










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 n.1-2 San José Mar./Jun. 2009
Patricia Trujillo-Jiménez1 & Miguel Ángel Castro-Lara2
1. Centro de Investigaciones Biológicas, 2Facultad de Ciencias Biológicas. Universidad Autónoma del Estado de Morelos. Av. Universidad 1001 Col. Chamilpa C.P. 62210. Cuernavaca, Morelos, México Tel (01 777) 329-70-29, Fax (01 777) 329-70-56; trujill@cib.uaem.mx
Abstract: Diet of the chub, Notropis moralesi (Pisces: Cyprinidae) in the Amacuzac River, Morelos, Mexico. Notropis moralesi, locally known as "carpa tepelneme", is a small endemic fish of the Papaloapan River. We analyzed its diet and feeding habits. Specimens were captured monthly from Amacuzac, River, Morelos, Mexico. The gut content of 294 specimens was identified to the most specific taxonomic category possible. We quantified gut content by the numerical and frequency of occurrence methods and used the Levins amplitude of trophic niche indices and MacArthur and Levin`s indices for niche overlaping. The N. moralesi diet was composed by eleven prey categories, ten of which were of animal item. Chironomids and mayfly were the most consumed prey, independently of season and fish sex or size. We found ten nutritional components in the dry season, and eight in the rainy season. The estimated amplitude of trophic niche (Levins method) revealed high values in both seasons. The diet had ten nutritional components in immatures, eight in females and five in males. A Chi-squared test showed not significant differences in the diet by development stage, sex, and season. The niche overlaping index was high (above 0.60) for all analyses. N. moralesi is a carnivorous species with entomophagus tendencies. Rev. Biol. Trop. 57 (1-2): 195-209. Epub 2009 June 30.
Key words. Carpa tepelneme, Notropis moralesi, Amacuzac River, diet, food habits, carnivorous.
La familia Cyprinidae se encuentra distribuida por todo el mundo a excepción de Sudamérica, cuenta con el mayor número de especies y la mayoría de ellas habitan en aguas continentales, comúnmente se les conoce como carpas o barbos. La abundancia y diversidad de los ciprínidos en muchos ríos sugieren que ellos probablemente tienen un papel importante en aspectos tróficos dentro de la estructura de las comunidades, ya sea como predadores o como presas (Jordan 1989). En México es una de las familias con mayor diversidad, tanto en géneros como en especies endémicas. El género Notropis está representado por al menos 20 especies, de las cuales solamente cinco están distribuidas entre el centro y sur de México Schönhuth y Doadrio 2003).
Las numerosas especies que integran la familia Cyprinidae se han adaptado a una amplia variedad de ambientes, lo que está directamente relacionado con la alta variabilidad morfológica y sus numerosas especializaciones tróficas que presentan (Werner 1977, Gatz 1979a, 1979b, Matthews 1982, Schmitt y Coyer 1983, Miranda y Escala 2005). Con base en los estudios sobre la dieta de los ciprínidos se ha reportado que estos ocupan diversos niveles tróficos, por lo que se han considerado como especialistas y generalistas, muchas de las especies se alimentan de productores secundarios: zooplancton, crustáceos, larvas, pupas y adultos de insectos, oligoquetos, etc., también, hay especies que consumen productores primarios (macrofitas y fitoplancton), o incluso productores terciarios (Ej. peces). Según el comportamiento alimenticio, los ciprínidos pueden ser clasificado en tres niveles tróficos: herbívoro, alimentadores pelágicos, y alimentadores bentónicos (Heins y Clemmer. 1975, De Silva et al. 1980, Angermeier 1982, Surat et al. 1982, Castelló 1983, Chen et al 1984, Winfield y Nelson 1991, Nelson 1994)
En México Notropis moralesi, es conocida comúnmente como carpa "tepelneme" en español y como chaca cuachi (chaca = pescado y cuachi = chico) en mixteco, es una especie endémica de las partes altas de las cuencas del Papaloapan, Balsas y Atoyac (Miller 1986, Martínez 1999 y 2002, Schönhuth y Doadrio 2003). Habita ambientes lóticos como manantiales y ríos de montaña, está conformada por poblaciones con individuos de talla pequeña, con una longitud máxima de 97 mm y presentan un marcado dimorfismo sexual.
A pesar de la amplia distribución que presenta esta especie, se encuentra registrada en la Norma Oficial Mexicana NOM-059ECOL-2001 en la categoría de amenazada y en la "Lista roja" IUCN (2006) como en peligro crítico. La categoría de amenazada se le otorgó debido a que de acuerdo a la regionalización hecha por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), únicamente la incluían en una región hidrológica prioritaria de la región Pacífico Tropical, la cual está considerada como área amenazada.
No obstante a lo anterior, se conoce muy poco sobre la especie, por lo que es indispensable hacer investigación básica sobre su biología y ecología que aporte información que permita, llevar a cabo estrategias para su conservación, por lo que el propósito de este estudio es contribuir con información sobre su dieta y hábitos alimentarios, y determinar el reparto del alimento por épocas del año, tallas y sexos.
Materiales y métodos
El presente estudio se realizó a lo largo de 80 km del río Amacuzac, Morelos, México entre las coordenadas 19º 13´ 12" - 17º 53´ 24" de latitud norte y 99º 42´ 36" - 98º 37´ 48" de longitud oeste, abarcando un total de ocho localidades representativas de las partes alta, media y baja del ecosistema. Se realizaron mensualmente entre septiembre de 1999 a agosto de 2000 recolectas de ejemplares de N. moralesi (n=294) con una red de arrastre de 10 m de largo y 2.5 m de caída, con luz de malla de 2 cm, con un esfuerzo de captura de una hora en cada localidad.
A cada uno de los organismos se les tomo la longitud patrón (LP) en milímetros con el uso de un calibrador digital, el peso húmedo se determinó con una proximidad a décimas de gramo con una balanza analítica, el sexo se estableció de acuerdo al análisis de las gónadas. Los tractos digestivos se extrajeron siguiendo la técnica usada por Trujillo-Jiménez (1991), mismos que fueron depositados en frascos con formaldehído al 10% para su posterior análisis (Windell y Bowen 1978).
El análisis del contenido estomacal se realizó con un microscopio estereoscópico. Los componentes alimenticios se identificaron hasta los táxones más particulares posibles, lo cual dependió entre otras cosas de la integridad de las presas en los tubos digestivos. Para la identificación de los componentes alimenticios se utilizaron las claves de Bland (1979), Chu (1979), Needham y Needham (1982), MacCafferty (1983), Merrit y Cummins (1996) y Pennak (1990).
Para la cuantificación de los contenidos estomacales se aplicaron dos métodos: el numérico por cuadrícula (No.) y el de frecuencia de preferencia (F.O.) (Lagler 1956, Windell y Bowen 1978), se usó el índice de Albertaine (1973) el cual establece una escala de valores de 0 a 0.10 que ubican al componente alimenticio como incidental; de 0.11 a 0.50 como ocasionales o secundarios y de 0.51 a 1.0 como preferentes. El análisis de la dieta se hizo general (no se realizó por localidad, debido a que el número de ejemplares recolectados en cada una fue diferente), por temporadas (estiaje y lluvias) sexos (inmaduros, hembras y machos) y clases de talla (establecidas aleatoriamente de acuerdo a las longitudes registradas). Para examinar la existencia de cambios estacionales, sexos y clases de talla en la dieta se aplicó una prueba de X2. Se consideraron diferencias significativas al nivel de significación p<0.05 (Siegel 1986).
Se realizó una evaluación de la amplitud del espectro trófico (amplitud de nicho) con el fin de determinar el grado de especialización en la dieta de N. moralesi. Para tal efecto se utilizó la medida de Levin (1968), estandarizada por el método de Hulbert (1978) utilizando una escala de 0 a 1, considerándose como especialista cuando el valor es cercano a cero y generalista cuando es próximo a 1 (Ludwing y Reynolds 1988). Para medir el traslape de nicho trófico se utilizó el índice de MacArthur y Levin (1967) modificado por Pianka (Krebs 1999), que presenta un intervalo de 0 (recursos no usados en común) a 1 (traslape máximo).
Resultados
Composición general de la dieta: De acuerdo con el análisis del contenido estomacal, en 294 ejemplares de Notropis moralesi se observó que la dieta general estuvo constituida por once componentes de los cuales diez fueron de origen animal (Cuadro 1). Con base a la agrupación de los diferentes componentes alimenticios por su origen animal y vegetal se registró que los primeros tuvieron los porcentajes de ingestión y preferencia más altos. Entre los componentes de origen animal los dípteros quironómidos fueron los que presentaron los porcentajes de ingestión (27.9% No.) y de preferencia (53.3% F.O.) más altos, y de acuerdo con el índice de Albertaine son considerados como alimento preferente. Así mismo, los efemerópteros baetidos, dípteros simúlidos y el tricóptero Leptonema sp., exhibieron porcentajes de ingestión y preferencia relativamente altos (16.7, 9.6 y 8.7% No. respectivamente; 23.2, 12.3 y14.0% F.O. en el mismo orden) considerados por esto como alimentos secundarios, el resto de los componentes de origen animal y los restos de vegetales mostraron porcentajes de ingestión y preferencia bajos, por lo que son alimentos incidentales (Cuadro 1). De acuerdo con los resultados con el índice de amplitud de Levin se obtuvo un valor de 0.59.
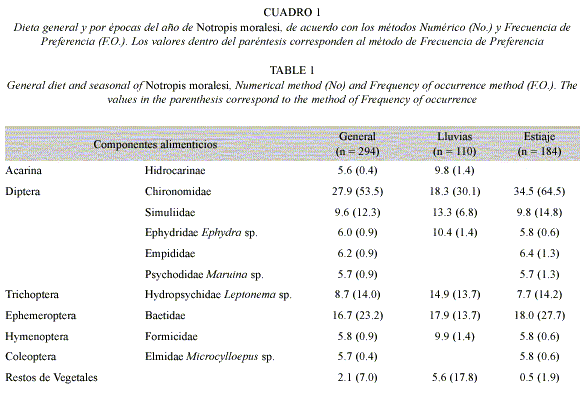
Análisis temporal de la dieta: Al realizar el análisis por épocas del año (lluvias y estiaje) se encontró algunas diferencias dentro de la dieta. De acuerdo al análisis en la época de lluvias que corresponde de mayo a octubre (110 ejemplares), se registraron ocho componentes alimenticios, de los cuales siete fueron de origen animal y uno vegetal, reconociendo a los dípteros quironómidos y los efemerópteros con los porcentajes más altos (18.3 y 17.9% No. respectivamente; 30.1 y 13.7% F.O. en el mismo orden), seguidos por los tricópteros (14.9% No. y 13.7% F.O.) y los dípteros simúlidos (13.3% No. y 6.8% F.O.). El resto de los componentes de origen animal mostraron los porcentajes de ingestión y preferencia bajos. En cuanto al componente de origen vegetal este presentó un valor de ingestión bajo (5.6%) y de preferencia relativamente alto (17.8%) por lo que es contemplado como alimento secundario (Cuadro 1). El índice de amplitud de nicho trófico, reportó un valor de 0.89, lo que representa un nicho trófico amplio.
Para la época de estiaje (noviembre a abril), se analizaron 184 ejemplares, exhibiendo diez de los once componentes de la dieta general, observándose que los dípteros quironómidos y los efemerópteros fueron los que presentaron las cifras de ingestión (34.5 y 18.0% respectivamente) y de preferencia (64.5 y 27.7% en el mismo orden) más altas, seguidos de los dípteros simúlidos y los tricópteros con porcentajes de ingestión (9.8 y 7.7% respectivamente) y de preferencia (14.8 y 14.2% en el mismo orden) relativamente altos, considerados como alimentos secundarios. El resto de los componentes de origen animal y vegetal exhibieron porcentajes de ingestión y preferencia bajos (Cuadro 1). El resultado de la prueba de X2 entre estiaje y lluvias demostró que las diferencias no son significativas (p>0.05). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.49. De acuerdo con los resultados con el Índice de traslape, se obtuvo un valor de 0.79, lo que representa un traslape amplio entre las épocas del año.
Análisis de la dieta por tallas: Para la clase de talla I (12-31 mm), se analizaron 178 ejemplares, registrándose once artículos alimenticios, siendo los dípteros quironómidos y los efemerópteros los que mostraron los valores de ingestión (30.2 y 17.3% respectivamente) y de preferencia (57.9 y 25.5% en el mismo orden) más altos, seguidos de los dípteros simúlidos (10.0% de ingestión, 15.9% de preferencia) y los tricópteros ((17.3 y 25.5% en el mismo orden), el resto de los componentes de origen animal y los restos de vegetales presentaron valores de ingestión y preferencia bajos, por lo que son estimados como alimentos incidentales (Cuadro 2). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.54, lo que representa un nicho trófico relativamente amplio.

Con base en el análisis de 93 ejemplares correspondientes a la clase de talla II (32–50 mm), se registró un total de ocho de los once componentes de la dieta general, registrándose a los dípteros quironómidos, efemerópteros y tricópteros con los porcentajes de ingestión (24.2, 19.5 y 11.9% respectivamente) y de preferencia (40.6, 15.6 y 18.8% correspondientemente) más altos, siendo considerados como alimentos secundarios. El resto de los componentes alimenticios de origen animal, exhibieron porcentajes de ingestión relativamente altos y valores bajos de preferencia considerándose alimentos incidentales. En cuanto a los restos de vegetales superiores, estos registraron porcentajes de ingestión (6.1%) y preferencia (15.6%) relativamente altos (Cuadro 2). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.80.
Para la clase de talla III (51-69 mm), se analizaron un total de 23 ejemplares, presentando únicamente cinco componentes alimenticios, siendo los dípteros quironómidos los que exhibieron las cifras de ingestión ((40.7%) y preferencia (63.2%) más altos lo cual los ubica como un alimento preferente, seguidos de los simúlidos, efemerópteros y tricópteros, que presentaron valores de ingestión (22.7, 19.3 y 16.6% respectivamente) y preferencia (10.5, 31.6 y 21.1% en el mismo orden) relativamente altos ubicándolos como alimentos secundarios. Los restos de vegetales superiores de acuerdo con el método de ingestión mostraron un porcentaje bajo (0.7%), sin embargo, de acuerdo con los resultados de frecuencia (15.8%) son estimados como alimentos secundarios (Cuadro 2). El resultado obtenido con el índice de amplitud de nicho trófico fue alto (0.64). El resultado de la prueba de X2 demostró que las diferencias entre tallas no son significativas (p>0.05). En cuanto al análisis de las combinaciones por tallas, con el Índice de traslape se registró que todas las tallas presentan traslape entre si (I - II 0.73; I - III 0.68 y II - III 0.63), representando traslapes amplios.
Análisis de la dieta por tallas y épocas del año: La dieta en la clase de talla I en la época de lluvias estuvo constituida de seis artículos alimenticios, de los cuales, cinco fueron de origen animal y uno vegetal. Todos los componentes de origen animal exhibieron porcentajes de ingestión altos, sin embargo, sólo los quironómidos y los tricóptero (Leptonema sp.) presentaron porcentajes de preferencia altos (47.6 y 28.6% respectivamente) considerados como alimentos secundarios, el resto de los componentes de origen animal y los vegetales, mostraron porcentajes bajos y fueron contemplados como alimentos incidentales (Cuadro 3). El resultado obtenido con el índice de amplitud de nicho trófico fue amplio (0.73).
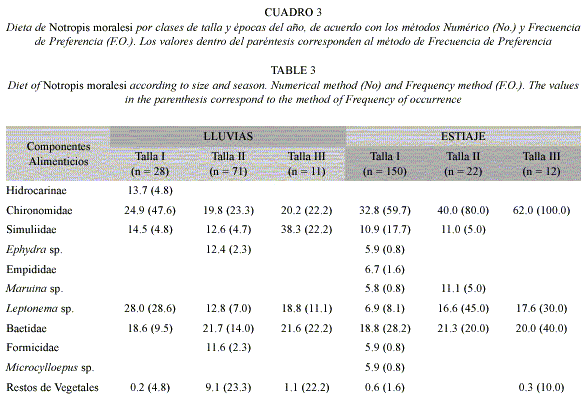
La clase de talla II presentó siete componentes alimenticios, registrando a los de origen animal con valores de ingestión altos y sólo los quironómidos y efemerópteros exhibieron porcentajes de preferencia relativamente altos (23.3 y 14.0% correspondientemente). Los restos de vegetales, registraron valores de ingestión (9.1%) y preferencia (23.3%) relativamente alto, que al igual que los quironómidos y efemerópteros son considerados alimentos secundarios. (Cuadro 3). El resultado obtenido con el índice de amplitud de nicho trófico fue alto (0.90).
La clase de talla III presentó cuatro componentes de origen animal con valores de ingestión y preferencia altos y uno de origen vegetal con porcentaje de ingestión bajo (1.1%) y un valor de preferencia alto (22.2%), considerados todos como alimentos secundarios (Cuadro 3). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.54.
En cuanto a la época de estiaje, la clase de talla I presentó un total de diez componentes de los cuales nueve son de origen animal y uno vegetal. En cuanto a los primeros se observó a los dípteros quironómidos, con porcentajes de ingesta (32.8%) y preferencia (59.7) altos siendo considerados como alimento preferente, seguidos de los efemerópteros (Baetidae) (17.3% No. y 28.2% F.O.) y dípteros simúlidos (10.9 y 17.7% en el mismo orden) por lo que se les ubica como alimentos secundarios. El resto de los componentes de origen animal y los restos de vegetales presentaron valores de ingestión y preferencia bajos, por lo que son alimentos incidentales (Cuadro 3). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.51.
La clase de talla II exhibió cinco componentes alimenticios, registrando a los dípteros quironómidos con los valores de ingestión (40.0%) y preferencia (80.0%) más altos considerados como alimento preferente, seguidos de los efemerópteros (21.3% No. y 20.0% F.O.) y tricópteros Leptonema sp (16.6 y 45.0% respectivamente), los cuales son alimentos secundarios. Los dípteros Maruina sp. y los simúlidos presentaron valores de ingestión (11.1 y 11.0% en el mismo orden) relativamente altos, no obstante, debido a sus valores de preferencia (5.0% ambos alimentos) bajos son considerados como alimentos incidentales (Cuadro 3). El resultado obtenido con el índice de amplitud de nicho trófico fue alto 0.72.
La clase de talla III presentó solo cuatro componentes, registrando a los dípteros quironómidos con los valores de ingesta (62.0%) y preferencia (100%) más altos considerados por esto como alimento preferente, seguidos de los efemerópteros (20.0% No. y 40.0% F.O.) y tricópteros (17.6 y 30.0% respectivamente) que de acuerdo a los valores de preferencia que exhibieron son alimentos secundarios. Los restos de vegetales superiores mostraron valores de ingestión y preferencia bajos por lo cual son alimento incidental (Cuadro 3). El resultado de la prueba de X2 demostró que las diferencias entre tallas en diferentes épocas no son significativas (p>0.05). El resultado obtenido con el índice de amplitud de nicho trófico fue bajo (0.40).
En cuanto al análisis de las combinaciones por tallas por época del año, con el Índice de traslape se registró que todas las tallas presentan traslape tanto en lluvias (I -II 0.91; I –III 0.98 y II – III 0.66) como en el estiaje (I -II 0.72; I –III 0.84 y II – III 0.85) representando traslapes amplios.
Análisis de la dieta por sexos: Los ejemplares inmaduros (160 ejemplares) presentaron diez componentes alimenticios, los dípteros quironómidos, los efemerópteros y los dípteros simúlidos con porcentajes de ingestión (32.7, 17.1 y 10.5% respectivamente) y de preferencia (57.0, 22.7 y 15.6% en el mismo orden) más altos, por lo que los primeros son alimento preferente y los efemerópteros junto con los simúlidos son secundarios, el resto de los componentes de origen animal junto con el de origen vegetal mostraron valores de ingestión y preferencia bajos, por lo que son alimentos incidentales (Cuadro 4). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.53.

La dieta de las hembras (76 ejemplares) estuvo constituida por ocho componentes alimenticios, de los cuales los dípteros quironómidos, efemerópteros, tricópteros y dípteros simúlidos exhibieron los porcentajes de ingestión (24.7, 19.1, 12.9 y 12.3% correspondientemente) y preferencia (48.2, 25.0, 21.4 y 8.9% en el mismo orden) más altos, por lo que son alimento ocasional o secundario, excepto los dípteros simúlidos que son ubicados como incidentales. El componente vegetal obtuvo porcentajes de ingestión y preferencia bajos por lo cual son considerados como un alimento incidental (Cuadro 4). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.53.
En los machos (58 ejemplares) se presentaron un total de cinco componentes alimenticios de origen animal y uno de origen vegetal, los porcentajes de ingestión y preferencia más elevados fueron registrados por los dípteros quironómidos (32.4% No. y 50.0% F.O.) ubicados como alimento preferente, seguidos por los efemerópteros (26.4% No. y 22.7% F.O.) y tricópteros Leptonema sp (18.1 y 18.2% respectivamente), siendo considerados como alimentos secundarios, así mismo, los dípteros simúlidos exhibieron un porcentaje de ingestión (17.6%) relativamente alto, sin embargo, debido al valor bajo de preferencia (6.8%) que mostraron son alimento incidental. En cuanto al componente de origen vegetal, presentó un porcentaje de ingestión (5.5%) bajo, mientras que el valor de preferencia (18.2%) que registro fue relativamente alto lo que los ubica como un alimento secundario (Cuadro 4). El resultado de la prueba de X2 demostró que las diferencias entre sexos no son significativas (p>0.05). El resultado obtenido con el índice de amplitud de nicho trófico fue alto (0.79). Los valores del Índice de traslape por sexos fueron amplios (Hembras – Machos 0.66; Hembras – Inmaduros 0.64 y Machos – Inmaduros 0.61).
Análisis de la dieta por sexos y épocas del año: la dieta de los inmaduros (n=139) en la época de lluvias, estuvo constituida por cinco componentes, registrando valores de ingestión altos, no obstante, sólo los quironómidos, los tricópteros Leptonema sp y efemerópteros presentaron porcentajes de preferencia (38.5, 38.5 y 15.4% respectivamente) relativamente altos, mientras que los ácaros hidrocáridos y los dípteros simúlidos mostraron valores de preferencia (7.7% cada uno) bajos por lo que son alimentos incidentales (Cuadro 5). El resultado obtenido con el índice de amplitud de nicho trófico fue alto (0.90).
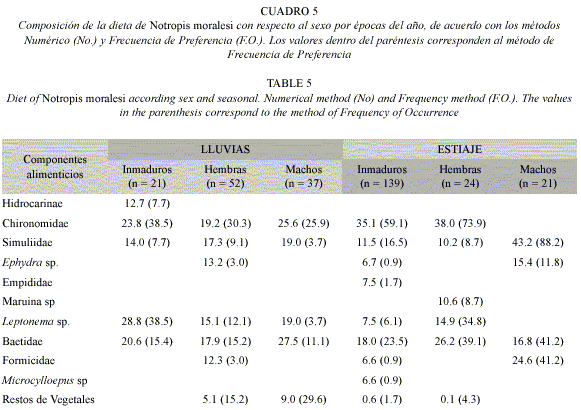
Con lo que respecta a las hembras (lluvias) (n=52) siete componentes estuvieron presentes, seis corresponden a insectos y uno a restos de vegetales superiores, en cuanto a los componentes de origen animal, todos presentaron valores de ingestión relativamente altos, mientras que solamente los quironómidos, tricópteros y efemerópteros exhibieron porcentajes de preferencia (30.3, 12.1 y 15.2% respectivamente) relativamente altos, ubicándolos como alimentos secundarios, el resto de los componentes de origen animal mostraron valores de preferencia bajos por lo que son alimentos incidentales. El componente de origen vegetal presentó un porcentaje de ingesta bajo (5.1%), no obstante, debido a su valor de preferencia (15.2%) relativamente alto, es considerado alimento secundario (Cuadro 5). El resultado obtenido con el índice de amplitud de nicho trófico fue alto 0.90.
Los machos (n=37) en las lluvias presentaron un número reducido de componentes en su dieta (5), cuatro de de origen animal y uno vegetal, todos los componentes alimenticios presentaron valores de ingestión relativamente altos, sin embargo, los quironómidos, efemerópteros y los restos de vegetales por sus valores de preferencia (25.9, 11.1 y 29.6% respectivamente) relativamente altos fueron considerados como alimentos secundarios, mientras que los simúlidos y tricópteros, por sus porcentajes de preferencia (3.7% cada uno) bajos son incidentales (Cuadro 5). El resultado de la prueba de X2 demostró que las diferencias entre sexos por épocas de año no son significativas (p>0.05). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.88, representando un nicho trófico amplio.
En el análisis de la dieta por sexos durante el estiaje, los inmaduros presentaron una dieta constituida por nueve componentes alimenticios, registrando a los dípteros quironómidos con los porcentajes de ingestión (35.1%) y preferencia (59.1%) más altos, lo cual lo ubica como alimento preferente, seguidos de los efemerópteros (18.0% No. y 23.5% F.O.) y los dípteros simúlidos (11.5 y 16.5% respectivamente) considerados por esto como alimentos secundarios, el resto de los componentes de origen animal y los vegetales presentaron valores de ingesta y preferencia bajos por lo que son ubicados como alimentos incidentales (Cuadro 5). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.52.
En lo referente a las hembras, estas presentaron seis componentes dentro de su dieta, de los cuales cinco fueron insectos y uno vegetales, el grupo de alimentos de origen animal exhibieron porcentajes de ingestión relativamente altos, en cuanto a los valores de preferencia los dípteros quironómidos, tricópteros y efemerópteros (73.9, 34.8 y 39.1% respectivamente) estos fueron altos, considerados a los quironómidos como alimento preferente y a los dos restantes como secundarios, sin embargo, los simúlidos y los dípteros Maruina sp, mostraron valores de preferencia bajos (8.7% cada uno) por lo que son incidentales, al igual que los vegetales (Cuadro 5). El resultado obtenido con el índice de amplitud de nicho trófico fue de 0.58.
Los machos presentaron cuatro componentes de origen animal, todos registraron porcentajes de ingestión altos, en cuanto a los valores de preferencia los quironómidos exhibieron el valor más alto (88.2%) por lo que son considerados como alimento preferente, el resto de los componentes alimenticios mostraron porcentajes de preferencia relativamente altos por lo que son considerados como alimentos secundarios (Cuadro 5). Las diferencias por sexos en la época de estiaje no son significativas (p>0.05). El resultado obtenido con el índice de amplitud de nicho trófico fue alto (0.78).
En cuanto al análisis de las combinaciones por sexos por época del año, con el Índice de traslape se registró un traslape amplio tanto en lluvias (Hembras – Machos 0.95; Hembras – Inmaduros 0.92 y Machos – Inmaduros 0.84) como en el estiaje (Hembras – Machos 0.70; Hembras – Inmaduros 0.82 y Machos – Inmaduros 0.78).
Discusión
En algunos estudios realizados con ciprínidos en pequeños ecosistemas acuáticos, se ha reportado que exhiben una considerable plasticidad alimenticia, registrándose que sus dietas están constituidas por microalgas, semillas, vegetales, detritos, arácnidos, crustáceos, larvas y adultos de diferentes especies de insectos acuáticos y terrestres, indicando una tendencia general a la omnivoría (Starrett 1950, Withaker 1977, Jordan 1989). No obstante, a lo anterior, el análisis cualitativo y cuantitativo del contenido estomacal de Notropis moralesi a lo largo del río Amacuzac, Morelos, registró que la dieta estuvo compuesta por once componentes alimenticios, de los cuales diez fueron insectos, por lo que en este trabajo es considerada como una especie carnívora. Análogamente, Mendelson (1975), reportó la dieta de cuatro especies del género Notropis en un río de Wisconsin (Roxbury Creek) y menciona que la dieta de estas especies está formada principalmente por componentes de origen animal, primordialmente invertebrados. Existen diversos trabajos de N. volucellus (Moyle 1973, Gascon y Leggett 1977, Magnin et al. 1978, Johnson y Dropkin 1992), en donde se menciona que la dieta de esta especie está constituida por componentes de origen animal como cladóceros, anfípodos dípteros, megalópteros y larvas de peces (Alosa sapidissima). Jordan (1989) señaló que la dieta de N. longirostris; N. venustus; N. texanus e Hybopsis winchelli está constituida de insectos terrestres (Solenopsis spp y dípteros adultos), coleópteros adultos, ninfas de efemerópteros, larvas de lepidópteros y microalgas por lo que son especies reportadas como carnívoras, al igual que N. moralesi en el presente estudio. Dahle (2001) reporta que el 75% de la dieta de Notropis topeka en el suroeste de Minnesota, estuvo constituida por microcrustáceos e insectos, el resto fueron plantas vasculares, algas y detrito, no obstante, más del 70% de la dieta fueron de origen animal, este autor considera a N. topeka como una especie omnívora generalista, a diferencia de N. moralesi, que es considerada como carnívora entomófaga ya que su dieta estuvo representada por insectos.
Hurlbert (1978), menciona que todos los valores de amplitud de nicho trófico cercanos a 0 indican que los organismos ingieren pocos componentes alimenticios (mínima amplitud de nicho trófico, máxima especialización) y los cercanos a 1, un nicho trófico amplio (mínima especialización), a pesar de que el valor de amplitud del nicho trófico obtenido en el presente estudio fue relativamente alto (0.59) y de acuerdo a lo anterior, representa una mínima especialización. La dieta de N. moralesi estuvo constituida por el 97.88% de insectos, por lo que sugiere que esta población se podría considerar como especialista, sin embargo, no se especializa en un orden ó especie, ya que se alimenta de diez diferentes especies de insectos, observándose a los quironómidos y efemerópteros con los porcentajes mayores, mientras que el resto de los componentes alimenticios, presentaron porcentajes de ingestión relativamente homogéneos.
Acorde a los componentes alimenticios dentro de la dieta de N. moralesi, se puede determinar que esta especie presentó hábitos alimentarios de tipo bentófago, ya que la mayoría de los alimentos de la dieta se encuentran en el fondo del ecosistema, excepto los himenópteros que son terrestres y los vegetales que ocupan la totalidad de la columna de agua. Granado-Lorencio y García-Novo (1986), mencionan que el principal factor que permite a los ciprínidos explotar de manera exitosa tanto organismos bentónicos como detritos, semillas ó materia vegetal, es la posesión de diferentes estructuras y mecanismos, entre los que cabe destacar: labios sensoriales y barbillones, mandíbulas protráctiles, succión lenta, selección interna por el órgano palatal, y aparato de masticación faríngeo. Encina y Granados-Lorencio (1990) reportan que Barbus bocagei presenta más desarrollados los barbillones o barbillas, que actúan como órganos quimiotáctiles e intervienen en la detección de las presas; permitiendo una más fácil localización de los organismos del bentos, lo que parece estar correlacionado con los datos obtenidos acerca de su composición alimenticia (el alimento principal fueron las larvas de quironómidos), por lo que consideran que esta especie tiene hábitos alimentarios preferentemente bentófagos, al igual que N. moralesis, sin embargo, se deben de realizar estudios sobre la ecomorfología de esta última especie para poder asegurar que sus barbillas presentan la misma importancia para la obtención de su alimento que B. bocagei.
El análisis de amplitud de nicho trófico por épocas del año, exhibió que existen diferencias en la dieta ya que en la época de lluvias se registró una amplitud de nicho alto (0.89), mientras que en la época seca se registró un valor bajo (0.49), esto posiblemente se deba a que en el estiaje el número de componentes alimenticios fue menor que en la época de secas, no obstante, todos los componentes de alimentación de la época de lluvias presentaron valores relativamente altos, a diferencia de la época seca ó estiaje en donde los quironómidos registraron un porcentaje mayor (34.5%) seguidos de los efemerópteros (18%), observándose que la dieta en esta temporada se basó principalmente en dos componentes alimenticios. Jordan (1989), reportó que la dieta de N. longirostris; N. venustus; N. texanus e Hybopsis winchelli, presentó variabilidad estacional alta, así mismo, Angermier (1982) sugiere que la plasticidad conductual (ejemplo: Hábitos alimentarios oportunistas) exhibido por los ciprínidos es debido a los sucesos que se presentan en los sistemas lóticos. Lo anterior, también se ha observado en otro grupo de peces en el río Amacuzac, como los ciclídos (Trujillo-Jiménez 1996 y 1998b) quien menciona que la dieta del convicto Cichlasoma nigrofasciatum, presenta variaciones alimenticias por un incremento de componentes en la época de lluvias, a diferencia del presente estudio, que fue en la época de secas cuando incrementó el número de componentes alimenticios, Trujillo-Jiménez (1996) también señala que las variaciones de la dieta del convicto por temporadas están relacionadas con las fluctuaciones ambientales que se presentan en el ecosistema y con la preferencia y abundancia de plantas e insectos, observando que el río Amacuzac tiene fuertes cambios, dado que en la época lluviosa muestra un abundante caudal de agua turbia, que dificulta la existencia de vegetación acuática sumergida, en cambio abundan larvas de insectos y, además existe una mayor área inundada que cubre porciones con vegetación terrestre que es utilizada por los peces como alimento. Esta plasticidad trófica es un resultado concordante con lo que la teoría predice en el caso de especies que habitan en ecosistemas cambiantes; éstas han de adaptarse a los cambios de recursos con cambios en sus preferencias alimenticias (Angermier 1982).
Los resultados con el análisis de amplitud de nicho, aumento conforme los componentes alimentarios disminuyeron, lo cual ocurrió con el crecimiento, registrándose un valor de 0.54 para la talla pequeña (I) y mayor en las tallas más grandes (II y III). La talla I presentó el número mayor de componentes alimenticios, no obstante, los porcentajes de ingestión mas altos fueron exhibidos en dos componentes alimenticios (quironómidos y efemerópteros) a diferencia de las otras dos tallas, que exhibieron un número menor de componentes alimenticios, pero sus porcentajes de ingestión fueron relativamente altos en la mayoría de los alimentos. El mismo comportamiento fue observado en Barbus sclateri, (Encina y Granado-Lorencio 1997), Cichlasoma nigrofasciatum, Poeciliopsis gracilis y Poecilia sphenops (Trujillo-Jiménez 1998a). No obstante, este no es un comportamiento generalizado en los peces, ya que en algunas especies exhiben todo lo contrario, como en el caso de N. venustus y N. texanus (Jordan 1989), Ilyodon whitei y Heterandria bimaculata (Trujillo-Jiménez, 1998a), quienes al alcanzar tallas mayores presentaron un aumento en la ingesta de los componentes alimenticios.
Con lo referente a las tallas y la época del año, se registraron ciertas variaciones, observándose que la clase de talla I en la temporada lluviosa presentó un número menor de componentes de la dieta a diferencia de la temporada seca, observándose que la ingestión de los dípteros quironómidos fue mayor en el estiaje en todas las tallas, a diferencia de la época de lluvias en donde se registró en todas las tallas a los tricópteros con los porcentajes mayores, esto es debido a que en la época lluviosa los quironómidos son arrastrados por las corrientes, puesto que no tienen estructuras especializadas que les permita asirse al sustrato, mientras que los tricópteros forman un habitáculo (canutillo), en donde se desarrollan y además les sirve de protección contra las corrientes.
Con lo que respecta al sexo, se observó que los inmaduros y las hembras fueron los que registraron el mayor número de componentes alimenticios, mientras que los machos exhibieron una marcada disminución en la composición de su dieta, sin embargo, no se observaron diferencias en cuanto a la ingestión y preferencia por los componentes alimenticios, siendo los dípteros quironómidos los más ingeridos tanto en los inmaduros como en hembras y machos. Trujillo-Jiménez y Díaz-Pardo (1996) y Trujillo-Jiménez y Toledo (2007) reportaron que no existen diferencias alimenticias por sexos en el goodeido Ilyodon whitei y los poeciliidos Heterandia bimaculata y Poecilia sphenops.
En cuanto al traslape de nicho en todas las combinaciones realizadas, tanto en épocas del año, tallas y sexos fueron altas, esto posiblemente se deba a que en todos los análisis los dípteros quironómidos y los efemerópteros fueron los que presentaron los porcentajes de ingestión mas altos, sin embargo, esto no quiere decir que los organismos compitan por este alimento ya que en los sistemas lóticos ambos grupos son los mas abundantes, como ha sido mencionado por Resh y Rosenberg (1984), McCafferty y Lugo-Ortiz 1996, McCafferty et al. (1997), Dodds (2002) y Poi de Neiff y Neiff (2006), así mismo, autores como Inger y Chin (1962); Costas y Fernando (1967); Zaret y Rand (1971) y Bishop (1973), mencionan que la segregación espacial, al menos por el uso del espacio vertical y horizontal entre las especies o incluso entre individuos coespecíficos, contribuye al reparto del recurso alimento, por lo que no existe competencia por el alimento. Así mismo, Schmitter-Soto y Castro-Aguirre (1996) estudiaron la repartición ecológica entre peces de la familia Triglidae en la costa oeste de Baja California Sur y encontraron que principalmente está determinada por factores espaciales, así como por el tipo de sedimento, la temperatura y por diferencias tróficas. Estos autores mencionan que lo anterior coincide con lo declarado por Ross (1978) y otros autores, en que el orden de importancia de los factores de repartición del recurso es espacio, dieta y tiempo; lo que se ajusta con los resultados en el presente estudio, ya que se registró la existencia de segregación espacial, así como de diferenciación alimenticia intraspecífica, lo que determina un buen reparto del recurso.
Con base en los estudios realizados sobre la dieta de los peces del río Amacuzac, Morelos (Trujillo-Jiménez 1996, 1998a y 1998b) existen dos categorías ictiotróficas. 1) Consumidores Primarios, donde se incluyen las especies detrictivoras Poeciliopsis gracilis y Poecilia sphenops (repopete) y herbívora Ilyodon whitei (mixpapal del Balsas) y 2) Consumidores Secundarios, categoría en la que se incluyen los peces predominantemente carnívoros, aun cuando pueden incluir en su dieta algunos vegetales y detritos pero sin mucha significación cuantitativa, donde se incluyen los ciclídos Cichlasoma nigrofasciatum (convicto), Cichlasoma istlaum (mojarra criolla), el bagre del balsas Ictalurus balsanus, Astyanax fasciatus (platilla), Heterandria bimaculata (repopete) y Notropis moralesi (carpa Tepelneme), esta última considerada como carnivora con tendencias a la entomofagía ya que su alimentación esta constituida por insectos, principalmente los quironómidos y los efemerópteros y presenta hábitos alimentarios bentofagos, todas estas especies representan un papel ecológico importante, en la transformación de la energía de este ecosistema.
Agradecimientos
Agradecemos a Joel Paulo Maya, Isela MolinaAstudillo, Cesar Jiménez Piedragil, Héctor Quiroz Castelán y Julio Camargo Benjumeda por la revisión crítica del manuscrito.
Resumen
Notropis moralesi, conocido comúnmente como "carpa tepelneme", es un pez endémico del río Papaloapan. Se analizó su dieta y hábitos alimentarios. Se tomaron muestras mensuales en el río Amacuzac, Morelos, México. La identificación del contenido estomacal (294 ejemplares) se llevó hasta el taxon más específico posible. Para la cuantificación se utilizaron los métodos numérico (No) y frecuencia de aparición (F.O.). Se utilizó el índice de amplitud de nicho trófico de Levin y el índice de traslape de MacArthur y Levin. N. moralesi presentó la ingestión de once componentes alimenticios, de los cuales diez son de origen animal y uno vegetal. Los quironómidos y efemerópteros fueron los más consumidos independientemente del sexo, talla y época del año. El análisis por épocas del año, reportó diez componentes en el estiaje y ocho en las lluvias. La dieta de los inmaduros estuvo constituida por diez componentes, la de las hembras por ocho y el de los machos por cinco. La prueba de Ji-cuadrada reveló que no existen diferencias significativas entre las dietas por estadio de desarrollo, sexos y épocas del año. El traslape de los nichos tróficos, reveló valores superiores a 0.60 en todas las combinaciones por sexos y épocas del año. N. moralesi es una especie carnívora con tendencias insectívoras y presenta hábitos alimentarios de tipo bentófago
Recibido 17-IV-2008. Corregido 30-VI-2008. Aceptado 29-VII-2008.
Referencias
Albertaine, B.J. 1973. Biologie des stades juveniles de teleosteens Mugilidae Mugil auratus Rissso 1810, Mugil saliens Risso 1810. I Régimen alimentaire. Aquaculture 2: 251-266. [ Links ]
Angermier, P.L. 1982. Resource seasonality and fish diets in an Illinois stream. Env. Biol. Fish. 7: 251-264. [ Links ]
Bishop, J.E. 1973. Limnology of a small Malayan river Sungai Gombak. Monographiae Biologicae No. 22. [ Links ]
Bland, R.G. 1979. How to know the immature insects. WMC Brown, Iowa, EEUU. 409 p. [ Links ]
Castelló, V.J. 1983. Utilización de técnicas multivariantes en el estudio de la variabilidad morfológica de una población de barbos (Barbus barbus sclateri G.) (Pisces, Cyprinidae) en el sur de España. His. Nat. 3: 1-15. [ Links ]
Chen, X., P. Yue & R. Lin. 1984. Major groups within the Family Cyprinidae and their phylogenetic relationships. Acta Zootaxonomica Sinica 9: 424–440. [ Links ]
Chu, H.F. 1979. The immature insects. WMC Brown, Iowa, EEUU. 243 p. [ Links ]
Costas, H.H. & C.M. Fernando. 1967. The food and feeding relationships of common meso and microfauna in the Maha Oya, a small mountainous stream at peradeniya, Ceylon. J. Sci. 7: 75-90. [ Links ]
Dahle, S.P. 2001. Studies of Topeka Shiner (Notropis topeka) life history and distribution in Minnesota. M.D. Thesis, University of Minnesota, Minnesota, EEUU. 62 p. [ Links ]
De Silva, S.S., P.R.T. Camaranatunga & C.D. De Silva. 1980. Food, feeding ecology and morphological features associated with feeding of four co-occurring cyprinids (Pisces:Cyprinidae). Neth. J. Zool. 30: 54-73. [ Links ]
Dodds, W.K. 2002. Freshwater ecology. Concepts and environmental applications. Academic, Nueva York, New York, EEUU. 569 p. [ Links ]
Encina, L. & C. Granado. 1990. Morfoecología trófica en el género Barbus (Pisces, Cyprinidae). Limnetica 6: 35-46. [ Links ]
Encina, L., & C. Granado. 1997. Seasonal changes in condition, nutrition, gonad maturation and energy content in barbel, Barbus sclateri, inhabiting in a fluctuating river. Env. Biol. Fish. 50: 75-84. [ Links ]
Gascon, D. & W.C. Leggett. 1977. Distribution, abundance, and resource utilization of littoral zone fishes in response to a nutrient/production gradient in Lake Memphremagog. J. Fish. Res. Board Can. 34: 1105-1117. [ Links ]
Gatz, A.J. 1979. Community organization in fishes as indicated by morphological features. Ecology 60: 711-718. [ Links ]
Gatz, A.J. 1979. Ecological morphology of freshwater stream fishes. Tulane Stud. Zool. Bot. 21: 91-124. [ Links ]
Granado, C. & F. García. 1986. Feeding habits of the fish community in a eutrophic reservoir in Spain. Ekol. Pol. 34: 95-110. [ Links ]
Heins, D.C. & G.H. Clemmer. 1975. Ecology, Foods and Feeding of the Longnose Shiner, Notropis longirostris (Hay) in Mississippi. Am. Midland Naturalist 94: 284-295. [ Links ]
Hurlbert, S.H. 1978. The measurement of niche overlap and some relatives. Ecology 59: 67-77. [ Links ]
Inger, R.F. & C.P. Chin. 1962. The freshwater fishes of north Borneo. Field. Zool. 45:1-268. [ Links ]
IUCN. 2006. Red list of threatened animals. International Union for Conservation of Nature and natural Resources (IUCN). Gland, Switzerland and Cambridge, UK. [ Links ]
Johnson, J.H. & D.S. Dropkin. 1992. Predation on recently released larval American shad in the Susquehanna River basin. North Am. J. Fish. Management 12: 504-508. [ Links ]
Jordan, F. 1989. The comparative feeding ecology of cyprinid fishes of the Choctawhatchee river, Florida. Thesis of Master of Science. University College of Arts and Sciences, Florida. 122 p. [ Links ]
Krebs, J. 1999. Ecological methodology. Benjamín Cummings, EEUU. 620 p. [ Links ]
Lagler, K.F. 1956. Freshwater fishery biology. WMC Brown, Iowa, EEUU. 421 p. [ Links ]
Levins, R. 1968. Evolution in changing environments: Some theoretical explorations. Princeton, Nueva Jersey, EEUU. 120 p. [ Links ]
Ludwing, J.A. & J.F. Reynolds. 1988. Statistical ecology. Wiley, Nueva York, EEUU. 337 p. [ Links ]
MacArthur, R.H. & R. Levin. 1967. The limiting similarity, converge, and divergence of coexisting species. Am. Nat. 101: 377-385. [ Links ]
MacCafferty, W.P. 1983. Aquatic Entomology. Jones & Bartlett, Boston, EEUU. 448 p. [ Links ]
McCafferty, W.P. & C.R. Lugo. 1996. Ephemeroptera, p. 133-145. In J.E. Llorente, A. García & E. González (eds.). Biodiversidad, taxonomía y biogeografía de artrópodos de México: Hacia una síntesis de su conocimiento. UNAM, México, D.F, México. 640 p. [ Links ]
McCafferty, W.P., C.R. Lugo, A.V. Provosha & T.Q. Wang. 1997. Los Efemerópteros de México: I. Clasificación superior, diagnosis de familias y composición. Dugesiana 4: 1-29. [ Links ]
Magnin, E., E. Murawska & A. Clement. 1978. Diet of seven littoral fishes from the Grand Arse de lile Perrot, on the St. Louis Lake, Montreal, Quebec. Canadian Naturalist 105: 89-101. [ Links ]
Martínez, R.E. 1999. Taxonomía y zoogeografía de la ictiofauna dulceacuícola del estado de Oaxaca, México. Tesis de doctorado, Universidad de Barcelona, Barcelona, España. 122 p. [ Links ]
Martínez, R.E. 2002. Notropis moralesi. Propuesta de categorías de riego para la ictiofauna dulceacuícola de Oaxaca. Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional-Oaxaca. Instituto Politécnico Nacional. Bases de datos SNIBCONABIO. [ Links ]
Mathwes, W.J. 1982. Small fish community structure in Ozark streams: structured assembly patterns or random abundance of species? Amer. Midl. Nat. 107: 43-54. [ Links ]
Mendelson, J. 1975. Feeding relationships among species of Notropis (Pisces: Cyprinidae) in a Wisconsin stream. Ecol. Monogr. 45: 199.230. [ Links ]
Merrit, R.W. & K.W. Cummins. 1996. An introduction to the aquatic insects of North America. Kendall Hunt, Iowa, EEUU. 862 p. [ Links ]
Miller, R.R. 1986. Composition and derivation of the freshwater fish fauna of Mexico. An. Esc. Nac. Cienc. Biol. 30: 121-153. [ Links ]
Miranda, R. & M.C. Escala. 2005. Morphometrical comparison of cleithra, opercular and pharyngeal bones of autochthonous Leuciscinae (Cyprinidae) of Spain. Folia Zool. 54: 173-188. [ Links ]
Moyle, P.B. 1973. Ecological segregation among three species of minnows (Cyprinidae) in a Minnesota lake. Transactions Am. Fish. Soc. 102: 794-805. [ Links ]
Needham, J.G. & P.R. Needham. 1982. Guía para el estudio de los seres vivos de las aguas dulces. Reverté, Barcelona, España. 131 p. [ Links ]
Nelson, J.S. 1994. Fishes of the World. Wiley, Nueva York, EEUU. 600 p. [ Links ]
Pennak, K. 1990. Freshwater invertebrate of the United States. Wiley, Nueva York, EEUU. 420 p. [ Links ]
Poi de Neiff, A. & J.J. Neiff. 2006. Riqueza de especies y similaridad de los invertebrados que viven en plantas flotantes de la planicie de inundación del río Paraná (Argentina). Interciencia 31: 220-225. [ Links ]
Resh, V.H. & D.M. Rosenberg. 1984. The ecology of aquatic insects. Praeger, Nueva York, EEUU. 625 p. [ Links ]
Ross, S.T. 1978. Trophic ontogeny of the leopard searobin Prinotus scitulus (Pisces:Triglidae). Fish. Bull. U.S. 76: 225-234. [ Links ]
Schmitt, R.J. & J.A. Coyer. 1983. Variation in surfperch diets between allopatry and sympatry: circumstantial evidence for competition. Oecologia 58: 402-410. [ Links ]
Schmitter J.J. & J.L. Castro. 1996. Trophic comparison among Triglidae (Pisces:Scorpaeniformes) of Baja California Sur, Mexico. Rev. Biol. Trop. 44: 803-811. [ Links ]
Schönhuth S. & I. Doadrio. 2003. Phylogenetic relationships of Mexican minnows of the genus Notropis (Actinopterygii, Cyprinidae). Biol. J. Linnean Soc. 80: 323-337. [ Links ]
Siegel, S. 1986. Estadística no paramétrica. Trillas, México D.F., México. 346 p. [ Links ]
Starrett, W.C. 1950. Food relationships of the minnows of the Des Moines River, Iowa. Ecology. 31: 216-233. [ Links ]
Surat, E.M., W.J. Matthews & R.B. Jeffrey. 1982. Comparative Ecology of Notropis albeolus, N. ardens and N. cerasinus (Cyprinidae) in the Upper Roanoke River Drainage, Virginia. Am. Midland Naturalist. 107: 13-24. [ Links ]
Trujillo, P. 1991. Variación de un ciclo anual de la dieta y hábitos alimentarios de Ilyodon whitei (Eigenman) (Pisces:Goodeidae) en el río del Muerto, Morelos. Tesis Profesional de Licenciatura, Facultad de Ciencias Biológicas, UAEM, Morelos, México. 39 p. [ Links ]
Trujillo, P. 1996. Ecología Alimentaría de "Cichlasoma (Archocentrus) nigrofasciatum" (Pisces: Cichlidae) en el río Amacuzac, Morelos. Zoología Informa 33: 13-32. [ Links ]
Trujillo, P. 1998. Dinámica Trófica de la Ictiofauna del río Amacuzac, Morelos. Tesis de Maestría, Facultad de Ciencias Biológicas, UAEM, Morelos, México. 101 p. [ Links ]
Trujillo, P. 1998. Trophic spectrum of the cichlids Cichlasoma (Parapetenia) istlanum and Cichlasoma (Arcocentrus) nigrofasciatum in the Amacuzac River, Morelos, México. J. Freshwater Ecol. 13: 465-473. [ Links ]
Trujillo, P & E. Díaz. 1996. Espectro Trófico de Ilyodon whitei (Pisces: Goodeidae) en el río Muerto, Morelos. Rev. Biol. Trop. 44: 755-761. [ Links ]
Trujillo, P. & H.B. Toledo. 2007. Alimentación de los peces dulceacuícolas tropicales Heterandria bimaculata y Poecilia sphenops (Cyprinodontiformes: Poeciliidae). Rev. Biol. Trop. 55: 603-615. [ Links ]
Werner, E.E. 1977. Species packing and niche complementarity in three sunfishes. Am. Nat. 111: 553-578. [ Links ]
Windell, J.T. & S.H. Bowen. 1978. Study of fish diets based on analysis of stomach contents, p. 219-226. In T. Bagenal (ed.). Methods for assessment of fish production in freshwater. Blackwell, Londres, Inglaterra. [ Links ]
Surat, E.M., W.J. Matthews & R.B. Jeffrey. 1982. Comparative Ecology of Notropis albeolus, N. ardens and N. cerasinus (Cyprinidae) in the Upper Roanoke River Drainage, Virginia. Am. Midland Naturalist. 107: 13-24. [ Links ]
Winfield, I. & J. Nelson. 1991. Cyprinid Fishes: Systematic Biology and Exploitation. Chapman. Nueva York, EEUU. 667 p. [ Links ]
Whitaker, J.O., Jr. 1977. Seasonal changes in food habits of some cyprinid fishes from the White River at Petersburg, Indiana. Am. Midland Naturalist 97: 411-418. [ Links ]
Zaret, T.M. & A.S. Rand. 1971. Competition in tropical stream fishes: Support for the competitive exclusion principle. Ecology 52: 336-342. [ Links ]

