










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.4 San José Dec. 2008
Papilionoidea de la Sierra de Huautla, Morelos y Puebla, México (Insecta: Lepidoptera)
Mercedes Luna-Reyes1, Jorge Llorente-Bousquets2* & Armando Luis-Martínez2
- Museo de Zoología, Facultad de Estudios Superiores Zaragoza, Av. Guelatao No. 66. Col. Ejército de Oriente. Iztapalapa. México D.F. 09230; mmluna@puma2.zaragoza.unam.mx, alm@hp.fciencias.unam.mx
- Museo de Zoología, Facultad de Ciencias, UNAM. Apartado Postal 70-399, México D.F. 04510. *Responsable de la publicación; jlb@ hp.fciencias.unam.mx
Abstract: Papilionoidea from Sierra de Huautla, Morelos and Puebla, México (Insecta: Lepidoptera).
The Cuenca del Balsas region has significant biodiversity and endemicity of its herpetofauna, avifauna and vascular plants. Despite this, our knowledge of the Papilionoidea of the region is poor. We analyzed the local and temporal distribution of Papilionoidea at 24 localities in the states of Morelos and Puebla. The study sites are situated between 900 and 1300 m. a. s. l., and are composed of dry tropical forest (dtf). We recorded 8790 individuals of 83 genera and 142 species of Papilionoidea (sensu Kristensen, 1975), over 79 days of field work, with 2-4 days at each of the 24 localities. Twenty five species were newly recorded for the state of Puebla. Our data render Morelos and Puebla among the seven richest Mexican states, in terms of Papilionoidea diversity. Our results show that the Sierra de Huautla has the lowest diversity, but the highest standard abundance, compared to other Mexican regions with similar vegetation. Patterns of diversity and seasonal abundance are atypical, in that individuals of many species are unusually abundant during the wet months. Rev. Biol. Trop. 56 (4): 16771716. Epub 2008 December 12.
Key words: Cuenca del Balsas, dry tropical forest, phenology, richness, tropical forest.
En los últimos años se ha aplicado el término megadiverso para aquellos países que tienen representados en su superficie valores extraordinarios de la biodiversidad mundial; este reducido grupo solo incluye 12 países, México ocupa el séptimo lugar, por su gran riqueza biológica y por su alta proporción de endemismos (Mittermeier y Goettsch 1992, CONABIO 2002). Rzedowski (1992) estimó que en el territorio mexicano existen 22,800 especies de plantas vasculares, que representan alrededor del 10% de las registradas a nivel de mundial (Flores y Gerez 1994). En cuanto a la fauna, el análisis de la riqueza muestra que en México habita el 10% de los vertebrados del mundo, aunque en los anfibios está representado alrededor del 7% faltan muchos por descubrir y describir. Los valores de endemismo también resultan altos, oscilando entre 10% en aves y 55.7% en herpetofauna (Espinosa 1993, Flores 1993, Navarro y Benítez 1993, Arita y León 1993, Fa y Morales 1993).
Los insectos incluyen alrededor del 60-65% de la biota descrita (Morón y valenzuela 1993, Llorente et al. 1996, Luis et al. 2000). El orden Lepidoptera se considera uno de los grupos hiperdiversos (Martín-Piera y Lobo 2000), con 146,000 especies, agrupadas en 27 superfamilias; una de las cuales es Papilionoidea que representa el 13.1% especies descritas lepidópteros en todo el mundo (Llorente et al. 1996, Lamas 2000). De acuerdo con los resultados por Luis et al. (2003b), del total de 1016 especies de papilionoideos (sensu Kristensen 1975) reconocidas para México, el 8.66% es endémico. Además de la importancia que tiene por su riqueza, Papilionoidea ha sido ampliamente utilizado como modelo en trabajos sobre análisis de la biodiversidad, en impacto ambiental y bioconservación, así como en estudios sistemáticos, biogeográficos, genéticos, etológicos y ecológicos; muchas de sus especies han sido consideradas por los especialistas como indicadoras del estado de los hábitats y su riqueza (Beccaloni y Gaston 1995, Llorente et al. 1996, Brown K. 1997).
Una de las áreas bióticas poco estudiadas, que a la vez coincide con perturbaciones extensas, corresponde al tipo de vegetación de selvas caducifolias (Trejo y Dirzo 2000). Esta comunidad vegetal predomina en la cuenca del Balsas, en el estado de Morelos y sur del estado de Puebla. La sierra de Huautla forma parte de la Cuenca en estos estados, región donde habita gran diversificación biológica, concentrándose en ella parte de las especies endémicas de fanerógamas (Flores y Gerez 1994) y valores altos de riqueza y especies endémicas para la herpetofauna (Flores 1993) y aves mexicanas (Escalante et al. 1993).
Este estudio tiene el propósito de contribuir al conocimiento de la diversidad de los Papilionoidea de la cuenca oriental del Balsas y, en particular, de la sierra de Huautla y áreas adyacentes en Morelos y Puebla. Además del reconocimiento de la riqueza del área, se describe la distribución regional y la estacionalidad de la comunidad.
Antecedentes: De acuerdo con la información registrada en la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), existen más de 500 mil registros de ejemplares de mariposas recolectadas en México. A pesar de esto, el mapa de las localidades de recolecta del Atlas de Mariposas de México, desarrollado actualmente por los investigadores del Museo de Zoología de la Facultad de Ciencias de la UNAM (Llorente et al. 1997, Luis et al. 2003b), claramente indica que la lepidopterofauna de las selvas bajas caducifolias de México está poco estudiada y que, en particular, en la sierra de Huautla solo se habían registrado cinco especies distribuidas en tres localidades. Por ello es que resulta necesario el desarrollo de estudios que permitan conocer la composición faunística de las comunidades de mariposas asociadas con este tipo de vegetación.
Un estudio equivalente al nuestro en áreas próximas corresponde al publicado por De la Maza (1975); otros desarrollados por este autor se efectuaron con fines biogeográficos en áreas de menor altitud, que también incluyen otros tipos de vegetación (De la Maza et al. 1995a,b).
Descripción del Área de Estudio: La sierra de Huautla en conjunto con la sierra de Taxco, actualmente se considera como una de la Regiones Terrestres Prioritarias para la conservación en México (RTP-120). Esta región denominada como sierras de Taxco-Huautla, se ubica entre las coordenadas 18°1832"18°5221" N, 98°4849"-100°0900" W, e incluye parte de los estados de México, Guerrero, Morelos y Puebla, ocupando una superficie de 2959 km2 (CONABIO 2000).
Geología y Edafología: La sierra de Huautla fundamentalmente está constituida por rocas ígneas extrusivas del Terciario, sobre las que se han desarrollado asociaciones de suelos someros (10 a 50 cm de profundidad) de tipo feozem háplico más regosoles y vertisoles.
Fisiografía: La sierra de Huautla está en un límite de la provincia fisiográfica del Eje Neovolcánico y presenta características geológicas heterogéneas debido a su proximidad con la Sierra Madre del Sur, con la que comparte varios rasgos. Se ubica dentro de la Subprovincia de las sierras abruptas del sur de Puebla e incluye áreas de los estados de Puebla y Morelos. Esta sierra es adyacente a la cuenca alta del Balsas.
Hidrografía: El área está drenada por diferentes ríos afluentes del río Balsas, como el Amacuzac en el extremo occidental y el Tepalcingo, el Nexapa y el Huehuetlán hacia la parte oriental. Todos ellos nutridos por multitud de arroyos.
Clima y vegetación: En la sierra de Taxco-Huautla se presentan cinco subtipos de clima de acuerdo con la clasificación de Köppen modificada por García (1981). La mayor parte corresponde a (A)C(w2), que es un clima semicálido y subhúmedo, con lluvias en verano; después y casi en la misma proporción Aw0 y Aw1 que son climas cálidos-subhúmedos con lluvias en verano; (A)C(w1), clima semicálido y subhúmedo, con lluvias en verano y C(w2)x, clima templado al que le corresponde la menor parte (CONABIO 2000).
En el área de estudio predomina el clima de tipo cálido subhúmedo con régimen de lluvias de verano, con poca oscilación térmica y marcha de temperatura tipo ganges Aw0(w) (i)g, donde se desarrolla la selva baja caducifolia que se presenta con intensidades de perturbación diferentes, en asociación con porciones transformadas en pastizales inducidos y áreas de cultivo (INEGI 1981). Aunque existen áreas cubiertas por bosques de encino, pino, pino-encino, mesófilo y zacatonal alpino, y otras en las que se realizan actividades agrícolas y pecuarias, para la sierra Taxco-Huautla la vegetación predominante (41%) es la selva baja caducifolia (CONABIO 2000).
Material y métodos
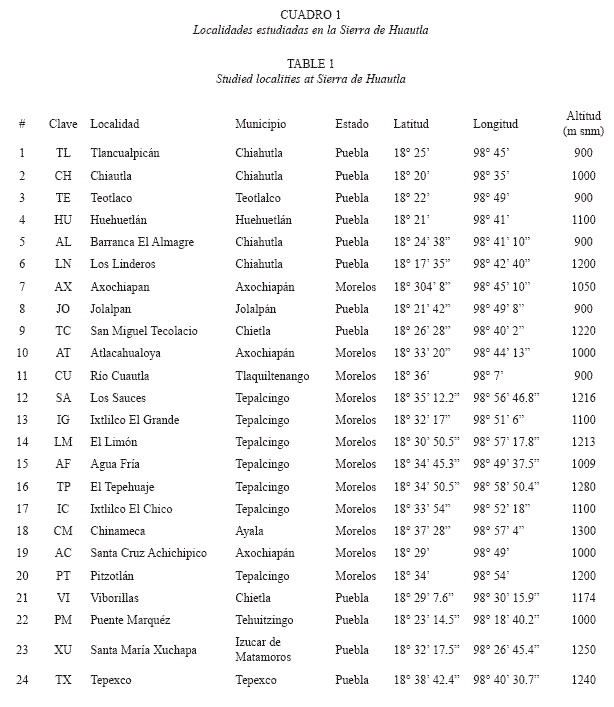
Recolección de ejemplares: De febrero de 1997 hasta julio de 2000, se efectuaron 79 visitas diarias al área de estudio, repartidas a lo largo de 25 meses con un promedio de dos a cuatro días por mes, muestreando así en 24 localidades de selva baja caducifolia ubicadas en un intervalo altitudinal entre los 900 y 1 300 m, en los estados de Morelos y Puebla. En el Cuadro 1 se presenta la relación de los sitios elegidos, el municipio y estado al que pertenecen, así como los datos de la georreferenciación y altitud obtenidos con un GPS marca Garmin modelo12 XL.
La recolecta de individuos se llevó a cabo mediante el uso de redes entomológicas aéreas. En cada sitio se efectuó un recorrido a lo largo de un transecto de 2 km, explorando entre la vegetación, en las laderas, paredes y oquedades de las elevaciones, en el lecho de arroyos o ríos, o sobre el suelo húmedo de las orillas de charcos, arroyos o ríos, o sobre su lecho seco, en veredas y caminos y en cualquier microhábitat donde se observaran mariposas, siguiendo las técnicas recomendadas por Howe (1975), Pollard (1977) y Clench (1979). El tiempo de permanencia en cada localidad fue de cuatro horas en el período comprendido de las 0800 a las 1900 aproximadamente. Los ejemplares fueron capturados al vuelo, sobre el sustrato alimentario o sobre arena húmeda mediante la red aérea indicada; las recolectas se efectuaron por al menos dos personas, aunque en algunos sitios participaron seis con diversas habilidades.
Cada mariposa capturada fue sacrificada y colocada de manera individual en una bolsa de papel "glassine", en la que se anotaron los datos de localidad, fecha, hora y nombre del colector.
Procesamiento del material entomológico: Se preparó una muestra de ejemplares de cada localidad y especie, de acuerdo con las técnicas convencionales propuestas por Howe (1975) para el montaje de ejemplares. Así mismo, el material fue rotulado incluyendo los datos nomenclaturales (especie o subespecie, según el caso), de recolecta (localidad, fecha, coordenadas geográficas, altitud, hora, tipo de vegetación y nombre del colector) y el número de catálogo y el de recolecta que identifica a cada ejemplar. Finalmente, el material fue incorporado a la colección lepidopterológica del Museo de Zoología de la F.E.S. Zaragoza, UNAM.
Determinación taxonómica: Todos los ejemplares fueron mantenidos en cajas con cristales de paradiclorobenceno durante tres semanas. Después se procedió a su determinación taxonómica, para lo cual se consultaron obras especializadas e ilustradas como las de Ehrlich y Ehrlich (1961), Emmel (1975), Pyle y Knopf (1981), Scott (1986), De la Maza (1987), Devries (1987), Smart (1989), Llorente et al. (1997) y Luis et al. (2003a), así como la colección lepidopterológica del Museo de Zoología de la F.E.S. Zaragoza (MZFZ) y la colección del Museo de Zoología de la Facultad de Ciencias (MZFC), ambas de la UNAM.
Registro de ejemplares: Con los datos registrados se elaboró un catálogo escrito que después fue transferido a una base de datos electrónica en el modelo Biótica (versión 3.1), a partir de la cual se procesaron los registros con los cuales se elaboraron cuadros y figuras utilizados en el análisis de distribución regional y temporal de la riqueza, así como la abundancia de los papilionoideos del área estudiada.
Lista de especies: Se integró la lista de especies de papilionoideos que fueron registrados en la totalidad del área de estudio. Este inventario se presenta siguiendo la nomenclatura y el orden filogenético propuesto por Kristensen (1975) para la superfamilia Papilionoidea, y a Tyler et al. (1994) para Papilionidae; para las demás familias (Pieridae, Nymphalidae y Lycaenidae) el orden propuesto por Scott (1985), Luis et al. (1991, 2003a), vargas et al. (1992, 1999) y Llorente et al. (1997).
Esfuerzo de captura: Para cada sitio se elaboraron curvas de acumulación de especies donde se ilustran los valores observados de la riqueza específica y los estimados mediante el índice de Chao1 (Colwell y Coddington 1994). Los registros de especies por sitio se anexaron a una matriz de incidencia (presencia-ausencia) de tipo r x c, donde los renglones son especies y las columnas son localidades de recolecta. También se trazó la curva de acumulación de las especies registradas y se aplicó el índice de Chao2 (Colwell y Coddington 1994, León-Cortés et al. 1998), indicador que se utiliza en estudios que consideran pocos muestreos o que se realizan en períodos cortos y donde existe gran cantidad de especies raras; con él se puede estimar la riqueza teórica de una región requiriendo únicamente los datos de presenciaausencia de las especies (Feria 1997).
Con propósitos de comparación también se aplicaron los métodos sugeridos por Soberón y Llorente (1993) e instrumentados por Díaz-Francés y Soberón (2005).
Riqueza y abundancia: Se analizó la riqueza y la abundancia total, así como su distribución espacial y temporal en el área de estudio. Para ello la lista de especies fue contrastada con la obtenida en estudios realizados por González (1996) en Puebla, y Luna-Reyes (2007) y Luna-Reyes et al. (en prep.) en Morelos, en los que se llevó a cabo una revisión exhaustiva de la colección del MZFZ y del MZFC con su megabase de datos MARIPOSA y de la literatura especializada sobre las especies citadas para estas áreas. También se consultaron los trabajos de De la Maza (1975), Lara (1999), valencia (1999) y vences (2004), así como las obras : Catálogo Sistemático y Zoogeográfico de los Lepidópteros Mexicanos (Hoffmann 1940), Mariposas de México (Beutelspacher 1984), Mariposas Mexicanas (De la Maza 1987), Papilionidae y Pieridae de México: Distribución geográfica e Ilustración (Llorente et al. 1997) y Nymphalidae de México I (Danaine, Apaturinae, Biblidinae y Heliconiinae): Distribución geográfica e Ilustración (Luis et al. 2003a).
La riqueza total del área de estudio se comparó con la encontrada en trabajos metodológicamente similares a éste, en los que se reconoció la diversidad de Papilionoidea de regiones amplias en las que se incluyeron varias localidades, como la sierra de Juárez, Oaxaca (Luis et al. 1991), la sierra de Atoyac de Álvarez, Guerrero (vargas et al. 1992) y la sierra de Manantlán, Colima y Jalisco (vargas et al. 1999). También se establecieron comparaciones con trabajos efectuados en áreas con vegetación tropical (selva baja caducifolia o subcaducifolia) ubicadas en Morelos o en otros estados del país, como Pedernales (Balcázar 1988) y Chorros del varal (Arteaga 1991) en Michoacán, Zenzontla y Ahuacapán en Jalisco (vargas et al. 1999) y Cañón de Lobos en Morelos (Luna-Reyes 2007).
Puesto que el esfuerzo de captura (horas/ día/persona) efectuado fue diferente en las distintas localidades de la sierra de Huautla, en el análisis de abundancia se consideraron los datos de la abundancia estandarizada (abundancia/esfuerzo de captura). Se siguió el mismo tratamiento para los datos de los sitios con los que se comparó este trabajo y en los que también se registró con detalle la abundancia y el esfuerzo de captura empleado: Agua Dulce y Platanarillos en Colima, Ahuacapán, La Calera, Los Mazos y Zenzontla en Jalisco (vargas et al. 1999), y Cañón de Lobos en Morelos (Luna-Reyes 2007).
Abundancia por especie: Se contabilizó el número de especímenes directamente y por categorías de abundancia. Las especies fueron ordenadas de acuerdo con su abundancia en seis categorías siguiendo la escala geométrica en agrupamientos de tamaño por cuatro: MR (Muy Rara: especies con un ejemplar), R (Rara: dos a cinco ejemplares), E (Escasa: seis a 21), F (Frecuente: 22 a 85), C (Común: 86 a 331) y MC (Muy Común: especies con 332 ejemplares o más), de acuerdo con las propuestas de Lamas (1984), Krebs (1985) y Luis y Llorente (1990).
Fenología: Todos los datos de riqueza y abundancia del área de estudio fueron agrupados por mes y, como en los análisis anteriores, la abundancia fue estandarizada. Estos resultados fueron comparados con los trabajos de Arteaga (1991), vargas et al. (1992, 1999) y Luna-Reyes (2007).
Resultados
Lista sistemática y riqueza: Se registraron 142 especies, 83 géneros y cuatro familias de Papilionoidea, con base en la determinación taxonómica de los 8790 especímenes recolectados en 24 localidades de la sierra de Huautla.
Papilionoidea de la sierra de Huatla
Papiloidae
Baroniinae
Baronia
- B. brevicornis brevicornis Salvin, 1893
- P. epidaus fenochionis (Salvin & Godman, 1868)
- B. philenor philenor (Linnaeus, 1771)
- B. polydamas polydamas (Linnaeus, 1758)
Papilioninae
Protographium
Battus
Parides
5. P. erithalion trichopus (Rothschild & Jordan, 1906)
6. P. montezuma (Westwood, 1842)
7. P. photinus (Doubleday, 1844)
Heraclides
8. H. cresphontes (Cramer, 1777)
9. H. ornythion ornythion (Boisduval, 1836)
10. H. rogeri pharnaces (Doubleday, 1846)
11. H. thoas autocles (Rothschild & Jordan, 1906)
Papilio
12. P. polyxenes asterius Stoll, 1782 Pterourus
13. P. multicaudata multicaudata (W. F. Kirby, 1884)
Pieridae
Coliadinae
Zerene
14. Z. cesonia cesonia (Stoll, 1791)
Anteos
15. A. maerula (Fabricius, 1775)
Phoebis
16. P. agarithe agarithe (Boisduval, 1836)
17. P. argante ssp. n.
18. P. philea philea (Linnaeus, 1763)
19. P. sennae marcellina (Cramer, 1777)
Aphrissa
20. A. statira statira (Cramer, 1777)
Abaeis
21. A. nicippe (Cramer, 1779)
Pyrisitia
22. P. dina westwoodi (Boisduval, 1836)
23. P. lisa centralis (Herrich-Schäffer, 1865)
24. P. nise nelphe (R. Felder, 1869)
25. P. proterpia (Fabricius, 1775)
Eurema
26. E. arbela boisduvaliana (C. Felder & R. Felder, 1865)
27. E. daira sidonia (R. Felder, 1869)
28. E. mexicana mexicana (Boisduval, 1836)
29. E. salome jamapa (Reakirt, 1866)
Nathalis
30. N. iole Boisduval, 1836
Kricogonia
31. K. lyside (Godart, 1819)
Pierinae
Hesperocharis
32. H. costaricensis pasion (Reakirt, [1867])
Glutophrissa
33. G. drusilla tenuis (Lamas, 1981)
Leptophobia
34. L. aripa elodia (Boisduval, 1836)
Ascia
35. A. monuste monuste (Linnaeus, 1764)
Ganyra
36. G. josephina josepha (Salvin & Godman, 1868 )
Nymphalidae
Libytheinae
Libytheana
37. L. carinenta mexicana Michener, 1943
Danainae
Danaus
38. D. eresimus montezuma Talbot, 1943
39. D. gilippus thersippus (W. H. Bates, 1863)
40. D. plexippus plexippus (Linnaeus, 1758)
Morphiinae
Morpho
41. M. polyphemus polyphemus Westwood, 1851
Satyrinae
Cissia
42. C. pompilia (C.Felder & R. Felder, 1867)
43. C. similis (Butler, 1867)
44. C. themis (Butler, 1867)
Euptychia
45. E. fetna Butler, 1870
Hermeuptychia
46. H. hermes (Fabricius, 1775)
Megisto
47. M. rubricata rubricata (W. H. Edwards, 1871)
Pindis
48. P. squamistriga R. Felder, 1869
Taygetis
49. T. weymeri Draudt, 1912
Charaxinae
Anaea
50. A. troglodyta aidea (Guérin-Ménéville, [1844])
Apaturinae
Asterocampa
51. A. idyja argus (H. W. Bates, 1864)
Doxocopa
52. D. laure laure (Drury, 1773)
Nymphalinae
Vanessa
53. V. annabella (Field, 1971)
Smyrna
54. S. blomfildia datis Frühstorfer, 1908
Anartia
55. A. amathea fatima Fabricius, 1793
56. A. jatrophae luteipicta Frühstorfer, 1907
Junonia
57. J. coenia Hübner, [1822]
58. J. evarete nigrosuffusa Barnes & McDunnough, 1916
Siproeta
59. S. stelenes biplagiata (Frühstorfer, 1907)
Anthanassa
60. A. nebulosa alexon (Godman & Salvin, 1889)
61. A. ptolyca amator (A. Hall, 1929)
62. A. texana texana (W. H. Edwards, 1863)
Chlosyne
63. C. hippodrome hippodrome (Geyer, 1837)
64. C. janais janais (Drury, 1782)
65. C. lacinia lacinia (Geyer, 1837)
66. C. rosita riobalsensis Bauer, 1961
67. C. ehrenbergii (Geyer, [1833])
Microtia
68. M. elva elva H. W. Bates, 1864
Phyciodes
69. P. mylitta thebais Godman y Salvin, 1878
70. P. pallescens (R. Felder, 1869)
71. P. pulchella pulchella (Boisduval, 1852)
Tegosa
72. T. frisia tulcis (H.W. Bates, 1864)
Texola
73. T. anomalus coracara (Dyar, 1912)
74. T. elada elada (Hewitson, 1868)
Mestra
75. M. dorcas amymone (Ménétriés, 1857)
Eunica
76. E. monima (Stoll, 1782
Myscelia
77. M. cyananthe cyananthe C. & R. Felder, 1867
Hamadryas
78. H. amphinome mexicana (Lucas, 1853)
79. H. atlantis lelaps (Godman & Salvin, 1883)
80. H. februa ferentina (Godart, [1824])
81. H. glauconome glauconome (W. H. Bates, 1864)
82. H. guatemalena marmarice (Frühstorfer, 1916)
Bolboneura
83. B. sylphis beatrix R. G. Maza, 1985
Cyclogramma
84. C. bacchis (Doubleday, 1849)
Adelpha
85. A. iphicleola iphicleola (H.W. Bates, 1864)
Marpesia
86. M. chiron marius (Cramer, 1779)
87. M. petreus ssp. n.
Heliconiinae
Euptoieta
88. E. claudia daunius (Herbst, 1798)
89. E. hegesia meridiania Stichel, 1938
Agraulis
90. A. vanillae incarnata (Riley, 1926)
Dione
91. D. juno huascuma (Reakirt, 1866)
92. D. moneta poeyii Butler, 1873
Dryas
93. D. iulia moderata (Riley, 1926)
Heliconius
94. H. charithonia vazquezae W. P. Comstock y F. M. Brown, 1950
Lycaenidae
Theclinae
Cyanophrys
95. C. goodsoni Clench, 1946
Rekoa
96. R. palegon (Cramer, 1780)
97. R. zebina (Hewitson, 1869)
Arawacus
98. A. jada (Hewitson, 1867)
Chlorostrymon
99. C. simaethis (Drury, 1770)
100. C. telea (Hewitson, 1868)
Allosmaitia
101. A. strophius (Godart, [1824])
Calycopis
102. C. isobeon (Butler & H. Druce, 1872)
Electrostrymon
103. E. sangala (Hewitson, 1868)
Strymon
104. S. rufofusca (Hewitson, 1877)
105. S. yojoa (Reakirt, [1867])
106. S. cestri (Reakirt, [1867])
107. S. albata (C. Felder & R. Felder, 1865)
108. S. bebrycia (Hewitson, 1868)
109. S. istapa (Reakirt, [1867])
110. S. bazochii (Godart, [1824])
Ministrymon
111. M. clytie (W. H. Edwards, 1877)
112. M. azia (Hewitson, 1873)
Panthiades
113. P. bitias (Cramer, 1777)
114. P. bathildis (C. Felder & R. Felder, 1865)
115. "Thecla" sp.
Polyommatinae
Leptotes
116. L. cassius cassidula (Boisduval, 1870)
117. L. marina (Reakirt, 1868)
Zizula
118. Z. cyna (W. H. Edwards, 1881)
Everes
119. E. comyntas (Godart, [1824])
Hemiargus
120. H. hanno antibubastus Hübner, [1808]
Echinargus
121. E. isola (Reakirt, [1867])
Riodininae
Calephelis
122. C. nemesis nemesis (W. H. Edwards, 1871)
123. C. perditalis perditalis Barnes & McDunnough, 1918
124. C. rawsoni McAlpine, 1939
125. Calephelis sp.
Caria
126. C. stillaticia Dyar, 1912
Baeotis
127. B. zonata zonata R. Felder, 1869
Lasaia
128. L. sessilis Schaus, 1890
129. L. maria maria Clench, 1972
Melanis
130. M. cephise acroleuca R. Felder, 1869
Emesis
131. E. mandana furor Butler & H. Druce, 1872
132. E. vulpina Godman y Salvin, 1886
133. E. poeas Godman, 1901
134. E. tenedia C. Felder & R. Felder, 1861
135. E. lupina lupina Godman & Salvin, 1886
136. E. zela cleis (W. H. Edwards, 1882)
137. E. emesia (Hewitson, 1867)
Apodemia
138. A. palmeri australis Austin, 1988
139. A. walkeri Godman y Salvin, 1886
Thisbe
140. T. lycorias lycorias (Hewitson, [1853])
Juditha
141. J. molpe molpe (Hübner, [1808])
Theope
142. T. eupolis Schaus, 1890
Veinticinco especies son registros nuevos para Puebla (ver Apéndice I), de tal forma que con este trabajo, el número total de especies para Puebla es de 432. Por el contrario, para Morelos todas las especies que integran esta lista se encuentran entre las 355 del catálogo de Papilionoidea que se ha desarrollado desde hace más de 15 años para este estado (Luna-Reyes 2007, Luna-Reyes et al., en prep.).
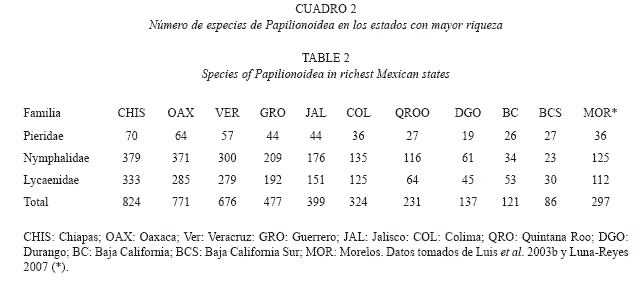
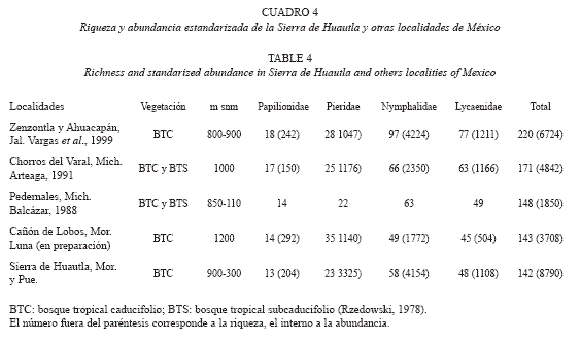
De acuerdo con Luis et al. (2003b), las entidades en las que se ha detectado la mayor riqueza de Papilionoidea en nuestro país son: Chiapas (824 especies), Oaxaca (771), veracruz (676), Guerrero (477), Jalisco (399), en esta jerarquización los estados de Puebla (432) y Morelos (355) ocupan el quinto y séptimo lugar, respectivamente (Cuadro 2).
En la sierra de Huautla se presentan 18 de los taxones endémicos a México (Luis et al. 2000, 2003b): Baronia b. brevicornis, P. epidaus fenochionis, P. erithalion trichopus, E. fetna, T. weymeri, A. nebulosa alexon, A. ptolyca amator, C. rosita riobalsensis, C. ehrenbergii, P. pallescens, T. anomalus coracara, T. e. elada, H. atlantis lelaps, H. guatemalena marmarice, B. sylphis beatrix y M. cephise acroleuca, E. poeas y A. palmeri australis (Apéndice I).
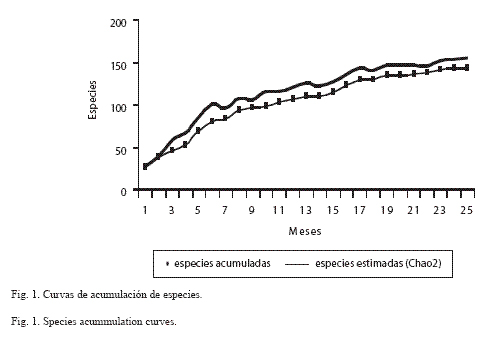
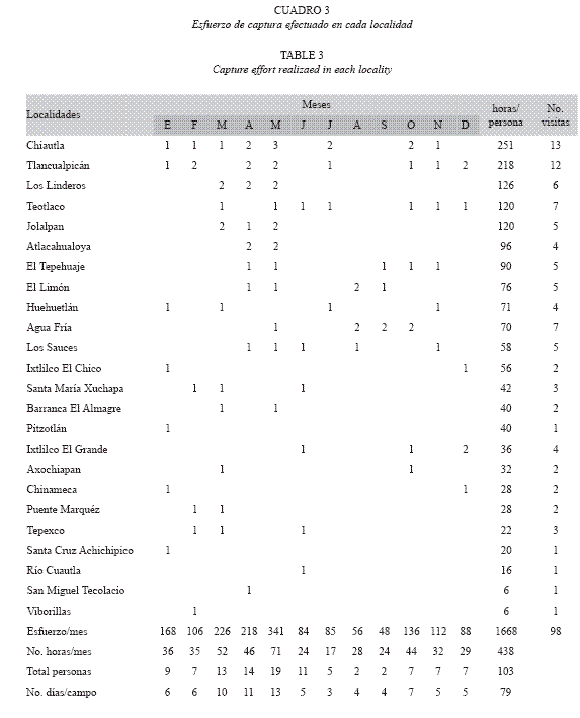
Esfuerzo de captura: El modelo asintótico de Chao2 (Colwell y Coddington 1994, León-Cortés et al. 1998) indicó que la riqueza potencial en la Sierra de Huautla es de 154 especies; en la Fig. 1 se ilustra esta tendencia (Chao2). El esfuerzo de captura total efectuado en la sierra de Huautla fue de 1668 horas/ persona.
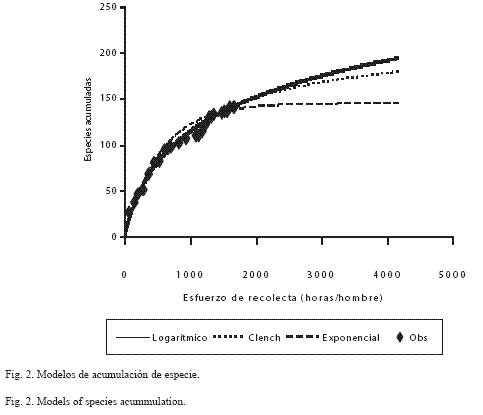
El tiempo de permanencia en cada sitio fue de cuatro horas en cada visita, pero el número de días de recolecta y la cantidad de recolectores (entre dos y seis personas) fueron variables; por ejemplo, en las localidades de Chiautla, Tlancualpicán, Los Linderos, Teotlalco y Jolalpan se efectuó el mayor esfuerzo de captura con más de 100 horas/persona (Cuadro 3). En este cuadro también se aprecia que al utilizar otros estimadores se obtuvieron resultados diferentes a los encontrados con Chao2 (Fig. 2): 194 especies con el modelo de Clench (1979) y 144 con el exponencial; aunque el logarítmico no calcula un valor para la asíntota, permite observar cuánto falta por recolectar. Cuando se evaluó cada opción de acuerdo con Díaz-Francés y Soberón (2005), se encontró que la curva con mayor éxito fue la logarítmica, después y con poca diferencia la de Clench, y al final la exponencial.
Riqueza por familia: En México existen aproximadamente 1200 especies de Papilionoidea (Luis et al. 2003b) repartidas en cuatro familias, siendo Nymphalidae la de mayor diversidad (43.37%), Lycaenidae (42.32%), Pieridae (8.85%) y Papilionidae con el menor número de especies (5.51%). Estos porcentajes se mantienen más o menos constantes (varían entre 5 y 10%) en todo el país, con excepción de los estados de la península de Baja California, en los que Lycaenidae tiene el mayor porcentaje de riqueza (Cuadro 2). En la Sierra de Huautla se presenta esta misma proporción para Nymphalidae 41% (58 especies), una disminución en Lycaenidae 34% (48) y un aumento considerable en Pieridae 16% (23) y Papilionidae 9% (13 especies) (ver lista sistemática).
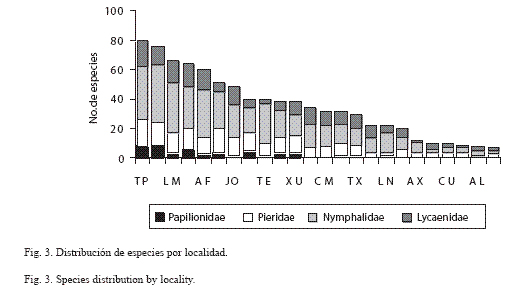
Riqueza y distribución por localidad: El Tepehuaje es el sitio donde se registró la mayor riqueza (80 spp.) y abundancia estandarizada, a pesar de que en función del esfuerzo de captura esta localidad ocupa el séptimo lugar, después y en orden decreciente: Chiautla (75 spp.), El Limón (66 spp.), Tlancualpicán (64 spp.) y Agua Fría (60 spp.) (Cuadro 6; Fig. 3). Chiautla fue el sitio con mayor esfuerzo de recolecta con 251 horas/persona, siguiéndole Tlancualpicán (218 horas/persona), Linderos (218 horas/persona) y Jolalpan y Teotlalco, ambas con 120 horas/persona. Las tres últimas localidades con menor riqueza son viborillas (9 spp., 6 horas/ persona), Barranca El Almagre (8 spp., 40 horas/persona) y San Miguel Tecolacio (7 spp., 6 horas/persona).
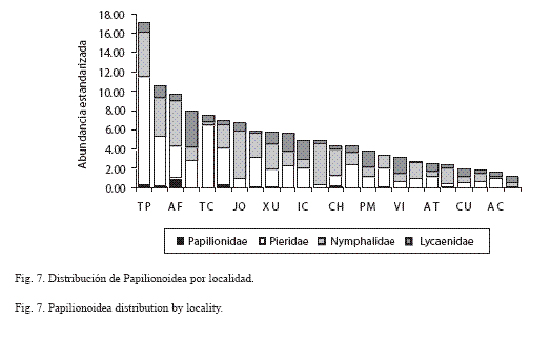
En cuanto a la distribución por familia, en la mayoría de las localidades se observó la predominancia de Nymphalidae y Lycaenidae y la escasa presencia de Papilionidae, incluso su ausencia en Axochiapan, San Miguel Tecolacio, Atlacahualoya, Río Cuautla, Ixtlilco El Chico, Chinameca, Santa Cruz Achichipico, viborillas, Puente Marquéz y Santa María Xuchapa (Fig. 3).
Se definieron cuatro grupos de especies de acuerdo con su distribución en el área. El primero incluye a las especies que se presentan en más del 75% de las localidades (euritópicas), el segundo entre el 50 y 74%, el tercero entre 25 y 49% y el último en menos del 25% (estenotópicas) (Apéndice I). En el primer grupo se registraron cuatro especies euritópicas: H. hanno, E. daira, P. proterpia y D. gilippus (24, 23, 20 y 19 localidades, respectivamente). El segundo comprende 16 especies (11% del total: ningún papiliónido). El tercero está conformado por 32 especies (20%). El último que comprende las estenotópicas, incluye el 66% de las especies registradas, 31 de estas especies con una distribución restringida a un solo sitio, la mayoría en El Tepehuaje y Chiautla. En este último grupo se incluyó la mayor parte de los licénidos (41 spp.) reconocidos para la sierra de Huautla.
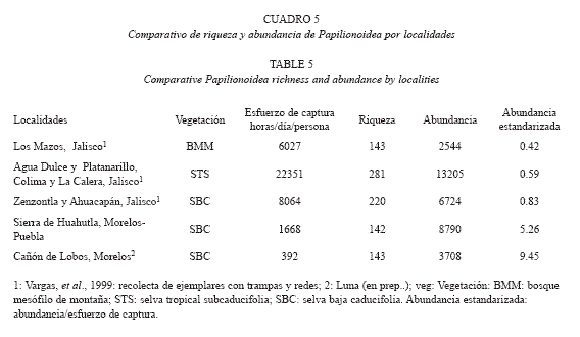
Abundancia total de Papilionoidea: En este análisis, Cañón de Lobos y la sierra de Huautla tuvieron los valores más altos de abundancia estandarizada con respecto a otras localidades con vegetación equivalente (Zenzontla, Ahuacapán) y otros con vegetación de selva baja subcaducifolia (La Calera, Agua Dulce y Platanarillos), a pesar de la gran diferencia del esfuerzo de recolecta que existió entre ellas (Cuadro 5).
Abundancia por familia: La familia más abundante en la sierra de Huautla es Nymphalidae (4154 ejemplares), y le sigue Pieridae (3324), que consideradas de manera conjunta constituyen el 85% de los especímenes capturados (Fig. 4); el 15% restante lo conforman Lycaenidae (1108) y Papilionidae (204 ejemplares).
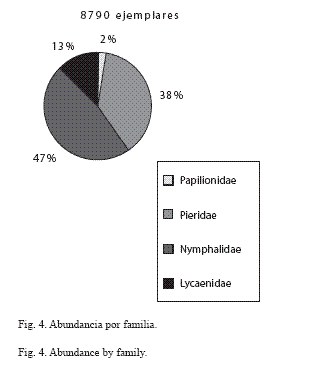
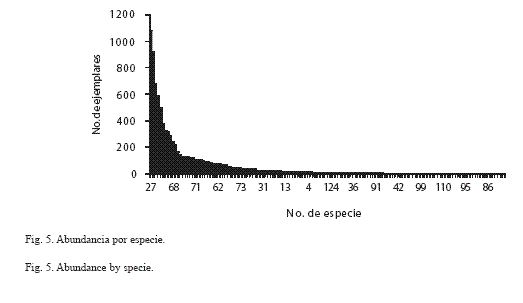
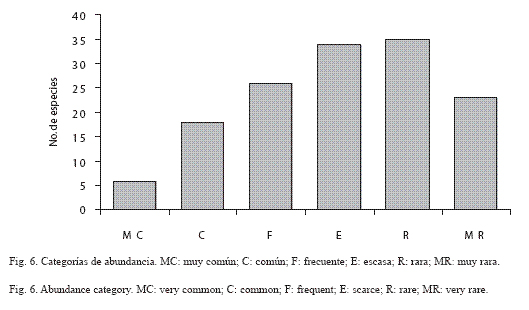
Abundancia por especie: En la sierra de Huautla los piéridos E. daira, P. proterpia, P. sennae y E. arbela se ubicaron entre las diez especies más abundantes, mientras que 13 especies de licénidos entre las especies raras: C. goodsoni, S. cestri, S. albata, C. stillaticia y L. sessilis (Apéndice I). E. daira y P. proterpia juntas tuvieron más del 22% del total de los especímenes capturados (12.27% y 10.45% respectivamente), mientras que 118 especies estuvieron representadas cada una por menos del 1% del total (Fig. 5; Apéndice I). De las seis categorías de abundancia relativa definidas en este trabajo, seis especies fueron muy comunes (MC): E. daira, P. proterpia, H. hanno, C. themis, M. cyananthe y P. sennae; 18 fueron comunes (C), 26 frecuentes (F), 34 escasas (E), 35 raras (R) y 23 muy raras (MR); las tres primeras categorías, MC, C y F contienen el 94% de los ejemplares recolectados y las restantes tan solo el 6% del total (Apéndice I; Fig. 6).
Abundacia por localidad: En El Tepehuaje y El Limón se registraron los máximos de abundancia estandarizada, aunque en Agua Fría, Tepexco y San Miguel Tecolacio también se escontraron valores altos (Cuadro 6: Fig. 7). Barranca El Almagre, Santa Cruz de Achichipico, Axochiapan y Río Cuautla tuvieron la menor abundacia estandarizada de toda el área de estudio.
Fenología: En el análisis de la fluctuación temporal de la comunidad de Papilionoidea se detectaron dos máximos de riqueza, el primero de abril-mayo y el segundo en septiembre-noviembre, así como un valor mímino en junio (Figura 8: Apéndice II).
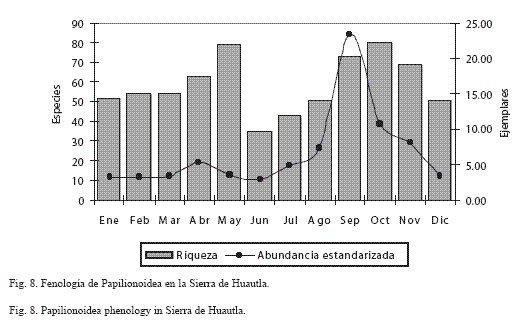
Los valores máximos de abundancia ocurrienron en la época húmeda y calidad, y los menores en la época seca. Se encontró que las especies muy comunes (MC) se presentaron durante todo el año (v. gr. E. daira, P. proterpia, H. hanno, C. themis, M. cyananthe y P. sennae), con poblaciones mayores durante la época seca; las especies comunes (C) y frecuentes (F) estuvieron presentes a lo largo del año (v. gr. A. amathea, A. jatrophae, D eresimus y P. agarithe) o solo en una estacion (v. gr. seca: M. rubricata, y lluvias: M. elva y T. anomalus). En el caso de las especies escasas (E) (H. glauconome, H. costaricensis y P. epidaus), raras (R) (P. erithalion y R. palegon) y muy raras (MR) (T. weymeri, S. cestri y P. bithias), se encontró que su presencia mensual varió entre el 45% (mayo) al 23% (junio); más del 40% de las especies de estas categorías están presentes en mayo, septiembre y octubre, y más del 30% durante seis meses, lo que significa que estas especies definen la variación estacional en la sierra de Huautla.
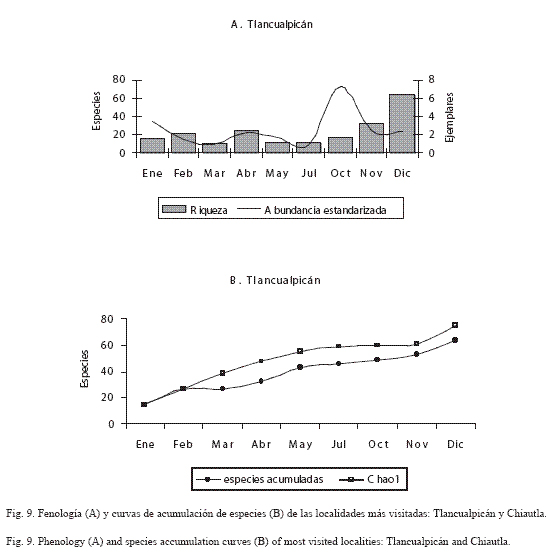
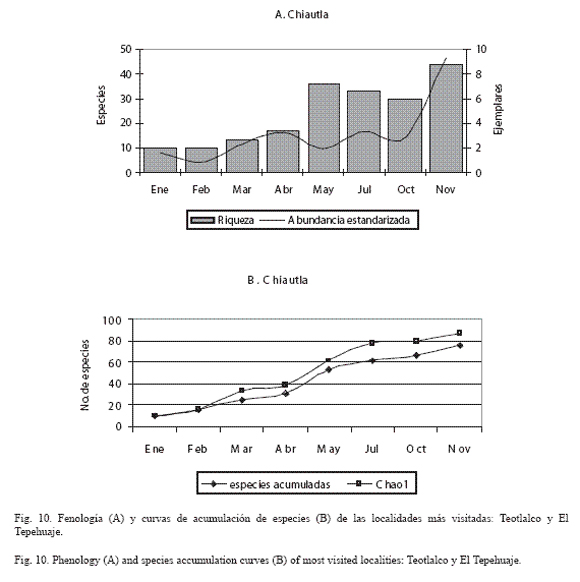
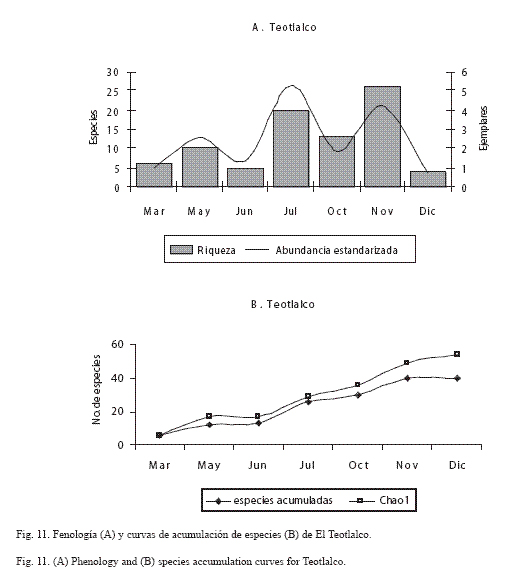
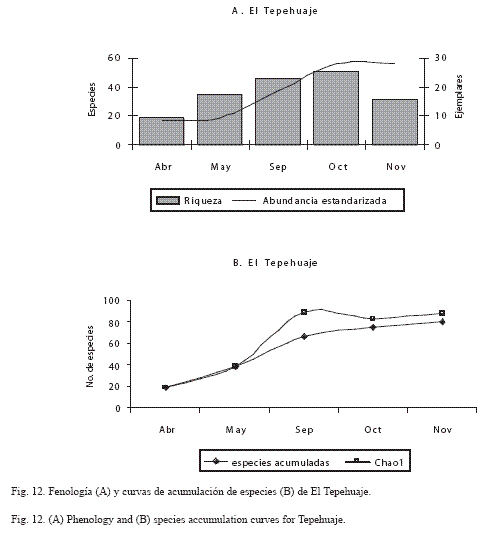
En las figuras 9 a 12 se muestra la fenología y la curva de acumulación de especies de las localidades en donde se efectuó mayor esfuerzo de recolecta (horas/persona) y más visitas por mes (Chiautla, Tlancualpicán, Teotlalco y El Tepehuaje); en estas figuras, se observa claramente que la riqueza fue mayor durante los meses húmedos (octubre-noviembre).
Discusión
Lista sistemática y riqueza: Desde finales del siglo XIX, más del 90% de las recolectas en México se han efectuado en las áreas húmedas, principalmente en los bosques tropicales perennifolios del sur y sureste (Salinas et al. 2004) y en los bosques mesófilos de montaña (Luis et al. 2003b). En las áreas con clima semicálido y subhúmedo, así como en las áreas templado secas y de coníferas, las recolectas han sido esporádicas y hay un menor número de estudios. Por lo que la lista de especies que se presenta es de gran importancia, ya que se describe la variación estacional y geográfica de Papilionoidea en una comunidad vegetal poco estudiada.
La riqueza de la sierra de Huautla (142 especies) hasta ahora es la menor registrada para un estudio faunístico efectuado en selvas bajas caducifolias y subcaducifolias, a pesar de ser la única que está compuesta por un conjunto de localidades de diferente altitud y distribuidas en dos estados. En el Cuadro 5, se observa que aunque el esfuerzo de recolecta en el área de estudio fue cuatro veces mayor al efectuado en Cañón de Lobos (Luna-Reyes 2007), la diferencia es solo de una especie, a pesar que están situados en una región similar con un solo tipo de vegetación; por el contrario, con La Calera (240 especies), Ahuacapán (197 especies) y Zenzontla (154 especies), Jalisco (vargas et al. 1999) que se ubican en una región de mayor heterogeneidad altitudinal, climática y vegetacional, en donde el esfuerzo empleado fue entre dos y cinco veces mayor al efectuado en la sierra de Huautla, la diferencia con la localidad más rica fue de 98, es decir, 70% más especies de papilionoideos. Solo Zenzontla presenta selva baja caducifolia y muestra un número de especies cercano a sierra de Huautla.
En México se han realizado muy pocos trabajos en la selva baja caducifolia que describan la distribución estacional y geográfica de la fauna de Papilionoidea, debido a la baja riqueza presente en estas comunidades y a la época seca muy larga y marcada, a pesar de las grandes extensiones que ocupa este ecosistema en todo el país, principalmente en la región del Pacífico. Además de las 142 especies, esta región se caracteriza por la presencia de 18 especies endémicas a México (Apéndice I), valor que proporcionalmente es superior (12.7%) al encontrado a nivel nacional (8.66%); lo que coincide con el hecho que en las zonas áridas y semiáridas se encuentra el mayor número de endemismos (Luis et al. 2003b).
Con este trabajo la lista de especies de Puebla se incrementó en 25 especies, (ver Apéndice I), debido a que la mayoría de los muestreos previos se habían realizado en el norte del Estado, principalmente en la región húmeda de la Barranca de Patla (área de transición entre selva alta, selva mediana y bosque mesófilo de montaña) y zonas contiguas que han sido recolectadas de forma intensiva, principalmente por aficionados. El resto del Estado ha sido poco o nada explorado (González 1996), incluyendo la parte suroeste que presenta una superficie extensa de selva baja caducifolia donde se localiza la zona de estudio.
Esfuerzo de captura: Los resultados indicaron que la metodología de Chao2 es practicable en recolecciones explorativas o mínimas, pero no en aquellas sistemáticas e intensivas como fue este caso. Al considerar el modelo de Clench (1979), se estimó que falta por registrar el 27% de las especies de la región (aproximadamente 52 especies) (Fig. 2).
Posiblemente sean especies de Lycaenidae (ver riqueza por familia) que con frecuencia son raras. De las cinco localidades más diversas (Cuadro 5), dos tuvieron un esfuerzo mayor a 200 horas/persona (Chiautla y Tlancualpicán) y tres menor o igual a 90 horas/persona (Agua Fría, El Limón y El Tepehuaje); entre El Tepehuaje y Chiautla con 80 y 75 especies respectivamente, la diferencia es de 161 horas/ persona, o como el caso de Teotlalco que con un esfuerzo de recolecta de 120 horas/persona se registraron únicamente 40 especies. La sierra de Huautla es una unidad biótica homogénea (selva baja caducifolia) y la diferencia entre las localidades se basa en la heterogeneidad de los microhábitats en cada una, lo que afecta de manera directa su riqueza específica; el esfuerzo de captura efectuado en esta gama de microhábitats es esencial para reconocer la riqueza total por localidad y por región.
Riqueza por familia: La riqueza por familia obtenida en este trabajo coincide proporcionalmente con los valores que se dan a nivel nacional y estatal, presentando a Nymphalidae como la más rica y a Papilionidae con la menor diversidad; sin embargo, la proporción para Pieridae se duplica (16%) y la de Lycaenidae se reduce (8%) (Cuadro 2). Es posible que el área de distribución promedio de las poblaciones de papiliónidos y piéridos sea más amplia con respecto al de las otras familias en la selva baja caducifolia y subcaducifolia. En México, estos tipos de vegetación se presentan en la región del Pacífico en forma casi continua en la cota altitudinal de los 600 a los 1800 m, lo que favorece su distribución; además, estos resultados podrían indicar que las poblaciones de ninfálidos y licénidos están más restringidas en condiciones extremas. Oñate-Ocaña et al. (2005) señalan que a nivel nacional algunas especies de Papilionidae y Pieridae tienen un área de distribución restringida; sin embargo, local y regionalmente, de acuerdo con los datos obtenidos en este trabajo y el de Luna-Reyes (2007), las áreas de distribución de las especies de estas familias son más amplias que las de Nymphalidae y Lycaenidae; al aplicar el índice que estos autores proponen se esperaría entonces que las especies de Papilionidae y Pieridae de este trabajo tuvieran áreas de distribución mayor. Por otro lado, las especies obtenidas de estas dos últimas familias son muy vágiles y en ocasiones están asociadas con ambientes perturbados (principalmente Pieridae), lo cual es muy común en el área de estudio por la alteración antropogénica acentuada. En el caso particular de los licénidos, su aparente disminución puede deberse al esfuerzo de captura empleado, pues las especies de esta familia requieren un mayor esfuerzo de recolecta, así como una recolecta más orientada o especializada, considerando microhábitats y horarios más específicos para muchos de sus imagos.
Riqueza y distribución por localidad: La riqueza por localidad depende de la heterogeneidad ecológica que presenta cada una de ellas, pues como se observa claramente en el Cuadro 6, el esfuerzo de captura no es la explicación fundamental, ya que el sitio con mayor riqueza no es aquel en el que se efectuó el mayor esfuerzo de recolecta. Por ejemplo, en una de las localidades más pobres como Barranca El Almagre (8 spp., 40 horas/persona), el esfuerzo de recolecta fue igual al de Pitzotlán que cuenta con 31 especies. Chiautla, El Tepehuaje y El Limón presentan la mayor riqueza y son las localidades donde se observó el menor impacto de las actividades antropogénicas, a pesar que son utilizadas por los habitantes de los poblados aledaños como áreas de cultivo y de pastoreo, principalmente para ganado vacuno. En Tlancualpicán, Jolalpan, Tepexco, Puente Marquéz y Río Cuautla existen cuerpos de agua permanentes (ríos de 10 m de ancho o más), donde crece vegetación hidrófila abundante y herbáceas cuyas flores son un recurso alimentario importante para los imagos. Otras localidades con cuerpos de agua presentes solo en la época lluviosa son Huehuetlán, Agua Fría, Atlacahualoya, Ixtlilco El Grande, Ixtlilco El Chico y Santa María Xuchapa. En Los Sauces, Los Linderos y Barranca El Almagre se ubican cañadas donde la humedad propicia una mayor riqueza de especies vegetales; final-mente Axochiapan, Santa Cruz Achichipico, viborillas y San Miguel Tecolacio corresponden a lugares abiertos con algunas lomas de poca pendiente; estas localidades presentaron los valores más bajos de riqueza (Cuadro 6).
La distribución por familia, coincide con los patrones marcados en otras regiones, en donde las familias más ricas (Nymphalidae y Lycaenidae) son las que predominan, solo destaca el hecho que en el 40% de las localidades no se registraron especies de la familia Papilionidae.
Abundancia total de Papilionoidea: La característica principal de las selvas bajas es la abundancia alta de varias especies de Papilionoidea; sin embargo, en la sierra de Huautla ésta depende básicamente de cinco especies, que en conjunto constituyen del 40 al 60% de los ejemplares registrados: E. daira, P. proterpia, H. hanno, C. themis y M. cyananthe, que conforman el 42.76% de la abundancia.
Abundancia por familia: En general, las proporciones de abundancia coinciden con trabajos realizados en áreas tropicales, por debajo de los 1 800 m de altitud, en donde la mayoría de las especies de papiliónidos y licénidos presentan densidades bajas. Ehrlich (1960) menciona que Lycaenidae es una familia muy diversa pero con poblaciones reducidas, a diferencia de los piéridos que están menos diversificados pero tienen poblaciones mayores, son más conspicuos, y fáciles de recolectar y observar. Además Pieridae se caracteriza por contener especies colonizadoras, que son favorecidas con la perturbación de las selvas bajas, debido a que sus larvas están asociadas a plantas secundarias de gran éxito.
Abundancia por especie: El 65% de las especies de la sierra de Huautla están representadas por menos de 22 ejemplares cada una de ellas, 23 de las cuales solo por un ejemplar (Fig. 6). Es probable que muchas de estas especies no se vuelvan a recolectar en la zona durante mucho tiempo, ya que su obtención es más casuística debido a su rareza relativa y al hecho de que no se reconoce su status de residencia. El 35% restante incluye 50 especies, de las cuales más del 70% de éstas se registrarán año tras año, debido a su abundancia relativa y que a muchas de ellas presentan una o más generaciones anuales. Las especies euritópicas E. daira, P. proterpia y H. hanno fueron las más abundantes en la zona (Apéndice II).
Abundancia por localidad: La abundancia en El Tepehuaje se debe a la gran cantidad de piéridos registrados v. gr. P. proterpia (447), E. daira (224), E. arbela (144) y P. sennae (93) los cuales hacen un total de 908 ejemplares que constituyen más del 10% del total de papilionoideos y el 28.25% de los piéridos registrados (Cuadro 6; Fig. 7). Estas especies se caracterizan por su amplia distribución, gran vagilidad y versatilidad para adecuarse a los recursos alimentarios disponibles, tanto en su estadio larval, como en el adulto (Courtney 1986). En las localidades donde se registró la menor abundancia, se observó el efecto del impacto humano en el hábitat, pues gran parte del área se ha transformado en tierras de cultivo (maíz, sorgo, jitomate y otros) y zonas de pastoreo para ganado vacuno y ovino, por lo que son frecuentes zonas de vegetación secundaria, con pocos manchones de selva caducifolia original, favoreciendo exclusivamente a los piéridos y afectando directamente a las poblaciones de licénidos.
Fenología: La distribución estacional de la riqueza no coincide con el patrón esperado para un área que presenta un clima cálido subhúmedo con régimen de lluvias de verano muy marcado y una época seca extrema, donde la fenología de la vegetación cambia drásticamente entre la época seca y húmeda, y en la que se esperaría que la riqueza y abundancia de los Papilionoidea varíe de manera similar (Fig. 8; Apéndice II). Lo peculiar de la sierra de Huautla es que la época seca se divide en dos, la primera de diciembre a marzo con un 38% de la riqueza, bastante alta para una zona semiárida, y la segunda que comprende los meses más secos en la región (abril-mayo) con 44.4 y 55.6% respectivamente, este último muy similar al de mayor diversidad de la época húmeda (octubre), patrón distinto al encontrado en otras regiones (Luis y Llorente 1990, Luis et al. 1991, vargas et al. 1992, 1999, valencia 1999, Pozo et al. 2003), en donde los meses más pobres son abril-mayo, por ser en general los meses más secos. De acuerdo con Balcázar (1988), Arteaga (1991), De la Maza R. et al. (1995a,b), Lara (1999), vargas et al. (1999) y Luna-Reyes (2007), en los ambientes secos y semisecos el número de especies y de ejemplares se reduce notablemente durante la época seca. Un factor metodológico que influyó a que el patrón de la sierra de Huautla fuera distinto, es el esfuerzo de recolecta entre las dos estaciones, ya que durante la época seca (enero-mayo) se efectuó más del doble (1059 horas/persona) que en la época húmeda (julio-noviembre) (437 horas/persona). Este esfuerzo diferencial mucho mayor en la época seca es único para un trabajo sistemático en México.
La diferencia entre los picos de riqueza en la época seca (marzo-mayo) y húmeda (agosto-octubre), es de solo una especie (mayo: 79 especies y octubre: 80 especies). Tal riqueza y similitud principalmente se debe a las 92 especies que ocupan la categoría de Escasas, Raras y Muy Raras (Fig. 8), que ocurren durante algunos meses, constituyendo entre el 9 y el 40% de la riqueza mensual, pero con una representación mayor en mayo (36 spp.). Por el contrario, las especies Muy Comunes y gran parte de las Comunes (60%) estuvieron presentes todo el año, aunque la mayoría de esta última categoría (más del 90%) principalmente se concentró en la época húmeda. Estos resultados coinciden con Pozo et al. (2005), quienes consideran que el cambio en la fenología de la fauna de una región se debe a las especies univoltinas y de baja densidad.
A diferencia de la riqueza, la abundancia estandarizada presenta una distribución típica, con el máximo durante la época húmeda y una baja sustancial en la época seca (Fig. 8). Esta marcada estacionalidad de las poblaciones de papilionoideos fue detectada de manera muy clara con los valores de abundancia estandarizada, patrón que se corresponde al encontrado en otros trabajos realizados en zonas de selva baja caducifolia v. gr. Chorros del varal, Michoacán (Arteaga 1991) y Cañón de Lobos, Morelos (Luna-Reyes 2007), incluso con otros desarrollados en ambientes más húmedos como los de sierra de Atoyac (vargas et al. 1992) y sierra de Manantlán (vargas et al. 1999).
El análisis de la fenología de los papilionoideos de las localidades más ricas y con mayor abundancia estandarizada (Tlancualpicán, Chiautla, Teotlalco y El Tepehuaje) mostró que la comunidad de mariposas presenta un patrón estacional ya descrito por Wolda (1988).
En áreas tropicales como en la sierra de Huautla, la precipitación puede ser el factor climático determinante en la dinámica de las poblaciones de insectos, especialmente de los fitófagos como las mariposas.
Agradecimientos
Queremos agradecer a todas las personas que ayudaron en el trabajo de campo, quienes hicieron posible este artículo. A Omar Ávalos por su ayuda en la aplicación de los métodos automatizados y sus comentarios al texto. Así como el apoyo recibido de los proyectos CONABIO R185, DGAPA IN 212006, PAPIME PE 201507y CONACyT 83237. De manera especial a la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Un revisor anónimo ayudó a mejorar sensiblemente este trabajo.
Resumen
La Cuenca del Balsas es una región singular donde se encuentra una representación significativa de la riqueza y el endemismo de la flora, la herpetofauna y la avifauna mexicanas. Sin embargo, es escaso el conocimiento respecto a los papilionoideos (Lepidoptera), en especial aquellos asociados con la selva baja caducifolia. Aquí se presenta un estudio sobre la distribución local y temporal de los Papilionoidea de la cuenca alta en su vertiente oriental al río Balsas, en particular de la sierra de Huautla y áreas adyacentes. Se efectuaron 25 periodos de trabajo en campo para 24 localidades de sbc de los estados de Morelos y Puebla, ubicadas en altitudes entre los 900 y 1300 m. Se capturaron 8790 ejemplares que corresponden a 83 géneros y 142 especies de Papilionoidea (sensu Kristensen, 1975: Papilionidae, Pieridae, Nymphalidae y Lycaenidae) 18 endémicas a México, además 25 especies constituyen nuevos registros para Puebla. Con este trabajo, ambos estados se ubican entre los siete más ricos de México. En el análisis de la distribución local de la riqueza y la abundancia de los Papilionoidea, se encontró que la sierra de Huautla es la que presenta la menor riqueza con respecto a otras regiones con sbc y uno de los valores más altos de abundancia estandarizada. El Tepehuaje (Morelos) fue la localidad donde se encontraron los valores máximos de todo el estudio. La lepidopterofauna de la sierra de Huautla presenta un patrón estacional atípico de la riqueza, mientras que su abundancia tiene un comportamiento claramente estacional en el que las poblaciones de papilionoideos fueron bastante mayores durante la época húmeda.
Palabras clave: Bosque tropical, Cuenca del río Balsas, fenología, Mariposas, riqueza, selva baja caducifolia.
Recibido 04-VIII-2007. Corregido 30-VI-2008. Aceptado 31-VII-2008.
Referencias
Arita, H.T. & L. León. 1993. Diversidad de mamíferos terrestres. Ciencias (número especial) 7: 13-22. [ Links ]
Arteaga, G.L.E. 1991. Aspectos de la distribución y fenología de los Papilionoidea (Insecta: Lepidoptera) de la Cañada de los Chorros del varal, municipio de Los Reyes, Michoacán. Tesis Licenciatura. Escuela de Biología. Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México. 70 p. [ Links ]
Balcázar, M.A. 1988. Fauna de mariposas de Pedernales, municipio de Tacámbaro, Michoacán (Lepidoptera: Papilionoidea y Hesperioidea). Tesis Licenciatura. Escuela de Biología. Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México. 83 p. [ Links ]
Beccaloni, G.W. & K.J. Gaston. 1995. Predicting the species richness of neotropical forest butterflies: Ithomiidae (Lepidoptera: Nymphalidae) as indicators. Biol. Conserv. 71: 77-86. [ Links ]
Beutelspacher, C.R. 1984. Mariposas de México. Prensa Médica Mexicana. México D.F., México. 128 p., 20 láminas. [ Links ]
Brown, Jr., K.S. 1997. Diversity, disturbance and sustainable use of Neotropical forest: Insects as indicators for conservation monitoring. J. Ins. Conserv. 1:1-18. [ Links ]
Clench, H.K. 1979. How to make regional lists of butterflies: some thoughts. J. Lep. Soc. 33: 215-231. [ Links ]
Colwell, R.K. & J.A. Coddington. 1994. Estimating terrestrial biodiversity through extrapolation. Phil. Trans. R. Soc. London 345: 101-118. [ Links ]
CONABIO. 2000. Sierras de Taxco-Huautla (RTP-120). Regiones Terrestres Prioritarias de México D.F., México. [ Links ]
Courtney, S.P. 1986. The ecology of pierid butterflies: dynamics and interactions. Adv. Ecol. Res. 15: 51-131. [ Links ]
De la Maza, E.R. 1975. Notas sobre lepidópteros de Rancho viejo y Tepoztlán, Morelos, México. Primera parte: Papilionoidea. Rev. Soc. Mex. Lep. 1: 43-61. [ Links ]
De la Maza, R.R. 1987. Mariposas mexicanas. Fondo de Cultura Económica. México D.F., México. 302 p. [ Links ]
De la Maza, E.R., A. White, & R. De la Maza. 1995. Exploración de factores compensatorios que permiten el refugio de Rhopalocerofauna hidrófila en cinco cañadas de clima subhúmedo en Morelos, México. Rev. Soc. Mex. Lep. 16: 1-63. [ Links ]
De la Maza, E.R., A. White & A. Ojeda. 1995. La horofauna hidrófila de la Cañada de la Toma Tilzapotla, Morelos, México (Lepidoptera-Rhopalocera). Rev. Soc. Mex. Lep. 15: 1-35. [ Links ]
Devries, J.P. 1987. The butterflies of Costa Rica and their natural history. Papilionidae, Pieridae, Nymphalidae. Princeton University, Nueva Jersey, EEUU. 327 p. [ Links ]
Díaz, E. & J. Soberón. 2005. Statistical estimation and model selection of species-accumulation functions. Conserv. Biol. 19: 569-573. [ Links ]
Ehrlich, P.R. 1960. Lepidoptera, p. 459-473. In McGraw-Hill. Encyclopedia of Science and Technology. McGraw-Hill, Nueva York, EEUU. [ Links ]
Ehrlich, P.R. & A.H. Ehrlich. 1961. The Butterflies. Brown, Dubuque, Iowa, EEUU. 262 p. [ Links ]
Emmel, T.C. 1975. Butterflies, their world, their life cycle, their behavior. Knopf, Nueva York, EEUU. 260 p. [ Links ]
Escalante, P., A. Navarro & T. Peterson. 1993. Ageographic, ecological and historical analysis of land bird diversity en Mexico, p. 282-307. In T.P. Ramamoorthy, R. Bye, A. Lot & J. Fa (eds.). Biological Diversity of Mexico: Origins and Distribution. Oxford, Nueva York, EEUU. [ Links ]
Espinosa, P.H. 1993. Riqueza y diversidad de peces. Ciencias (número especial) 7: 77-84. [ Links ]
Fa, J.E. & L.M. Morales. 1993. Patterns of Mammalian Diversity in Mexico, p. 319-361. In T.P. Ramamoorthy, R. Bye, A. Lot & J. Fa (eds.). Biological Diversity of Mexico: Origins and Distribution. Oxford, Nueva York, EEUU. [ Links ]
Feria, P. 1997. Diversidad y distribución avifaunística en una localidad del municipio de Chiautla de Tapia, Puebla. Tesis Licenciatura, Facultad de Estudios Superiores Zaragoza, UNAM, México D.F., México. 66 p. [ Links ]
Flores, V.O. 1993. Riqueza de los anfibios y reptiles. Ciencias (número especial) 7: 33-42. [ Links ]
Flores, V.O. & P. Gerez. 1994. Biodiversidad y conservación en México: vertebrados, vegetación y uso del suelo. UNAM-CONABIO, México D.F., México. 439 p. [ Links ]
García, E. 1981. Modificaciones al Sistema de Clasificación Climática de Köppen. UNAM, México D.F., México. 241 p. [ Links ]
González, M.L. 1996. Listado faunístico de los papilionoidea del estado de Puebla, con especial referencia a la zona de Barranca de Patla. Tesis de Licenciatura, Facultad de Ciencias, UNAM, México D.F., México. 112 p. [ Links ]
Hoffmann, C.C. 1940. Catálogo sistemático y zoogeográfico de los lepidópteros mexicanos. Primera parte. Papilionoidea. An. Inst. Biol. UNAM. 11: 639-739. [ Links ]
Howe, W.H. 1975. The butterflies of Noth America. Doubleday, Nueva York, EEUU. 633 p. [ Links ]
INEGI. 1981. Síntesis Geográfica de Morelos. SPP, México D.F., México. [ Links ]
Krebs, C.J. 1985. Ecología. Estudio de la distribución y abundancia. Harla, México D.F., México. 753 p. [ Links ]
Kristensen, N.P. 1975. Remarks on the family level phylogeny of butterflies (Insecta:Lepidoptera, Rhopalocera). Zool. Syst. Evol. Forsch. 14: 23-33. [ Links ]
Lamas, G. 1984. Los papilionoidea (Lepidoptera) de la Zona reservada de Tambopata, Madre de Dios, Perú. I: Papilionidae, Pieridae y Nymphalidae. Rev. Peruana Ent. 27: 59-73. [ Links ]
Lamas, G. 2000. Estado actual del conocimiento de la sistemática de los lepidópteros, con especial referencia a la Región Neotropical, p. 253-260. In F. Martín, J.J. Morrone & A. Melic (eds.). Hacia un proyecto para el inventario y estimación de la diversidad biológica en Iberoamérica: PrIBES-2000. SEA. Zaragoza, España. [ Links ]
Lara, A.M.A. 1999. Riqueza de especies y abundancia relativa de la superfamilia Papilionoidea (Lepidoptera) en un bosque de Quercus, en Huitzilac, Morelos. Tesis Licenciatura, Facultad de Estudios Superiores Zaragoza, UNAM, México D.F., México. 69 p. [ Links ]
León, J.L., J. Soberón & J. Llorente. 1998. Assessing completeness of Mexican Sphinx moths inventories through species accumulation functions (Lepidoptera: Sphingidae). Divers. Distrib. 4: 37-44. [ Links ]
Luis, A. & J. Llorente. 1990. Mariposas del valle de México: Introducción e Historia I. Distribución local y estacional de los Papilionoidea de la Cañada de Los Dínamos, Magdalena Contreras, D. F., México. Folia Entomol. Méx. 78: 95-198. [ Links ]
Luis, A., I. vargas & J. Llorente. 1991. Lepidopterofauna de Oaxaca I: distribución y fenología de los Papilionoidea de la Sierra de Juárez. Publ. Esp. Mus. Zool. UNAM. 3: 118. [ Links ]
Luis, A., J. Llorente & I. vargas. 2003. Nymphalidae de México I (Danainae, Apaturinae, Biblidinae y Heliconiinae): distribución geográfica e ilustración. CONABIO-UNAM, México D.F., México. 249 p. [ Links ]
Luis, A., J. Llorente, I. vargas & A.L. Gutiérrez. 2000. Síntesis preliminar del conocimiento de los Papilionoidea (Lepidoptera: Insecta) de México, p. 275-285. In F. Martín, J.J. Morrone & A. Melic (eds.). Hacia un proyecto para el inventario y estimación de la diversidad biológica en Iberoamérica: PrIBES-2000. SEA, Zaragoza, España. [ Links ]
Luis, A., J. Llorente, I. vargas & A.D. Warren. 2003. Biodiversity and biogeography of Mexican butterflies (Lepidoptera: Papilionoidea and Hesperoidea). Proc. Entomol. Soc. Wash. 105: 209-224. [ Links ]
Llorente, J., A. Luis, I. vargas & J. Soberón. 1996. Papilionoidea (Lepidoptera), p. 531-548. In J. Llorente, A.N. García, & E. González (eds.). Biodiversidad, Taxonomía y Biogeografía de los artrópodos de México: Hacia una síntesis de su conocimiento. UNAM, México D.F., México. [ Links ]
Llorente, J., L. Oñate, A. Luis & I. vargas. 1997. Papilionidae y Pieridae de México: distribución geográfica e ilustración. CONABIO-UNAM, México D.F., México. 228 p. [ Links ]
Luna, M.M. 2007. Estudio faunístico sobre Papilionoidea (Lepidoptera) en Cañón de Lobos, Yautepec, Morelos. Tesis de Maestría, Facultad de Ciencias, UNAM, México D.F., México. 102 p. [ Links ]
Martín, F. & J.M. Lobo. 2000. Diagnóstico sobre el conocimiento sistemático y biogeográfico de tres órdenes de insectos hiperdiversos en España: Coleoptera, Hymenoptera y Lepidoptera, p. 287-308. In F. Martín, J.J. Morrone & A. Melic (eds.). Hacia un proyecto para el inventario y estimación de la diversidad biológica en Iberoamérica: PrIBES-2000. SEA, Zaragoza, España. [ Links ]
Mittermeier, R.A. & C. Goettsch. 1992. La importancia de la diversidad biológica de México, p. 63-73. In J. Sarukhán & R. Dirzo (eds.). México ante los retos de la Biodiversidad. CONABIO, México D.F., México. [ Links ]
Morón, M.A. & J.E. valenzuela. 1993. Estimación de la Biodiversidad de Insectos en México; Análisis de un caso. Rev. Soc. Mex. Hist. Nat. 44: 303-312. [ Links ]
Navarro, S.A. & H. Benítez. 1993. Patrones de riqueza y endemismo de las aves. Ciencias (número especial) 7: 45-54. [ Links ]
Oñate, L., J. Llorente & J. Meave. 2005. Ideas para un índice de amplitud biogeográfica, p. 189-202. In J. Llorente y J.J. Morrone (eds.). Regionalización biogeográfica en Iberoamérica y tópicos afines. Prensas de Ciencias, Facultad de Ciencias, UNAM, México D.F., México. [ Links ]
Pollard, E. 1977. A method for assessing changes in the abundance of butterflies. Biol. Conserv. 12: 115-134. [ Links ]
Pozo, C., A. Luis, S. Uc, N. Salas & A. Maya. 2003. Butterflies (Papilionoidea And Hesperoidea) of Calakmul, Campeche, México. Southw. Nat. 48: 505-525. [ Links ]
Pozo, C., J. Llorente, A. Luis, I. vargas & N. Salas. 2005. Reflexiones acerca de los métodos de muestreo para mariposas en las comparaciones biogeográficas, p. 203-215. In J. Llorente & J.J. Morrone (eds.). Regionalización biogeográfica en Iberoamérica y tópicos afines. Prensas de Ciencias, Facultad de Ciencias, UNAM, México D.F., México [ Links ]
Pyle, R.M. & A.A. Knopf. 1981. The Audubon Society field guide to North American butterflies. Chanticleer, Nueva York, EEUU. 916 p. [ Links ]
Rzedowski, J. 1992. El endemismo en la Flora Fanerogámica de México, p. 337-359. In G. Halffter (ed.). La Diversidad Biológica de Iberoamérica I. Acta Zool. Mex. vol. Esp. [ Links ]
Salinas, J.L., A. Luis & J. Llorente. 2004. Papilionoidea of the evergreen tropical forest of Mexico. Jour. Lep. Soc. 58: 125-142. [ Links ]
Scott, J.A. 1985. The phylogeny butterfly (Papilionidae and Hesperidae). Jour. Res. Lep. 23: 1-60. [ Links ]
Scott, J.A. 1986. The butterflies of North America. A natural history and field guide. Stanford, California, EEUU. 583 p. [ Links ]
Smart, P. 1989. The illustrated Encyclopedia of the butterfly world. Crescent, Nueva York, EEUU. 275 p. [ Links ]
Soberón, J. & J. Llorente. 1993. The use of species accumulation curves for the prediction of species richness. Conserv. Biol. 7: 480-488. [ Links ]
Trejo, I. & R. Dirzo. 2000. Deforestation of seasonally dry tropical forest: a national and local analysis in Mexico. Biol. Conserv. 94: 133-142. [ Links ]
Tyler, H., K.S. Brown & K. Wilson. 1994. Swallowtail butterflies of the Americas. A study in biological dynamics, ecological diversity, biosystematics, and conservation. Scientific Publisher, Gainesville, Florida, EEUU. 376 p. [ Links ]
Valencia, G.M.S. 1999. Listado faunístico de los Papilionoidea (Lepidoptera: Rhopalocera) del derrame lávico del volcán Chichinautzin, Estado de Morelos. Tesis de Licenciatura, Facultad de Ciencias, UNAM, México D.F., México. 46 p. [ Links ]
Vargas, I., J. Llorente & A. Luis. 1992. Listado lepidopterofaunístico de la Sierra de Atoyac de Álvarez en el estado de Guerrero: notas acerca de su distribución local y estacional (Rhopalocera: Papilionoidea). Folia Entomol. Mex. 86: 41-178. [ Links ]
Vargas, I., J. Llorente & A. Luis. 1999. Distribución de los Papilionoidea (Lepidoptera: Rhopalocera) de la Sierra de Manantlán (250-1 650 m) en los estados de Jalisco y Colima. Publ. Esp. Mus. Zool. UNAM. 11: 153. [ Links ]
Vences, B.M.O. 2004. Lista lepidopterofaunística del estado de Morelos (Insecta: Lepidoptera: Papilionoidea). Tesis de Licenciatura, Facultad de Ciencias, UNAM, 51 p. [ Links ]
Wolda, H. 1988. Insect seasonality: Why? Annu. Rev. Ecol. Syst. 19: 1-18. [ Links ]

