfica y a la escasa intervención humana, la reserva se mantiene relativamente poco alterada.
Las arañas, por su abundancia, biomasa y diversidad están directamente correlacionadas con la diversidad ambiental (Samu y Lövei 1995). Se estima que la reserva alberga diferentes comunidades de arañas que caracterizan sus numerosos biotopos.
La presente investigación tiene como objetivo conocer la riqueza y la estructura de la comunidad de Araneae y describir la diversidad en dos unidades ambientales (bosque y pastizal) de la Reserva provincial Iberá.
Materiales y métodos
Área de estudio: El iberá se caracteriza por presentar un clima subtropical húmedo, con baja frecuencia de heladas anuales y volumen pluvial de 1 700-1 800 mm por año (Neiff y Poi de Neiff 2006).
Se seleccionaron dos unidades ambientales terrestres: bosque higrófilo y pastizal. El bosque higrófilo se presenta como "isleta de selva". La vegetación del estrato herbáceo se caracteriza por la abundancia de Oplismenus hirtellus y Chaptalia nutans. En el estrato arbustivo, de densidad variable, se encuentran Brunfelsia australis, Byttneria urticifolia y Ephedra tweediana. El estrato arbóreo de mediano y bajo porte (8-15 m), está representado principalmente por Tabebuia heptaphylla, Peltophorum dubium, Patagonula americana, Enterolobium contortisiliquum, Nectandra angustifolia, Guadua chocoensis y en menor frecuencia Fagara hyemalis y Pithecellobium scalare. En el pastizal es dominante Andropogon lateralis, que conforma el estrato más sobresaliente; forman parte de un estrato más bajo Paspalum notatum, Cuphea sp., Tibouchina gracilis, Angelonia integerrima y Desmodium canum, entre otras (Tressens et al. 2002).
El estudio se llevó a cabo en Colonia Pellegrini (28°3241" S, 57°1157" W) departamento San Martín, Estancia Rincón (28º2446" S, 57º2114" W) departamento Mercedes, y Paraje Galarza (28°0401" S, 56°4202" W) departamento Santo Tomé. Las dos primeras localidades están incluidas en la provincia fitogeográfica del Espinal, distrito del Ñandubay, y el Paraje Galarza está situado en la provincia Paranaense, distrito de los Campos Correntinos Misioneros (Carnevali 1994).
Trabajo de campo: Las recolecciones se realizaron entre el 24 y 31 de agosto, y el 15 y 22 de diciembre del 2005, y entre el 3 y 10 de abril de 2006 en cada localidad. En las unidades ambientales, de cada una de las tres localidades de estudio, se trazó un transecto de 200 m de longitud por 2 m de ancho y se seleccionaron cinco sitios de muestreo separados entre sí por 40 m. Para las diferentes técnicas utilizadas en cada sitio, se recolectaron tres muestras.
Tanto en el bosque como en el pastizal se efectuaron capturas manuales nocturnas, llevadas a cabo por dos personas durante una hora; los ejemplares se recolectaron con pinzas y la observación se realizó desde 50 cm arriba del nivel del suelo hasta 2 m de altura. Asimismo se instalaron 3 trampas de caída ("pit-fall") en cada sitio, separadas por dos metros y formando un triángulo en el transecto, se utilizaron recipientes plásticos de 500 ml de capacidad y 11 cm de diámetro con solución conservante (etilenglicol:agua, en proporción 1:10, con gotas de detergente para disminuir la tensión superficial). Las trampas se enterraron a nivel del suelo sin cubierta y permanecieron activas durante 48 horas en cada campaña.
En el bosque se realizó golpeteo de follaje, que consistió en 15 golpes con una vara sobre la vegetación arbustiva y en la sección baja del estrato arbóreo, el material se recolectó sobre un lienzo blanco de 2.50 m2. Se tamizó también una superficie de 0.50 m2 de hojarasca, con un tamiz de 10 x 15mm de apertura de malla. En el pastizal se efectuaron 100 golpes con red de segado al azar a lo largo de los transectos. Las arañas recolectadas se colocaron en bolsas de polipropileno rotuladas según la unidad ambiental, localidad y técnica mediante la cual se capturaron, y se fijaron en alcohol etílico al 70%.
Trabajo de laboratorio: Los ejemplares se identificaron a nivel de familia, morfoespecie y especie con claves específicas, se separaron según sexo, y se agruparon en gremios según la conducta adoptada por el depredador en relación con la presa. (Enders 1976, Dippenaar- Schoeman et al. 1989, Uetz 1991, Liljestrhröm et al. 2002).
Análisis de datos: La abundancia se describió según modelo de distribución geométrica, logarítmico y log-normal, y se probó el modelo de distribución mediante el test c² de bondad de ajuste.
Se calcularon los índices de diversidad de Shannon-Wiener, dominancia de Berger-Parker, equitatividad, y la riqueza específica para cada unidad ambiental y localidad, con el programa PAST (Hammer et al. 2006). La significación estadística de estos índices se probó mediante pruebas de permutaciones (Good 2000) a un nivel de significación a= 0.05.
La similitud entre localidades y ambientes se midió según el índice de Jaccard (IJ). La partición de la diversidad gamma () en sus componentes alfa y beta (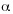 y ß) se analizó a nivel del total del área muestreada y para cada ambiente, aplicando la fórmula propuesta por Lande (1996) basada en el índice de Shannon (Moreno 2001).
y ß) se analizó a nivel del total del área muestreada y para cada ambiente, aplicando la fórmula propuesta por Lande (1996) basada en el índice de Shannon (Moreno 2001).
La estimación de riqueza de especies se efectuó mediante los índices no paramétricos Chao 1, Chao 2, Jackknife de primero y de segundo orden con el programa EstimateS 7.5 (Colwell 2005). Para el cálculo de los índices solamente se ingresaron las arañas que pudieron identificarse a nivel taxonómico de especie o morfoespecie (n= 3808).
Los individuos recolectados se depositaron en la colección de la Cátedra de Biología de los Artrópodos de la Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste (CARTROUNNE).
Resultados
El total de arañas recolectadas fue 4 138 de las cuales el 20.5% resultaron hembras (n= 847) y el 10.9% machos (n= 453), el resto de los ejemplares fueron juveniles. En Colonia Pellegrini se recolectó el 43.3% de los ejemplares, en Paraje Galarza el 36.2%, y en Estancia Rincón el 20.4%. (Cuadro 1).

Se identificaron 33 familias y 150 especies del infraorden Araneomorphae, y dos familias y dos especies del infraorden Mygalomorphae, totalizando 152 especies/ morfoespecies (Cuadro 2). Se registraron 19.1% (n=29) de especies/morfoespecies con un solo individuo ("singletons"), y 7.2% (n=11) con dos individuos ("doubletons").
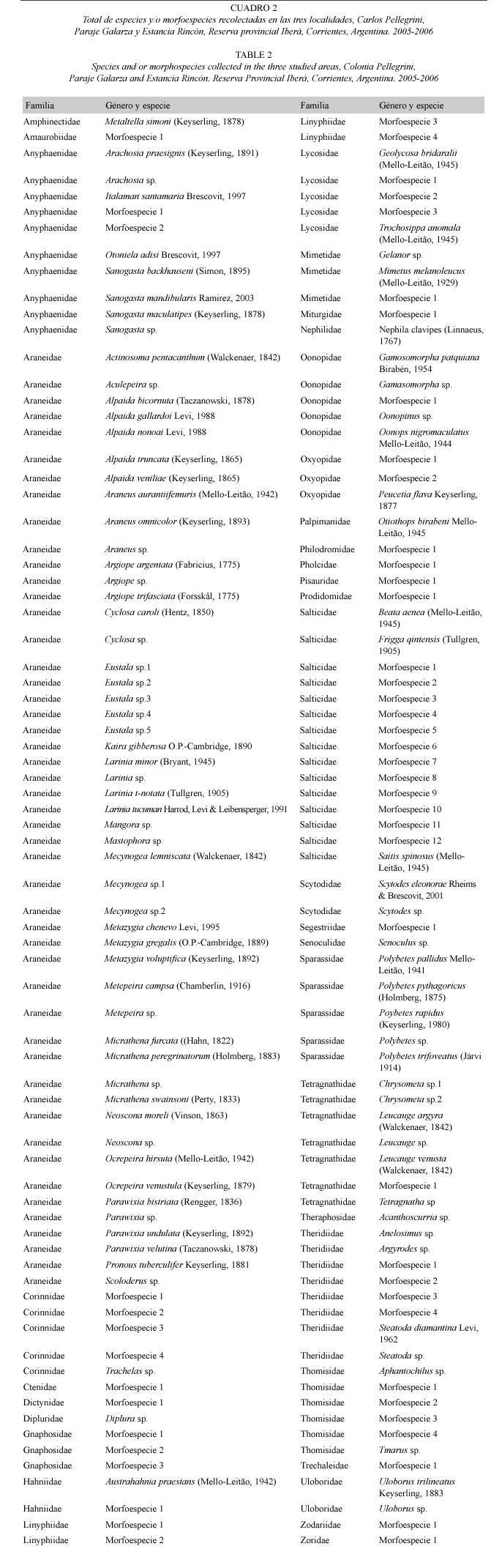
Se destaca que Otoniela adisi Brescovit, 1997, Kaira gibberosa O.P.-Cambridge, 1890, Mimetus melanoleucus (Mello-Leitão, 1929), Leucauge argyra (Walckenaer, 1842), Leucauge venusta (Walckenaer, 1842), son nuevos registros para Argentina. Metaltella simoni (Keyserling, 1877), Italaman santamaria Brescovit, 1997, Sanogasta backauseni (Simon, 1895), Sanogasta maculatipes (Keyserling, 1878), Alpaida gallardoi Levi, 1988, Araneus omnicolor (Keyserling, 1893), Larinia t-notata (Tullgren, 1905), Mecynogea lemniscata (Walckenaer, 1842), Micrathena peregrinatorum (Holmberg, 1883), Ocrepeira venustula (Keyserling, 1879) y Geolycosa bridaralii (Mello-Leitão, 1945) son nuevos registros para la provincia de Corrientes.
Las familias encontradas únicamente en Colonia Pellegrini fueron Amphinectidae, Dipluridae, Palpimanidae, Segestridae, Uloboridae, Nephilidae y Zodariidae; en tanto Prodidomidae fue exclusiva de Estancia Rincón, y Senoculidae de Paraje Galarza.
Ctenidae y Pisauridae se hallaron en Estancia Rincón y en Galarza; Pholcidae y Scytodidae se detectaron en Colonia Pellegrini y en Estancia Rincón, Miturgidae y Amaurobiidae en Paraje Galarza y Colonia Pellegrini, las restantes familias se encontraron en las tres localidades estudiadas.
Araneidae resultó la familia más abundante y predominó en las tres localidades (n=1647), seguida por Salticidae (n=452), Anyphaenidae (n=329), Tetragnathidae (n=306), Lycosidae (n=229) y Theridiidae (n=217).
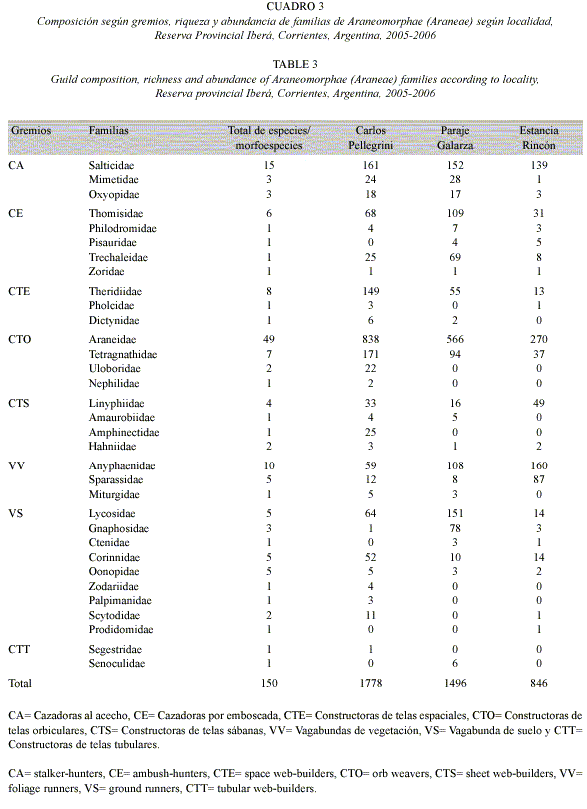
Se reconocieron 8 grupos funcionales: cazadoras al acecho, cazadoras por emboscada, constructoras de telas espaciales, constructoras de telas orbiculares, constructoras de telas sábanas, vagabundas de vegetación, vagabundas de suelo y constructoras de telas tubulares.
Las arañas tejedoras de telas orbiculares fueron las de mayor abundancia y riqueza de especies (S= 59) (Cuadro 3). En Colonia Pellegrini el número de individuos de este gremio (n= 1033) sumado al de cazadoras al acecho (n= 203) conformaron el 69% de la abundancia relativa. En Paraje Galarza las tejedoras de telas orbiculares (n= 660) y las vagabundas de suelo (n= 245) constituyeron el 60%, mientras que en Estancia Rincón el mismo gremio, junto con las vagabundas de vegetación (n= 307 y n= 247) representaron el 65% del
total de individuos. Las constructoras de telas espaciales, de telas sábana, y telas tubulares fueron los gremios de menor abundancia en las tres localidades, con un porcentaje menor al 10%.
En las tres localidades y ambientes muestreados, la distribución de la abundancia se ajustó al modelo logarítmico (p= 0.05) (Fig. 1).
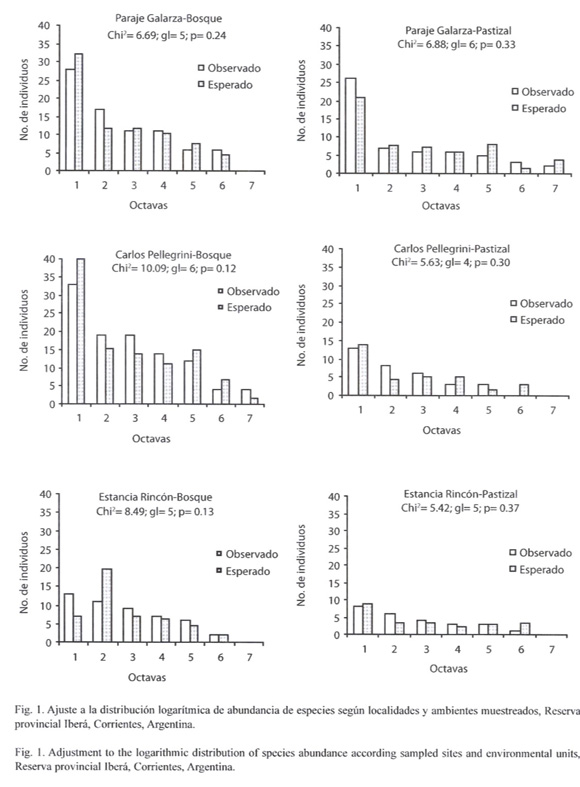
Colonia Pellegrini presentó la mayor riqueza específica, abundancia y diversidad (Cuadro 4), la dominancia fue superior en Estancia Rincón (p< 0.05), y no hubo diferencia en los valores de equitatividad (p> 0.05). Al comparar los parámetros entre las unidades ambientales y entre localidades, resultó que la riqueza de especies, abundancia, diversidad y dominancia fueron superiores en los bosques muestreados en Colonia Pellegrini (p< 0.05). En la unidad muestreal del pastizal, no hubo diferencia en la equitatividad (p> 0.05), la riqueza, abundancia y diversidad fueron mayores en Galarza, y la dominancia en Estancia Rincón (p< 0.05). En el pastizal de Estancia Rincón, Alpaida veniliae (Keyserling, 1865), Alpaida nonoai Levi, 1988 y dos morfoespecies de la familia Anyphaenidae, representaron el 63% de la abundancia total.
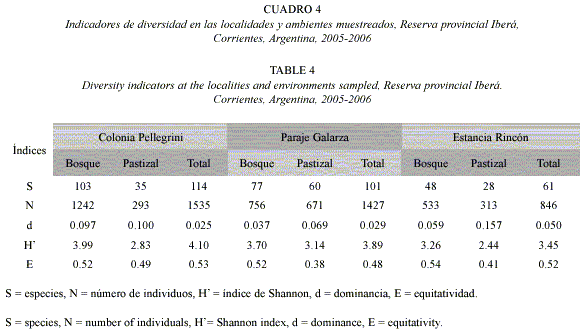
La mayor similitud entre unidades ambientales y localidades se registró entre bosques de Colonia Pellegrini y de Paraje Galarza (IJ= 0.46). La diversidad a fue alta y representó el 89% de la diversidad registrada en las localidades muestreadas, la diversidad ß, resultante del recambio de especies entre localidades, constituyó el 11% restante. valores similares de estos índices de diversidad se obtuvieron tanto para bosque como para pastizal (Cuadro 5).
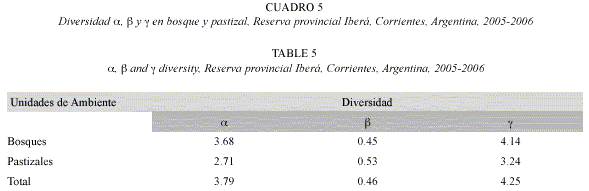
Al comparar la riqueza observada y estimada en las unidades ambientales y localidades según los diferentes estimadores, se comprobó que la recolección osciló entre el 67% y el 97% de las especies que se consideran están presentes en la Reserva (Cuadro 6). El estimador Jackknife 2 predijo las estimaciones más altas para las distintas localidades. La menor diferencia de valores estimados se registró en Colonia Pellegrini, localidad en que la riqueza hallada varió entre el 77 y 86% de la riqueza calculada. En Paraje Galarza, en cambio, la divergencia entre los valores observados y los estimados fue amplia, colectándose entre el 67 y 90% de especies presentes. Faltaría aún localizar de 5 a 28 especies en bosque, y de 24 a 41 especies en pastizal. Según el estimador Chao 2 o Jackknife 2, en Estancia Rincón sólo quedan por encontrar de 2 a 12 especies.
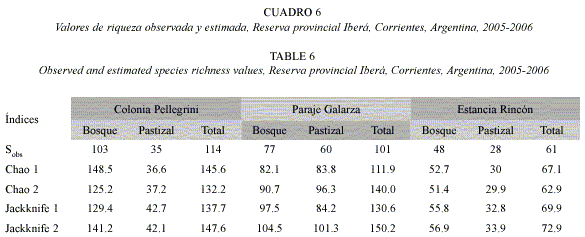
Discusión
Las familias de arañas halladas en el área de estudio representan el 79.5% (n= 35) del total de familias registradas en 19 departamentos de la provincia de Corrientes (Avalos et al. 2006), y el 56.5% identificadas hasta el presente para Argentina (Platnick 2008).
El mayor número de ejemplares juveniles, de adultos hembras, y el abundante hallazgo de arañas de las familias Araneidae, Salticidae y Anyphaenidae, es coincidente con las observaciones en dos bosques húmedos en el departamento Capital de Corrientes (Avalos et al. 2007). La abundancia de ejemplares de estas familias, junto a Lycosidae y Theridiidae, es también mencionada para el Parque nacional Mburucuyá (Rubio et al. 2008).
La presencia de Linyphiidae y Lycosidae no superan juntas el 45% en áreas protegidas, y alcanzan el 85% en áreas degradadas (Ruzika 1987). La abundancia de Linyphiidae y Lycosidae representan menos de la cuarta parte de las capturas en Carlos Pellegrini, Paraje Galarza y Estancia Rincón, lo que indicaría un bajo grado de alteración en las localidades estudiadas. Estos resultados son similares a los hallados en dos bosques húmedos en el departamento capital de la provincia de Corrientes (Avalos et al. 2007), y menores en relación a los obtenidos por Pérez-Miles et al. (1999) en áreas urbanizadas.
Se registró un mayor número de especies en el Parque provincial iberá con respecto a la riqueza observada en Parque nacional Mburucuyá , no obstante, la diversidad tanto del bosque como del pastizal en las tres localidades fue similar a la observada en unidades ambientales semejantes de dicho Parque (Rubio et al. 2008).
La mayor diversidad verificada en el bosque, en relación al pastizal, coincide con estudios que demuestran que existe una correlación entre la complejidad estructural del hábitat y la diversidad de especies (Uetz 1991). La abundancia del gremio de las tejedoras orbiculares y cazadoras al acecho sería una medida indirecta del importante número y variedad de insectos que son presas de estas arañas y, tal como lo señalan Liljestrhröm et al. (2002), la diversidad de gremios presentes en el área demostraría el efecto limitador de las arañas sobre las poblaciones de insectos.
Las arañas son extremadamente sensibles a los cambios en la estructura y la complejidad del hábitat (Downie et al. 1999, New 1999). Sin embargo, las diferencias significativas detectadas en la diversidad y riqueza específicas entre unidades ambientales de las tres localidades, indicarían que otros factores, no tenidos en cuenta en esta investigación, además de la estructura física del ambiente, influyen en la mayor o menor diversidad de las especies.
El menor número de especies que restan por hallar, de acuerdo a los distintos estimadores en Estancia Rincón, podría indicar que esta localidad fue suficientemente muestreada. Sin embargo también puede interpretarse como una consecuencia de la escasez de especies vegetales observadas en el bosque y la acción intensa de pastoreo en el pastizal, que se refleja también en la menor riqueza y diversidad de especies.
La alta proporción de "singletons" es una característica evidente de la fauna tropical de artrópodos en bosques y sabanas tropicales y templados (Florez 1998, Withmore et al. 2002, Sorensen et al. 2002). En iberá, la distribución de abundancia de especies en todas las localidades se ajustó al modelo de distribución logarítmica, y resultados similares se obtuvieron en estudios de la fauna de arañas en bosques neotropicales (Silva y Coddington 1996). El ajuste a este tipo de distribución caracteriza a comunidades con bajo estrés y sometidas a la influencia de escasos factores (Moreno 2001).
Diversos factores afectan la estabilidad ambiental y consecuentemente la diversidad de especies en un hábitat determinado (Whitmore et al. 2002). El incendio bianual de los pastizales de la Reserva iberá es una práctica que provoca inestabilidad en este ambiente. A pesar de esta situación, la diversidad fue alta en el pastizal, particularmente en Paraje Galarza.
La presente contribución constituye el primer estudio sobre las comunidades de arañas de la Reserva provincial iberá, asimismo, el hallazgo de especies que no estaban citadas para Argentina y para la provincia de Corrientes demuestra la importancia de las áreas protegidas como reservorio de la biodiversidad.
Agradecimientos
Los autores manifiestan su agradecimiento a Gonzalo D. Rubio por su valiosa colaboración en los muestreos y en la determinación de algunas especies, y a Miguel Simó por la lectura crítica del manuscrito. A la Secretaría General de Ciencia y Técnica, Universidad Nacional del Nordeste, Argentina, que financió esta investigación.
Resumen
Se estudió la composición y diversidad de la comunidad de arañas de la Reserva provincial iberá, Corrientes, Argentina. Se realizaron capturas en bosque y pastizal en Colonia Pellegrini, Paraje Galarza y Estancia Rincón por medio de las técnicas de muestreo: trampas "pit-fall", tamizado, golpeteo de follaje, observación directa y red de arrastre. Las arañas se agruparon en gremios y se compiló un inventario. La similitud entre localidades y unidades ambientales se midió con el índice de Jaccard, y se calcularon los índices de diversidad de Shannon, equitabilidad, dominancia de Berger-Parker, y la diversidad beta y gamma. Para estimar la riqueza de especies se utilizó Chao 1, Chao 2, Jackknife 1 y 2. En total se recolectaron 4 138 arañas, se identificaron 33 familias y 150 especies de Araneomorphae, y dos familias y dos especies de Mygalomorphae. Cinco especies son nuevos registros para Argentina y 11 para la provincia de Corrientes. Araneidae fue la familia más abundante (39.8%), seguida por Salticidae (10.9%), Anyphaenidae (7.9%), Tetragnathidae (7.4%), Lycosidae (5.5%), y las restantes familias representaron menos del 5% de la captura total. El gremio de arañas tejedoras de telas orbiculares fue el de mayor abundancia y riqueza de especies. Entre las unidades ambientales, la mayor abundancia, riqueza y diversidad se verificó en el bosque de Colonia Pellegrini y en el pastizal de Paraje Galarza. La diversidad a fue alta, representó el 89% de la diversidad gamma, y la diversidad ß constituyó el 11% restante. Según los diferentes índices se capturó entre el 67% y el 97% de las especies que están presentes en la Reserva.
Palabras clave: Araneae, bosque, pastizal, diversidad, Reserva Iberá.
Referencias
Avalos G., Rubio G. D., Bar M.E. & González A. 2007. Arañas (Arachnida, Araneae) asociadas a dos bosques degradados del Chaco húmedo en Corrientes, Argentina. Rev. Biol. Trop. 55: 899-909. [ Links ]
Avalos G., Rubio G.D., Bar M.E. & Damborsky M.P. 2006. Lista preliminar de la araneofauna (Arachnida: Araneae) del Centro-Norte de la Provincia de Corrientes, Argentina. Rev. Ibérica Aracnol. 13: 189-194. [ Links ]
Carnevali, R. 1994. Fitogeografía de la Provincia de Corrientes. Inst. Nac. de Tec. Agrop. Corrientes, Argentina. 324 p. [ Links ]
Clausen I.H.S. 1986. The use of spiders (Araneae) as ecological indicators. B. British Arachnol. Soc. 7: 83–86. [ Links ]
Coddington J.A. & H.W. Levi. 1991. Systematics and evolution of spiders (Araneae). Annu. Rev. Ecol. Syst. 22: 565-592. [ Links ]
Corronca J.A. & C.S. Abdala. 1994. La fauna araneológica de la Reserva Ecológica "El Bagual", Formosa, Argentina. Nota preliminar. Aracnología Supl. 9: 1-6. [ Links ]
Dippenaar-Schoeman A.S., A.M. van den Berg & A. van den Berg. 1989. Species composition and relative seasonal abundance of spiders from the field and tree layers of the Roodeplaat Dam Nature Reserve. Koedoe 32: 25-38. [ Links ]
Downie I.S., Wilson W.L., Abernethy V.J., McCracken D.I., Foster G.N, Ribera I., Waterhouse A. & Murphy K.J. 1999. The impact of different agricultural landuses on epigeal spider biodiversity in Scotland. J. Insect Conservation 3: 273-286. [ Links ]
Enders F. 1976. Clutch size related to hunting manner of Spider species. Ann. Entomol. Soc. Am. 69: 991-998. [ Links ]
Florez D.E. 1998. Estructura de comunidades de arañas (Araneae) en el departamento Del valle, suroccidente de Colombia. Caldasia 20: 173-192. [ Links ]
Florez D.E. 2000. Comunidad de arañas de la región Pacífica del departamento del valle del Cauca, Colombia. Rev. Colombiana de Entomol. 26: 77-81. [ Links ]
Good, P. 2000. Permutation Tests. A practical guide to resampling methods for testing hypotheses. Springer, Nueva York, EEUU. 226 p. [ Links ]
Höfer H. 1990. The spider community (Araneae) of a central Amazonian blackwater inundation forest (Igapó). Acta Zool. Fennica 190: 173-179. [ Links ]
Lande R. 1996. Statistics and partitioning of species diversity and similarities among multiple communities. Oikos 76: 5-13. [ Links ]
Liljesthröm G., E. Minervino, D. Castro & A. González. 2002. La comunidad de arañas del cultivo de soja en la provincia de Buenos Aires, Argentina. Neotrop. Entomol. 31: 197-209. [ Links ]
Maelfait J.P., R. Jocque, L. Baert & K. Desender. 1990. Heathland management and spiders. Acta Zool. Fennica 190: 261–166. [ Links ]
Moreno, C.E. 2001. Métodos para medir la biodiversidad. M & T- Manuales & Tesis. SEA, Zaragoza, España. 83 p. [ Links ]
New T.R. 1999. Untangling the web: spiders and the challenges of invertebrate conservation. J. Insect Conservation 3: 251-256. [ Links ]
Neiff, J. J. & A. S. G. Poi De Neiff. 2006. Ecorregión Esteros del Iberá, p 177-184. In La situación ambiental argentina 2005. A. Brown, U. Martinez Ortiz, M Acerbi & J. Corchera (eds.). Fundación Vida Silvestre, Buenos Aires, Argentina. [ Links ]
Pérez-Miles F., M. Simó, C. Toscano-Gadea & G. Useta. 1999. La comunidad de Araneae criptozóicas del Cerro de Montevideo, Uruguay: Un ambiente rodeado por urbanización. Physis C 57: 73-78. [ Links ]
Rubio, G.D. Corronca J. & Damborsky M.P. 2008. Do spider diversity and assemblages change in different contiguous habitats? A case study in the protected habitats of the Humid Chaco ecoregión, north-east Argentina. Environ. Entomol. 37: 419-430. [ Links ]
Russell-Smith, A. 1999. The spiders of Mkomazi Game Reserve, p 197-222. In Mkomazi: theEcology, Biodiversity and Conservation of a Tanzanian Savanna. Coe, M., N. McWilliam, G. Stone & M. Parker (eds.). Blackwell, Londres, Inglaterra. [ Links ]
Ruzicka V. 1987. Biodiagnostic evaluation of epigeic spider communities. Ekológia (CSSR) 6: 345-357. [ Links ]
Samu F. & G.L. Lövei, 1995. Species richness of a spider community (Araneae): Extrapolation from simulated increasing sampling effort. European J. Entomol. 92: 633-638. [ Links ]
Silva, D. & J.A. Coddington. 1996. Spiders of Pakitza (Madre de Dios, Peru): species richness and notes in community structure, p 241-299. In D.E. Wilson & A. Sandoval (eds.). The biodiversity of Pakitza and its environs. Smithsonian Institution, Washington DC, EEUU. [ Links ]
Sorensen L. L., J. A. Coddington & N. Scharff. 2002. Inventorying and estimating subcanopy spider diversity using semiquantitative sampling methods in an Afromontane forest. Environ. Entomol. 31: 319-330. [ Links ]
Tressens, S.G., R. O. Vanni & M.G. Lopez. 2002. Las plantas terrestres, p 201-380. In Flora del Iberá. M.M. Arbo & S.G. Tressens (eds). EUDENE, Corrientes, Argentina. [ Links ]
Uetz, G. W. 1991. Habitat structure and spider foraging, p 325-348. In S. S. Bell, E. D. McCoy & H. R. Mushinsky (eds). Habitat Structure: The physical arrangement of objects in space. Chapman & Hall, Londres, Inglaterra. [ Links ]
Whitmore C., R. Slotow, T.E. Crouch & A.S. Dippenaar-Schoeman. 2002. Diversity of spiders (Araneae) in a savanna Reserve, Northern Province, South Africa. J. Arachnol. 30: 344-356. [ Links ]
Wise, D.H. 1993. Spiders in Ecological Webs. Cambridge, Cambridge, Inglaterra. 328 p. [ Links ]
Wise, D. 2002. Efectos directos e indirectos de las arañas en la red trófica del mantillo del bosque, p. 53-55. In V Congr. Argent. de Entomol., Buenos Aires, Argentina. [ Links ]
Referencias de Internet
Colwell R.K. EstimateS: Statistical estimation of species richness and shared species from samples. 2005. Version 7.5.1. [Electronic resource]. Mode of access: http://purl.oclc.org/estimates. [ Links ]
Hammer O., D.A.T. Harper & P.D. Ryan. 2006. PAST Palaeontological Statistics, Version: 1.43. http://folk.uio.no/ohammer/past. [ Links ]
Platnick N. I. 2008. The World Spider Catalog, Version 9.0. American Museum of Natural History, online at
http://research.amnh.org/entomology/spiders/catalog/index.html

