










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 n.1-2 San José Mar./Jun. 2009
Jazzmin Arrivillaga1, Patricia Salerno1 & Yadira Rangel2
1. Lab. de genética de poblaciones, Ecología molecular de insectos vectores, Universidad Simón Bolívar, Caracas, Sarteneja, Venezuela; jazzmin.arrivillaga@gmail.com; patriciasalerno@gmail.com
2. Lab. de biología de poblaciones, Universidad Central de Venezuela, Caracas, Venezuela; yadira.rangel@ciens.ucv.ve
Abstract: Asymmetric reproductive isolation between Lutzomyia pseudolongipalpis and Lutzomyia longipalpis (species C2), Neotropical vectors of visceral leishmaniasis (Diptera: Pshychodidae). Lutzomyia pseudolongipalpis and Lutzomyia longipalpis (species C2, L. sp n.) are two endemic species of Phlebotominae sand fly vectors from Venezuela. The two insects are sympatric and monophyletic but have deeply diverging, morphological differences. They belong to the L. longipalpis complex. A study of their reproductive isolation is necessary to understand the process of speciation and maintenance of the two sister species as two discrete taxonomic and biological entities. Cross-mating tests were conducted (homo and hetero-specific) and monitored under two criteria: biological (presence of copulation and offspring) and genetic (using two isozymic markers diagnostic for the L. longipalpis complex; enzyme diagnosis AK and HK). Results indicate reproductive isolation, with an asymmetrical genetic exchange in a direction of hybridization between the two species under experimental conditions, and production of a low number of heterozygotes. These findings support the existence of negative selection on hybrids, and explain the absence of hybrids under natural conditions, in the sympatric locality. Rev. Biol. Trop. 57 (1-2): 23-31. Epub 2009 June 30.
Key words: Lutzomyia longipalpis complex, Lutzomyia pseudolongipalpis, reproductive isolation, Phlebotominae, speciation, sympatry, Venezuela.
El complejo "Lutzomyia longipalpis" (Lutz y Neiva, 1912) está conformado por cinco especies monofiléticas (Arrivillaga et al. 2003), identificadas por marcadores moleculares diagnóstico, ADN mitocondrial (región citocromo c unidad uno) e isoenzimas (Adenilatoquinasa, Hexoquinasa y Malato deshidrogenasa). Sin embargo,
solo una especie ha sido descrita formalmente con base en marcadores morfológicos (armadura cibarial, cercos y valvifer en la hembra adulta) para diferenciar a Lutzomyia pseudolongipalpis (Arrivillaga y Feliciangeli 2001) de las otras cuatro especies del complejo.
Históricamente, los primeros estudios de especiación dentro del complejo, fueron realizados por Ward et al. (1983, 1988) quienes señalaron la existencia de aislamiento precigó tico basado en la ausencia de progenie, entre algunas poblaciones simpátridas y alopátridas (Brasileras) de la especie A, según nominación Arrivillaga et al. (2003), las cuales presentaban fenotipos diferentes (número de manchas tergales en el macho), feromonas sexuales masculinas y sonidos de cortejo diferentes, pero no se demostró la existencia de diferencias genéticas entre las poblaciones aparentemente aisladas en términos reproductivos. La ausencia de bases genéticas robustas que apoyen este aislamiento reproductivo es evidenciada por varios autores, Mukopaday et al. (1997, 1998a, b) Mutebi et al. (1999, 2002) Soto et al. (2001), Arrivillaga et al. (2002), Hodgkrinson et al. (2003) y de Quiroz Balbino et al. (2006). A pesar de estos resultados, algunos autores han evaluado el aislamiento reproductivo dentro de poblaciones de la especie A, Bauzer et al. (2002a, b) y Bottecchia et al.( 2004).
Sin embargo, Lanzaro et al. (1993) mostraron por primera vez concordancia entre los conceptos de especie genética (basado en frecuencias alélicas por la técnica de isoenzimas) y el de especie biológica (basado en experimentos de cruzamiento), e indicaron un flujo génico restringido por mecanismos postcigóticos, dada a la producción de machos estériles en la primera generación filial, resultante de los experimentos de cruzamientos entre tres especies alopátridas del complejo (según la nominación de Arrivillaga et al. (2003) para las nuevas especies), L. longipalpis sensu estricto (especie A, poblaciones de Brasil), Lutzomyia. n. sp. (especies C1, poblaciones de Colombia), Lutzomyia n. sp. (especies C2, en parte poblaciones de Venezuela), Lutzomyia n. sp. (especie D, poblaciones de América Central). La existencia de bases genéticas para el aislamiento reproductivo entre estas tres especies ha sido apoyada por otros autores, Morrison et al. (1995), Lanzaro et al. (1998), Soto et al. (2001), Arrivillaga et al. (2002, 2003) y Watts et al. (2005). Desde el punto de vista genético y micro evolutivo, la evidencia mas robusta para hablar de especiación dentro del complejo, es el reporte de dos especies discretas en condición simpátrida (Venezuela, Edo. Lara, El Paso) L. longipalpis especie C2) y L. pseudolongipalpis, en ausencia de individuos heterocigotos en condiciones naturales (Lampo et al. 1999). Dicho aislamiento es apoyado por diferencias morfológicas que incluyen diferencias en los órganos genitales y piezas bucales de las larvas (Arrivillaga et al. 2000, Arrivillaga y Feliciangeli 2001), isoenzimas (Arrivillaga et al. 2000), micro-satélites (Watts et al. 2005), filogenéticas moleculares (Arrivillaga et al. 2002), filo-geográficas (Arrivillaga et al. 2003), ecológicas (Feliciangeli et al. 2004), diferencias biológicas en la productividad de las colonias (Feliciangeli y Arrivillaga, en prep.), y diferencias en el tipo de feromona sexual masculina (Hamilton et al. 2005).
En general se desconocen aspectos relacionados con el fundamento clave de la genética de la especiación, el aislamiento reproductivo entre las dos especies mas diferentes y divergentes del complejo, a pesar de la condición simpátrida, lo que permitiría explorar los mecanismo evolutivos potenciales que expliquen la existencia de dos especies filogenéticas hermanas, biológica y taxonómicamente válidas, dentro del complejo de especies vectores de leishmaniasis visceral en el Neotrópico. El objetivo principal del trabajo fue evaluar el entrecruzamiento entre L. pseudolongipalpis y L. longipalpis (especie C2).
Materiales y métodos
Experimentos de cruzamiento: Adultos de ambos sexos fueron inicialmente seleccionados según el criterio de rotación de los órganos genitales de los machos dentro de un periodo de dos a cuatro horas (excepto para el cruce Cc) al considerar a una hembra coespecífica estrictamente virgen para los ensayos. Experimentos de cruzamiento homoespecíficos (controles) y heteroespecíficos con cópulas recíprocas (entrecruzamiento sin selección) fueron realizados de la siguiente manera
Cruce control a (Cca): Hembras (N= 15) X machos (N=15) de L. pseudolongipalpis (generación filial 82, originaria de Curarigua, La Rinconada, Brasiliar, Lara, Venezuela, tres réplicas)
Cruce control b (Ccb): Hembras (N=15) X machos (N=15) de L. longipalpis (sexta generación filial, originaria de Las Cabreras, Margarita, Nueva Esparta, Venezuela, tres réplicas).
Cruce Ca: Hembras (N=15) de L. pseudolongipalpis (separadas entre las 2 a 4 hr de emergidas) x machos de L. longipalpis (N=15), tres réplicas.
Cruce Cb: Machos (N=15) de L. pseudolongipalpis X hembras (N=15) de L. longipalpis (separadas a las 2 a 4 hr de emergidas, tres réplicas).
Cruce Cc: Hembras (N=15) de L pseudolongipalpis (separadas a las 12 hr de emergidas) x macho de L. longipalpis (N=15), (una réplica).
Los cruces fueron realizados con grupos de individuos, por varios criterios convencionales de mantenimiento de colonias, tales como: a) con las hembras en grupos se asegura una mayor oviposición sobre el sustrato sólido (normalmente es del 60-70%), b) se estimula la toma de ingesta sanguínea masiva (70-80% sobre el hámster), c) con un grupo de machos se asegura la fertilización de las hembras (normalmente se usa una relación de 20 machos x 40 hembras por envase de cría), d) no todas las hembras que toman la ingesta sanguínea colocan huevos, e) las hembras mueren al oviponer, f) mayoritariamente las pupas separadas en forma individual no emergen, lo que no garantiza obtener un número adecuado de adultos vírgenes para los ensayos.
Análisis de isoenzimas con progenies híbridas: Los individuos híbridos obtenidos de los cruces efectivos fueron criados hasta la fase adulta, en condiciones de laboratorio controladas (humedad relativa de 80% y temperatura de 25 °C) según el protocolo de Killick- Kindrich (1977). Los adultos emergidos fueron preservados individualmente en viales secos a –80 °F para los análisis isoenzimáticos según protocolo de Arrivillaga et al. (2000).
Basado en los estudios genéticos previos dentro del complejo se seleccionaron tres enzimas para monitorear las dos especies de flebotominos. Dos enzimas diagnósticas, la adenilato quinasa (AK 2.7.4.3) y la hexoquinasa (HK 2.7.1.1), y una enzima quasidiagnóstico la glicerofosfato isomerasa (GPI 5.3.1.9). Los genotipos fueron designados de acuerdo al patrón de migración de los electromorfos, a las características de estructura de las enzimas analizadas, y a registros previos de los alelos presentes en las poblaciones venezolanas. Los datos de frecuencia alélicas obtenidas con el programa FSTA (Goudet 1999) fueron utilizadas para los análisis de conglomerados (estadística multivariante), a fin de obtener un estimado de similaridad empleando el programa GDA (Lewis y Zaykin 1999).
Resultados
Cruces homoespecíficos: Los cruces controles (Cca y Ccb) referenciales fueron efectivos con una viabilidad de híbridos hasta su fase adulta. Sin embargo, se observó una diferencia sustancial entre la productividad de los cruces, el control Cca presentó una producción promedio de adultos neta de 700 huevos, tres y medias veces mayor que la productividad neta del control Ccb (200 huevos). Los valores señalados para ambos controles están dentro del promedio de productividad de las colonias de L. pseudolongipalpis (desde la primera a la 87 generación filial) y L. longipalpis (desde la primera a la sexta generación filial) bajo las mismas condiciones de cría.
Cruce heteroespecífico, Cruce Ca: El ensayo de entrecruzamiento entre L. pseudolongipalpis hembra y L. longipalpis macho fue efectivo, con viabilidad de individuos inmaduros hasta la fase adulta, y con una productividad neta de 150 huevos, menor en comparación con los controles referenciales.
Cruce heteroespecífico, Cruce Cb: La línea híbrida de este cruce es inviable, la cópula visualizada no fue efectiva, ya que la ovipostura resultó en la producción de 13 huevos que no eclosionaron.
Cruce heteroespecífico, Cruce Cc: La línea híbrida de este cruce fue efectiva, la cópula fue visualizada, la ovipostura resultó en la producción neta de 189 huevos, menor en comparación con los controles referenciales.
Frecuencias alélicasCruces homoespecíficos: En todas las corridas electroforéticas una muestra de referencia de las especies L. longipalpis y L. pseudolongiaplpis fue incluida, para comparar los resultados de las expresiones fenotípicas de las enzimas AK, HK y GPI con los obtenidos para los adultos analizados de cada cruce. Para las enzimas diagnósticas todos los individuos de L. pseudolongipalpis (referencias y cruce Cca (parentales y primera filial) expresaron un único alelo para cada enzima (AK111 y HK118, con frecuencia alélica de 1.00, respectivamente). Los adultos de L. longipalpis (referencias y cruce Ccb (parentales y primera filial) expresaron un único alelo para cada enzima diagnóstica AK129 y HK85 (con frecuencia alélica de 1.00, respectivamente), siendo estos alelos privados y no expresados por L. pseudolongipalpis, confirmándose el carácter diagnóstico de los alelos evaluados.
Para la enzima GPI se observa que L. pseudolongipalpis (referencias y cruce Cca (parentales y primera filial) presenta un 100% de expresión homocigota del alelo GPI34 (con frecuencia alélica de 1.00). Mientras L. longipalpis (referencias y cruce Ccb (parentales y primera filial) presenta expresión homocigota del alelo quiasidiagnóstico GPi48 (con frecuencia alélica de 0.931), y del segundo alelo GPI34 (con frecuencia alélica de 0.069). En la primera generación de los experimentos controles (Cca y Ccb), todos los individuos híbridos expresaron un 100% de homocigosidad para los alelos diagnósticos de L. pseudolongiapalpis y L. longipalpis, respectivamente. Estos resultados confirman el caracter quasidiagnóstico de los alelos monitoreados para la enzima GPI, con base en sus frecuencias menores a 0.95.
Cruces heteroespecíficos sin selección
Progenie híbrida del cruce Ca: En la primera generación híbrida se monitorearon 100% de individuos heterocigotos para los alelos AK111/129 (doble banda, con frecuencia alélica de 0.5, respectivamente), HK85/118 (doble banda, frecuencia alélica de 0.5, respectivamente), y GPI34/48 (triple banda, frecuencias alélicas de 0.559 y 0.441 respectivamente), todos los alelos evidenciados para cada enzima son diagnósticos para las especies L. pseudolongipalpis y L. longipalpis, con base en las muestras de referencia y los controles Cca y Ccb. Estos resultados evidencia la formación de heterocigotos en condiciones de laboratorio (Fig. 1).

Progenie híbrida del cruce Cc: En la primera generación se observó un 100% de individuos homocigotos, con frecuencias alélicas de 1.00 para los alelos AK111 y HK118. Mientras, se observó una frecuencia de 0.800 para el alelo GPI134 y de 0.20 para el alelo GPI48 (contrario a lo obtenido en el cruce equivalente Ca). En este cruce, todos los alelos registrados son diagnóstico para la especie L. pseudolongipalpis, con base en las muestras de referencias y al control Cca. Estos resultados reflejan la ausencia de heterocigotos utilizando el criterio de virginidad en hembras igual al tiempo de 12 hr de emergidas, ya que se asumió que se necesitaba la rotación total de los órganos genitales del macho de 180º, la cual se completa después de las 12 hr de emergencia según literatura (Young y Duncan 1994).
Análisis de conglomerados: El análisis fenético (Fig. 2) indica tres grupos, el grupo mas externo, representado por los individuos obtenidos en el cruce homoespecífico Ccb, que representan a la especie L. longipalpis. El segundo grupo está conformado por los individuos obtenidos del cruce homoespecífico Cca y heteroespecífico Cc, los cuales representan geneticamente a la especie L. pseudolongipalpis, y el tercer grupo representado por los individuos obtenidos del cruce heteroespecífico Ca, los individuos heterocigotos, con una distancia significativa de 0,31 de las dos especies analizadas (representadas en los Cruces Cca y Ccb).
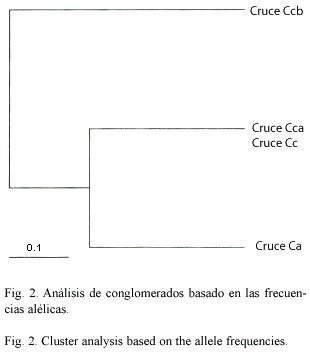
Esto implica que los híbridos se encuentra a una distancia intermedia entre ambas especies, y que la agrupación en el análisis conglomerado no sea única, la cual se realiza al azar resultando en dos dendogramas. El primer dendograma (Fig. 2) asocia a los heterocigotos con L. pseudolongipalpis, y el segundo dendograma (no mostrado) asocia a los heterocigotos con L. longipalpis. Adicionalmente, estos resultados reflejan que los heterocigotos tienen una estructuración genética diferencial de sus progenitores del cruce Ca, la cual permite monitorearlos en el campo.
Discusión
Los resultados indican un intercambio genético asimétrico entre las dos especies, basado en la inviabilidad de los huevos en uno de los cruces recíprocos (Cb), lo que apoya la existencia de mecanismos precigóticos, debido a la observación de una cópula no efectiva. Mientras, que el cruce recíproco Ca fue viable, pero con una productividad de híbridos promedio por debajo a los cruces controles, sugiriendo una selección negativa de los híbridos que apoya la existencia de mecanismos postcigóticos.
La combinación de ambos mecanismos para explicar el aislamiento reproductivo entre estos dos flebotominos neotropicales, es concordante con lo reportado para otras especies de dípteros en condición simpátrida (Orr y Coyne 1989, Reed y Markow 2004, Slotman et al. 2004), con una divergencia genética y evolutiva profunda que resulta en la asimetría en cruces recíprocos, postcruzamiento (Oliver 1978, Harrison 1983, Coyne y Orr 1989, Turreli y Orr 1995, Gallant y Fairban 1997, Presgraves y Orr 1998, Navajas et al. 2000, Willet y Burton 2001, Presgraves 2002, Turelli y oyle 2007).
En contraste, con nuestros resultados los experimentos realizados por Lanzaro et al. (1993), indican la ausencia de un intercambio asimétrico entre las especies A, C1 y D, y la existencia de huevos viables con la emergencia de machos estériles en los cruces recíprocos para todas las combinaciones ensayadas, implicando solo la existencia de mecanismos postcigóticos (Orr y Coyne 1989, Slotman et al. 2004).
Sin embargo, estos resultados lejos de ser contradictorios con los de este trabajo apoyan la conclusión planteada, ya que las tres especies genéticas (L. longipalpis sensu lato: especie A, C1 y D) entrecruzadas por Lanzaro et al. (1993) son alopátridas, están separadas por distancias genética pequeñas (distancia de Nei de 0.15) y con una baja divergencia nucleotídica (5%), lo cual apoya la existencia de mecanismos postcigóticos (Orr 1995).
En el caso de las especies L. pseudolongipalpis y L. longipalpis (especie C2), los valores de distancia genética son altos (distancia de Nei de 0.67) y existe una profunda divergencia nucleotídica (10%) que implica la existencia de mecanismos pre y postcigóticos (Coyne y Orr 1997).
Por otro lado, la baja productividad de adultos del cruce Ca, indica la existencia de selección negativa contra los híbridos, lo que disminuye la eficacia biológica de los individuos obtenidos de los cruces heteroespecíficos en comparación con los individuos derivados de cruces homoespecíficos (Coyne et al. 2002), pudiendo existir una desigualdad en los índices de competitividad ecológica (Naisbit 2001, Van Door y Wissing 2001, Mckinnon 2004, Jiggis 2005), que sumado a las diferencias en la dinámica poblacional de actividad diaria entre las dos especies (Feliciangeli et al. 2004), y a las diferencias en feromona entre estas dos especies (Hamilton et al. 2005), resulta en la diferenciación ecológica, lo que refuerza el aislamiento reproductivo en la condición simpátrida (Van Door y Wissing 2001, Via 2002, Mckinnon et al. 2004).
El orden decreciente de eficacia biológica en el sentido L. pseudolongipalpis, L. longipalpis (especie C2), y por último los individuos heterocigotos, podría estar explicando la dominancia simple (evidenciada por isoenzimas) de la especie L. pseudolongipalpis en un abundancia del 80% en la localidad simpátrida de El Paso (Lara, Venezuela), en relación a un 20% de abundancia de la especie L. longipalpis, y en ausencia total de heterocigotos (Lampo et al. 1999).
Los resultados para el cruce Cc podrían estar apuntando a la existencia de un reforzamiento del aislamiento reproductivo por preferencia y selección sexual (Price et al. 2000, 2004, Panhuis et al. 2001, Kirkpatrick y Nuismer 2004, Velthuis et al. 2005, Moehring et al. 2006). Basado en que las hembras de L. pseudolongipalpis, separadas como vírgenes a las 12 hr, fueron copuladas de forma efectiva por sus machos coespecíficos, y aunque lograron copular con los machos heteroespecíficos, no se logró una fertilización efectiva por parte de este segundo grupo de machos, dado a la ausencia de genotipos heterocigotos en las corridas electroforéticas, lo cual pudiera sugerir competencia espermática (Markowt 1997), estableciéndose la necesidad de orientar futuras investigaciones en esa dirección.
Adicionalmente, los resultados indican que no se necesita una rotación total de los órganos genitales de los machos para lograr una cópula efectiva entre los individuos coespecíficos, siendo esta variable temporal clave para los ensayos de cruzamiento.
Agradecimientos
Proyecto S1 financiado por el Decanato de investigación y desarrollo de la Universidad Simón Bolívar, bajo la responsabilidad de J. Arrivillaga. Los autores extienden su agradecimiento a Dora Feliciangeli, Hermes Piñango, y María Martínez por su colaboración durante el desarrollo del trabajo.
Resumen
Lutzomyia pseudolongipalpis y Lutzomyia longipalpis (especie C2, L. n. sp) son dos especies de flebotominos vectores endémicos de Venezuela, simpátricas, monofiléticas, con profundas divergencias, morfológicamente diferentes y pertenecientes al complejo de especies L. longipalpis. El estudio de su aislamiento reproductivo es clave para entender el proceso de especiación y el mantenimiento de estas dos especies hermanas como entidades taxonómicas y biológicas discretas. Por tal motivo, se realizaron ensayos de entrecruzamiento homo y heteroespecíficos, los cuales fueron monitoreados por dos criterios: biológico (presencia de cópula y progenie) y genético (utilizando dos marcadores isoenzimáticos diagnósticos para el complejo L. longipalpis, las enzimas AK y HK). Los resultados indican aislamiento reproductivo, con un intercambio genético asimétrico hacia la hibridización en condiciones experimentales, y producción de un bajo número de heterocigotos, lo que apoya la existencia de selección negativa sobre los híbridos y explica su ausencia en condiciones naturales en la localidad simpátrica.
Palabras clave: complejo Lutzomyia longipalpis, Lutzomyia pseudolongipalpis, aislamiento reproductivo, Phlebotominae, especiación, simpatría, Venezuela.
Recibido 14-III-2008. Corregido 14-VII-2008. Aceptado 14-VIII-2008.
Referencias
Arrivillaga, J. & M.D. Feliciangeli. 2000. Diferenciación morfológica larval entre poblaciones de Lutzomyia longipalpis (Lutz & Neiva, 1912) en Venezuela (Diptera: Psychodidae). Bol. Entomol. Venez. 15:229-234. [ Links ]
Arrivillaga, J., Y. Rangel, M. Oviedo & M.D. Feliciangeli. 2000. Genetic divergence among Venezuelan populations of Lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae). J. Med. Entomol. 37: 325-330. [ Links ]
Arrivillaga, J., & M.D. Feliciangeli. 2001. Lutzomyia pseudolongipalpis: the first new species within the longipalpis (Diptera: Psychodidae: Phlebotominae) complex from La Rinconada, Curarigua, Lara State, Venezuela. J. Med. Entomol. 38: 783-790. [ Links ]
Arrivillaga, J.C., D.E. Norris, M.D. Feliciangeli & G.C. Lanzaro. 2002. Phylogeography of the neotropical sand fly Lutzomyia longipalpis inferred from mitochondrial DNA sequences. Infect. Genet. Evol. 2: 83-95. [ Links ]
Arrivillaga, J., J.P. Mutebi, H. Piñango, D. Norris, B.Alexander, M.D. Feliciangeli & G.C. Lanzaro. 2003. The taxonomic status of genetically divergent populations of Lutzomyia longipalpis (Diptera: Psychodidae) based on the distribution of mitochondrial and isozyme variation. J. Med. Entomol. 40: 615-627. [ Links ]
Bauzer, L.G., N.A. Souza, R.D Ward, C.P. Kyriacou & A.A Peixoto. 2002a. The period gene and genetic differentiation between three Brazilian populations of Lutzomyia longipalpis. Insect Mol. Biol. 11: 315-323. [ Links ]
Bauzer, L.G., J.S. Gesto, N.A. Souza, R.D. Ward , J.G. Hamilton, C.P. Kyriacou & A.A Peixoto .2002b..Molecular divergence in the period gene between two putative sympatric species of the Lutzomyia longipalpis complex. Mol. Biol. Evol. 19:1624-1627. [ Links ]
Bottecchia, M., S.G. Oliveira , L.G. Bauzer , N.A. Souza, R.D. Ward, K.J. Garner, C.P. Kyriacou & A.A. Peixoto.2004. Genetic divergence in the cacophony IVS6 intron among five Brazilian populations of Lutzomyia longipalpis. J. Mol. Evol. 58: 754-761. [ Links ]
Coyne, J.A. & H.A. Orr. 1989. Patterns of speciation in Drosophila. Evolution 43:362-381. [ Links ]
Coyne, J.A. & H.A. Orr . 1997. Patterns of speciation in Drosophila revisited. Evolution 51:295-303. [ Links ]
Coyne, J .A. & H.A. Orr. 1998. The evolutionary genetics of speciation. Philos. T. Roy. Soc. B. 353:287-305. [ Links ]
Coyne, J. A., S.Y. Kim, A.S. Chang, D. Lachaise, & S. Elwyn. 2002. Sexual isolation between two sibling species with overlapping ranges Drosophila santomea y Drosophila yakuba. Evolution 56: 2424-2434. [ Links ]
de Queiroz Balbino, V., I.V. Coutinho-Abreu , I.V. Sonoda , M.A. Melo, P.P. de Andrade PP, J.M. de Castro JA, J.M. Rebelo, S.M. Carvalho & M. Ramalhoortigao.2006. Genetic structure of natural populations of the sand fly Lutzomyia longipalpis (Diptera: Psychodidae) from the Brazilian northeastern region. Acta Trop. 98: 15-24. [ Links ]
Feliciangeli, M.D., J.C. Arrivillaga, A. Bravo & F. Arias (2004) Activity of Lutzomyia pseudolongipalpis and Lutzomyia longipalpis s.l. (Diptera: Psychodidae) in Venezuela. Parasite 11:273-8. [ Links ]
Gallant, S.l. & D.J. Fairban. 1997. Patterns of postmating reproductive isolation in a newly- discovered species. Aquarius remigis and Aquarius. remigoides (Hemiptera; Gerridae). Heredity 78: 571-577. [ Links ]
Goudet, J. 1999. FSTAT, a program to estimate and test gene diversities and fixation indices (version 2.8). [ Links ]
Hamilton, J., R. Maingon , R. Ward & R . Brazil. 2005. Proceeding of fifth international symposium on Phlebotominae sandflies, Gammarth, Tunis, Tunisia. [ Links ]
Harrison, R.G. 1983. Barriers to gene exchange between closely related cricket species. I. Laboratory hybridization studies. Evolution 37:245-251. [ Links ]
Hodgkinson V.H., J. Birungi, M. Quintana, R. Dietze & L.E. Munstermann .2003. Mitochondrial cytochrome b variation in populations of the visceral leishmaniasis vector Lutzomyia longipalpis across Eastern Brazil. Am. J. Trop. Med. Hyg. 69: 386-392. [ Links ]
Jiggs, C.D. 2006. Speciation: Reinforced butterfly speciation. Heredity 96: 107–108. [ Links ]
Killick-Kendrick, R., A.J. Leaney & P.D Ready. 1977. The establishment, maintenance and productivity of a laboratory colony of Lutzomyia longipalpis (Diptera: Psychodidae). J. Med. Entomol. 13: 429-440. [ Links ]
Kirkpatrick, M., & S. Nuismer. 2004. Sexual selection can constrain sympatric speciation. P. Roy. Soc. B. 271: 687-693. [ Links ]
Lampo, M., D. Torgerson , L.M. Marquez, M. Rinaldi , C.Z. Garcia & A. Arab. 1999. occurrence of sibling species of Lutzomyia longipalpis (Diptera: Psychodidae) in Venezuela: first evidence from reproductively isolated sympatric populations. Am. J. Trop. Med. Hyg. 61: 1004-1009. [ Links ]
Lanzaro, G.C., K. Ostrovska , M.V. Herrero, P.G .Lawyer & A. Warburg.1993. Lutzomyia longipalpis is a species complex: genetic divergence and interspecific hybrid sterility among three populations. Am. J. Trop. Med. Hyg. 48: 839-847. [ Links ]
Lanzaro, G.C., B. Alexander, J.P. Mutebi , J. Montoya-Lerma & A. Warburg .1998. Genetic variation among natural and laboratory colony populations of Lutzomyia longipalpis (Lutz & Neiva, 1912) (Diptera: Psychodidae) from Colombia. Mem. I. Oswaldo Cruz. 93: 65-69. [ Links ]
Lewis P.O. & D. Zaykin.1999. Genetic data analysis: Computer program for the analysis of allelic data, version 1.0 (d12). [ Links ]
Markow, T.A. 1997. Assortative fertilization in Drosophila. P. Natl Acad Sci USA 94:7756-7760. [ Links ]
Mackinnon, J., S. Mori, K. Bhackmar, L. David, D.M. Kingsley, L. Jamreson, J. Chou & D. Schuluter. 2004. Evidence for ecologys role in speciation. Nature 429: 294- 297. [ Links ]
Maingon, R.D., R.D. Ward, J.G. Hamilton, H.A. Noyes, N. Souza , S.J Kemp & P.C Watts. 2003. Genetic identification of two sibling species of Lutzomyia longipalpis (Diptera: Psychodidae) that produce distinct male sex pheromones in Sobral, State, Brazil. Mol. Ecol. 12: 1879-1894. [ Links ]
Morrison, C.A., L.E. Munstermann, C. Ferro , R. Pardo & M. Torres. 1995. Ecological and genetic studies of Lutzomyia longipalpis in a central Colombian focus of visceral leishmaniasis. Bol. Dir. Malar San. Amb.35: 235-248. [ Links ]
Moehring, A, A. Llopart , S. Elwyn, J. Coyne & T. Mackay. 2006. The genetics basis of prezygotic reproductive isolation between Drosophila santomea and Drosophila yakuba due to mating preference. Genetics 173: 215-223. [ Links ]
Mukhopadhyay, J., E.F. Rangel, K. Ghosh & L. Munstermann. 1997. Patterns of genetic variability in colonized strains of Lutzomyia longipalpis (Diptera: Psychodidae) and its consequences. Am. J. Trop. Med. Hyg. 57: 216-221. [ Links ]
Mukhopadhyay., J., K. Ghosh, A.C. Azevedo , E.F. Rangel & L.E. Munsterman. 1998a. Genetic polymorphism of morphological and biochemical characters in a Natal, Brazil, population of Lutzomyia longipalpis (Diptera: Psychodidae). J. Am. Mosq. Control Assoc. 14: 277-282. [ Links ]
Mukhopadhyay, J., K. Ghosh, E.F. Rangel & L.E. Munsterman.1998b. Genetic variability in biochemical characters of Brazilian field populations of the Leishmania vector, Lutzomyia longipalpis (Diptera: Psychodidae). Am. J. Trop. Med. Hyg. 59: 893-901. [ Links ]
Mutebi, J.P., B. Alexander, I. Sherlock , J. Wellington, A.A. Souza , J. Shaw , E.F. Rangel & G.C. Lanzaro.1999. Breeding structure of the sand fly Lutzomyia longipalpis (Lutz & Neiva) in Brazil. Am. J. Trop. Med. Hyg. 61: 149-157. [ Links ]
Mutebi, J.P., F. Tripet , J.B. Alexander & G.C. Lanzaro. 2002. Genetic differentiation among populations of Lutzomyia longipalpis (Diptera: Psychodidae) in Central and South America. Ann. Entomol. Soc. Am. 95: 740:752. [ Links ]
Naisbit, R., C.D. Jiggins & J. Mallet. 2001. Disruptive sexual selection against hybrids contributes to speciation
between Heliconius cydo and Heliconius melpomene. P. Roy. Soc. B. 268: 1849-1854.
Navajas, M., A. Tsagkarakow, J. Lagnel & M. Perrot-Minnot. 2000. Genetic differentiation in the Tetranychusnychus urticae (Acari; Tetranychidae) polymorphims host races o sibling species?. Exp. Appl. Acarol. 24: 365-376. [ Links ]
Oliver, C.G. 1978. Experimental hybridization between the nymphalid Butterflies. Phyclodes tharos and campestris montana. Evolution 32: 594- 601. [ Links ]
Orr, H.A. 1995. The populations genetics of speciation: the evolution of hybrid incompatibilities. Genetics 139: 1805- 1813. [ Links ]
Orr, H.A. & J.A .Coyne .1989. The genetic of postzygotic isolation in the Drosophila virilis group. Genetics 121: 527-537. [ Links ]
Panhuis, T., R. Butlin, M. Zuk & T. Trengenza. 2001. Sexual selection and speciation. Trends Ecol. Evol. 16: 364-371. [ Links ]
Presgraves, D.C .2002.. Patterns of postzygotic isolation in Lepidoptera. Evolution 56:1168-1183. [ Links ]
Presgraves, D.C .& H.A. Orr (1998) Haldane rule in taxa lacking hemizygous. Science 282: 952- 954. [ Links ]
Price, C.S.C., C.H. Kim, J. Posluszny & J.A. Coyne (2000) Mechanims of conspecific sperm precedence in Drosophila. Evolution 54: 2028-2037. [ Links ]
Price, C.S.C., C.H. Kim, C.J. Gronlund & J.A. Coyne. 2001. Cryptic reproductive isolation in the Drosophila simulans species complex. Evolution 55: 81- 92. [ Links ]
Reed, L. & T. Markow. 2004. Early events in the speciation polymorphism of hybrid male sterility in Drosophila. P. Natl. Acad. Sci. USA. 101: 9009-9012. [ Links ]
Slotman, M., A. della Torre & J.R. Powell. 2004. The genetics of inviability and male sterility in hybrids between Anopheles gambiae and Anopheles arabiensis. Genetics 167: 275-287. [ Links ]
Soto, S.I., T. Lehmann, E.D. Rowton, B.I.D. Velez & C.H. Porter. 2001. Speciation and population structure in the morphospecies Lutzomyia longipalpis (Lutz & Neiva) as derived from the mitochondrial ND4 gene. Mol. Phylogen. Evol. 18: 84-93. [ Links ]
Souza, N.A., F.M. Vigoder, A.S. Araki, R.D. Ward, C.P. Kyriacou, H. Yin, J..P Mutebi, S. Marriot & G.C. Lanzaro. 1999. Metaphase karyotypes and G-banding in sandflies of the Lutzomyia longipalpis complex. Med. Vet. Entomol.13: 72-77. [ Links ]
Turelli, M. & H.A. Orr .1995. The dominance theory of Haldanes rule. Genetics 140: 389-402. [ Links ]Turelli, M. & L. Oyle. 2007. Assymetric postmating isolation Darwins corollary theory haldanes rule. Genetics 176: 1059-1088. [ Links ]
Van Doorn, G. S. & F.J. Weissing .2001. Ecological versus sexual selection models of sympatric speciation. A synthesis. Selection 2: 17-40. [ Links ]
Via, S. 2002. The ecological genetics of speciation. Am. Nat. 159:1-7. [ Links ]
Velthuis, B.J., W. Yang ,T. van Opijnen & J.H. Werren. 2005. Genetics of female mate discrimination of heterospecific males Nasonia (Hymenoptera, Pteromalidae). J. Anim. Behav. 69: 1.107-1.120. [ Links ]
Warburg, A., E. Saraiva, G.C.Lanzaro, R.G. Titus & F. Neva. 1994. Saliva of Lutzomyia longipalpis sibling species differs in its composition and capacity to enhance leishmaniasis. Philos. T . Roy. Soc. B. 29: 223-230. [ Links ]
Ward, R.D., A. Phillips, B. Burnet & C.B. Marcondes. 1988. The Lutzomyia longipalpis complex: reproduction and distribution. In M. W Service (eds). Biosystematics of Haematophagous insects, Oxford University, Oxford, UK. p. 258-269. [ Links ]
Ward, R.D, A.L. Ribeiro, P.R. Ready & A. Murtagh. 1983. Reproductive isolation between different forms of Lutzomyia longipalpis (Lutz & Neiva), (Diptera: Psychodidae), the vector of Leishmania donovani chagasi Cunha & Chagas and its significance to kalaazar distribution in South America. Mem.I. Oswaldo Cruz 78: 269-280. [ Links ]
Watts, P.C., J.C.G. Hamilton, R.D. Ward, H.A. Noyes, N.A. Souza , S.J. Kemp, M.D. Feliciangeli, R. Brazil & R.D.C. Maingon. 2005. Male sex pheromones and the phylogeographic structure of the Lutzomyia longipalpis species complex (Diptera: Psychodidae) from Brazil and Venezuela. Am. J. Trop. Med. Hyg. 73: 734-743. [ Links ]
Willet, C.S. & R.S. Burton. 2001. Viability of cytocrome C genotypes depends on cytoplamic backgrounds in Trigriopus californicus. Evolution 55: 1592- 1599. [ Links ]
Yin, H., D.E. Norris & G.C. Lanzaro. 2000. Sibling species in the Lutzomyia longipalpis complex differ in levels of mRNA expression for the salivary peptide, maxadilan. Insect Mol. Biol. 9: 309–314. [ Links ]
Young, D.G., M.A. Duncan. 1994. Guide to the identification and geographic distribution of Lutzomyia sand flies in Mexico, the West Indies, Central and South America (Diptera: Psychodidae). Associated Publishers, Gainesville, Florida, EEUU. [ Links ]

