











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.56 no.2 San José Jun. 2008
Underwater topography determines critical breeding habitat for humpback whales near Osa Peninsula, Costa Rica: implications for Marine Protected Areas
1. Fundación Vida Marina, Drake Bay, Península de Osa, Costa Rica.
2. PROCMAR: Programa de Maestría en Ciencias Marinas y Costeras, Universidad Nacional (UNA), Costa Rica; leninovi1@hotmail.com
3. Escuela de Biología Universidad Nacional, Heredía, Costa Rica; morris118@hotmail.com
Abstract: Migrating humpback whales from northern and southern feeding grounds come to the tropical waters near Osa Peninsula, Pacific of Costa Rica, to reproduce and raise their calves. Planning effective marine protected areas that encompass humpback critical habitats require data about which oceanographic features influence distribution during the breeding period. This study examines the relationship between water depth and ocean floor slope with humpback whale distribution, based on sightings during two breeding seasons (2005 and 2006). Data are from the Southern and Northern subpopulations in the Eastern Tropical Pacific (ETP). Analysis followed the basic principles of the Ecological Niche Factors Analysis (ENFA), where indices of Marginality and Tolerance provide insights on the restrictiveness of habitat use. At a fine scale, physical factors such as water depth and slope define the critical breeding and nursing habitat for M. novaeangliae. Divergence in the subsamples means of depths and slope distribution, with the global mean of the study area in both eco-geographical variables, determine habitat requirements restricted by topographic features such as depths (< 100 m) and slope (< 10%), and locate the key breeding and nursing habitat of the species within the continental shelf domains. Proposed Marine Protected Areas (MPAs) network plans should consider connectivity of Caño island–Drake Bay and the extension of Corcovado National Park maritime borders. Rev. Biol. Trop. 56 (2): 591-602. Epub 2008 June 30.
Key words: Critical habitat, breeding area, nursery area. Megaptera novaeangliae, marine protected area.
Humpback whales (Megaptera novaeangliae) aggregate in winter breeding and calving grounds along continental margins, coastal islands or archipelagos located in tropical waters. Migration to these breeding sites implies a long journey, some whales traveling as far as 11 000 miles, from feeding areas in temperate and circumpolar latitudes. Understanding the physical environmental factors that might play a crucial role in defining the breeding and nursing niche is important for effective planning of Marine Protected Areas intended to encompass critical habitat for humpback whales. Generally whales distribution is restricted to waters of less than 100 fathoms (183m) deep. Records of breeding whales, particularly singing males, have been gathered in open deep waters, sometimes beyond the shelf break (Caldwell et al. 1970, Whitehead and Moore 1982, Mattila and Clapham 1989, Mignucci-Giannoni 1998, Felix and Haase 2001, Acevedo et al. 2003, Swartz et al. 2003, Frantzis et al. 2004, Kaschner 2004, Felix and Haase 2005). At a basin scale, Rasmussen et al. (2007) identified a correlation on breeding sites in Central America with the occurrence of warm waters north of the equator (24 °C -28 °C) as opposed to coastal upwelling and cold tongue development during the austral winter, which create cool surface waters that will extend from the South American coast into the eastern equatorial Pacific. The same authors theorized that avoidance of cold fronts will explain migratory movement, based in the need of a temperature regime at breeding sites, as a major selective force to drive migration.
The feeding areas in the North Pacific Ocean include the coasts of Alaska and California, while breeding grounds encompass the Hawaiians islands and Mexican waters, as well as the Eastern Tropical Pacific coast of Central America. North Pacific humpback whales inhabit a broad region of Central America extending south from the wintering grounds previously described in Mexico all the way to Panama. There is also some evidence of site preferences with some whales returning in multiple years to the Drake Bay area. Humpback whales from this region are almost exclusively animals that use the California, Oregon, and Washington feeding area, with some tendency for animals from Central America to feed in the more southern portions of this feeding area (Rasmussen et al. 2004).
A Southern Humpback Whale population migrates north, along the southeast Pacific to the coasts of Ecuador, Colombia, Panama and Costa Rica where they breed (Flóres González 1995, Félix and Haase 2001, Flóres González et al. 2007). Two important feeding areas are the departure point of these whales; the Antarctic Peninsula and Chiles southern coast (Gibbons et al. 2003, Stevick et al. 2004, Acevedo et al. 2007). Wintering Humpback whales are found along the western coastline of the Eastern Tropical Pacific from May-November, with the greatest numbers occurring in July and August (Félix and Haase 2001). Similarly to their northern counterpart, Southern Humpback whales that forage in the northernmost feeding area along the South American Coast migrate to the northernmost breeding grounds off Costa Rica. Both sub-populations of humpback whales, the Northern and the Southern Pacific groups, overlap in Costa Ricas territorial waters, particularly off the Osa Peninsula.
Humpback whales appear to show a heterogeneous distribution during the breeding period. They segregate into sub-groups according to their age and class composition; for instance, groups containing mother-calf pairs prefer shallow waters of 20 meters or less in depth, single sub-adults also prefer shallow waters, whereas groups of adults occur in the deeper waters further from shore (Félix and Haase 2001).
Humpback whales coastal affiliation has made them vulnerable to anthropogenic activities within coastal communities. in most of the cases, those activities degenerate in chemical pollution, vessel traffic noise, industrial activities and particularly interactions with fishing gear (Reeves et al. 2003). Reports of humpback whales entangled in artisanal gillnets, somewhere else in their Southern Pacific range, are a cause for concern, and evidence suggests an increase in this trend. The development of whale watching programs along the coast of Western South and Central America constitutes another potential source of disturbance for whales. Changes in movement and activity patterns during encounters with tourist boats have been reported in several sites including Ecuador (Corkeron 1995, Scheidat et al. 2004, Félix and Haase 2005).
Conservation and management actions, particularly those addressed toward the establishment of MPAs, should be based on the determination of critical habitat. Critical habitat refers to the portions of a cetacean distribution range that have a key particular value for day to day survival and maintaining a healthy population growth. Therefore, areas that are used for breeding, including all aspects of the courtship, raising calves and migration are part of critical habitat. This concept itself, applied to cetacean conservation, is just recently conceived, yet to be explored and scarcely implemented (Hoyt 2005).
This contribution investigates the relationship between the distribution of humpback whale groups and some of the physical and geographic features that may be related to or determine whales spatial distribution; specifically water depth and slope as indicator of underwater topography. These relationships may be used to predict critical habitat for breeding and nursing humpback whales along the coast of Central America. Therefore, it can serve as a strong argument for the establishment of a network of MPAs, taking into account that it is a priority to characterize geographically and environmentally the essential habitats for the species, and create new Marine Protected Areas all along the regional range of M. novaeangliae in the ETP (Flóres-González et al. 2007).
Materials and methos
Study area: The Osa Peninsula is the southern-most portion of Costa Ricas Pacific Coast. The study area was framed by a polygon limited east by the shorelines of Terraba-Sierpe River System, Drake Bay and Corcovado National Park to Punta Salsipuedes, continuing south following the meridian 83° 35W. The northern and southern limits were represented by the parallels 08°55N, and 8°12N respectively, while the western-most limit of the study area was located at 84°00W. Overall the whole polygon covers an approximated area of 3800 Km2. This area is the target location of whalewatching boats (the platform of observation), and encloses what could be conceived as a local network of MPAs: Corcovado National Park shoreline - Caño Island and Drake Bay.
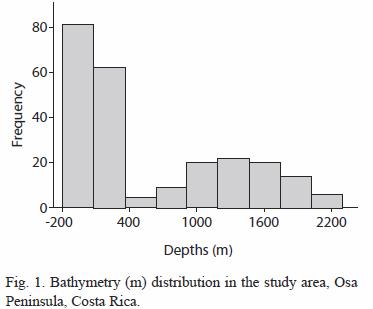
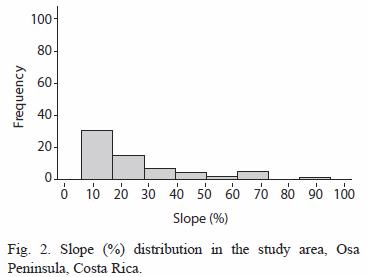
The study areas bathymetry is slightly dominated by shelf depths as shown in figure 1; 55.7 % (n= 133) of the cells analysed were below the 200 m isobath, major depths above the 200 m covered 31.3 % (n= 106) of the study area, with 2158 m as the deepest point considered in the analysis. Another important characteristic of this locale is the gentle slope (figure 2). Cells with slope indices of more than 50% were associated with topographic features such as seamounts or banks located southwest from the shelf.
Data collection: Data gathering on cetacean populations on the coast of Costa Rica has been carried out since 1998 by Fundación Vida Marina aboard their whale-watching vessels. Cetacean surveys in this contribution were accomplished through platform of opportunity (Evans and Hammond, 2004). Two important aspects framed the research effort: 1) Based on the most important cetacean studies in the area (May et al. 2005); boat trips were directed to hot spot areas of occurrence. 2) Observations were performed, during tours in day light hours from 08:00 to 15:00, with hours with cetacean sightings totalizing the effective observation time: the time interval between encounter with a group of whales, until the sighting is terminated (whales are left alone or out of sight). Information was recorded on group size, group composition, behavior, sea surface condition and the geographic position. Position was determined using a portable global positioning system (GPS).
All Humpback whales sightings were selected and pooled together whether northeast Pacific whales or southeast Pacific whales. Sightings were classified in three group classes: mother and calf groups, singer and single sightings. This study used data for a total of 154 groups recorded during 302 boat trips (1812 total hours of field efforts) between 2005 and 2006.
Data Analysis: Habitat characterization was accomplished by correlating spatial distribution of the whales sightings with two kinds of eco-geographical variables (Gregr & Trites 2001, Hirzel et al. 2001, 2002, Compton 2004): a) a continuous variable represented by the bathymetry b) a categorical variable; expressed in relief /depth classes through a slope index.
Using a GIS (Map-Info 5.0) and a bathymetric chart (US Navy 21560), a grid of 239 cells was established in the study area; each one defined as 2 min latitude by 2 min longitude cell. Only cells with a research effort equal to twice the standard deviation (per year, per season) were included in the analysis. A slope index related to sightings of the whales (Mignucci-Giannoni 1998, Oviedo et al. 2005) was calculated through the following formula:
Si = Zmax - Zmin / ZmaxRA X 100
Zmax is the maximum depth of a specific cell; Zmin is the minimum depth, considered 1 m for coastal areas; and ZmaxRA is the maximum depth of the research area.
The basic indices of the Ecological Niche Factor Analysis (ENFA) developed by Hirzel et al. (2002) were calculated for each group class, with the variant of being calculated individually for each eco-geographical variable considered. ENFA analysis was chosen for its properties in lessening the effect due to limitations in using opportunistic data, this approach minimize the bias of presence data and lack of real absent ones. To assess the bathymetry, Zmax was considered per each cell, while the Si was the input data for slope:
Marginality; M = µOs- µMn / 1.96X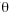 Os
Os
Tolerance; Tol= Mn/Os
Where µOs is the global mean of the study area, per factor in each 2 min. x 2 min cell, µMn is the mean of the distribution of groups of M. novaeangliae, per factor in each cell. Os is the global dispersion of the data, given by the standard deviation, and Mn is the dispersion in the humpback whales specific distributions in each environmental factor. These indices evaluate if the average environmental conditions associated with a particular distribution pattern differ from the global environmental conditions of the study area. Hence an M (Marginality) value closer to one expresses a distribution of values associated with a particular environmental factor and separate from the global conditions. Following Hirzel et al. (2004) suggestion, to ease the interpretation of specialization, the Tolerance (Tol) coefficient; which is the inverse of the specialization, is given as it is an indicator of the species niche breadth. Before calculating the indices, according with the bi-modal distribution of depths in the study area (Figure 2), it was stratified into shelf waters and transition-off shore waters. Data were tested for normality using the Shapiro Wilk test. Significant differences in sub-samples between the study area and the whales sightings were estimated by the Mann Whitney U-test (Analyze it 1.71), while differences between samples of whales sightings was analyzed using ANOVA (Statgraphic plus).
Results
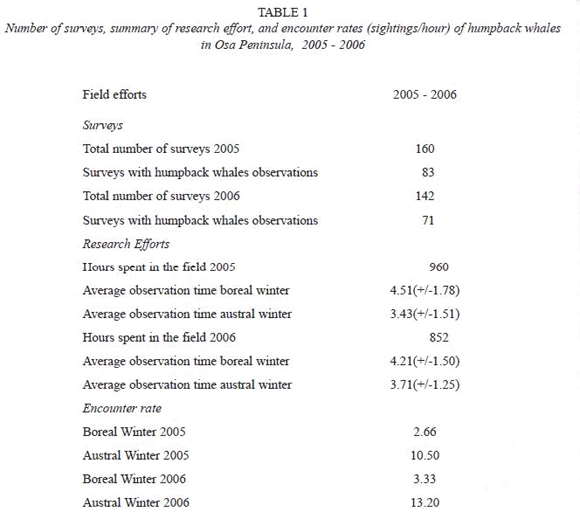
Occurrence of humpback whales in the study area is important through most of the year, as reflected in encounter rate indices (Table 1), which themselves act out as abundance estimates. There is no significant differences between years in both, the northern winter (T= 0.88, p = 0.38) and southern winter (T= -0.81, p = 0.42) encounters. The encounter rates in 2005 and 2006 increase due to a major amount of sightings events towards the months of July-September, that correspond with the breeding season of southern Pacific humpback whales. However, this should not be completely interpreted as an indication of major use of Osa Peninsula breeding site by those southern whales, since more systematic data is still needed.
Distribution of whales in the study area focalized in two major aggregations: around the east coast of Caño island - Drake Bay and the west coast of Osa Peninsula off Corcovado National Park (Figures 3). Whale groups (females and calves, single whales and male singers) overlap within these two focal aggregations, which preclude a determination of differences in habitat use, and enclosed the breeding and nursing portions of their distribution range within these locations in the study area (Figure 4).
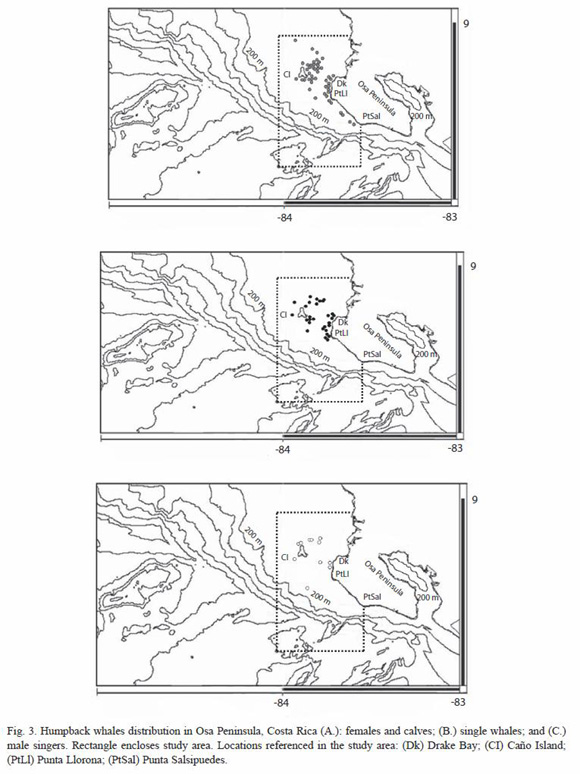
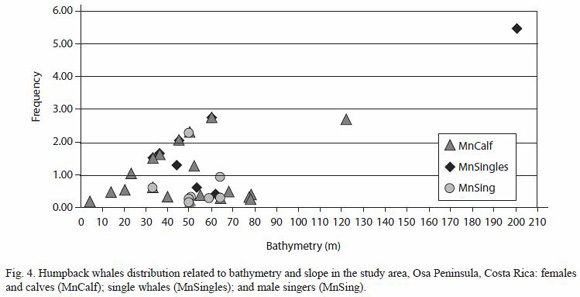
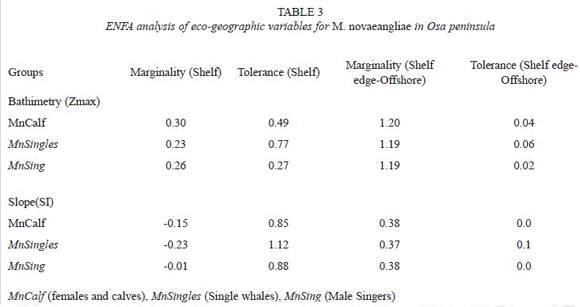
When considering each of the key factors (depths and slope), and statistically compare the whole study area (n= 239 cells) with the distribution of mothers and calves groups (n= 47 cells), single whales (n= 30 cells) and singers (n= 11 cells), there are significant differences in terms of slope, (except for singing whales) and a clear marked tendency for each of the three groups towards waters less than 100 m deep (table 2). These differences indicate a strong affiliation to the shallowest portion of the shelf ecosystem off Osa Peninsula. Between sighting classes; females with calves, singletons and singing whales, there is no significant differences in sightings distribution by depths (f = 0.55, df = 2, p = 0. 58) and slope (f = 0.81, df = 2, p = 0. 45).

This trend is also supported by the marginality and tolerance indices (table 3); mothers and calves grouped together with the loners and singers evidenced a degree of restrictiveness in their realized niche towards shallow waters. Such tendency is sustained by the divergence of the sub-samples mean of slope associated to whales sightings and the shelf waters mean of slope distribution. Additionally such marginality is markedly complemented by a very high tolerance to slopes higher than 1% within shelf limits (<10%). Differences are even more evident when the bathymetry of transition-offshore waters are considered, with whales highly specialized to shallow depths (with a marginality index off chart) and a major lack of tolerance to higher depths off the shelf area.
Discussion
Humpback whale distribution by eco-geographical variables depths and slope: In terms of habitat use; determined by depth and slope; as variables defining underwater topography, whales groups favored shallow bathymetry and gentle slope. Both preferences are consistent with the average characteristics of the entire study area, and with the affiliation to continental shelfs depths and contours that distribution of humpback whales exhibit as a general pattern reported elsewhere (Caldwell et al. 1970, Whitehead and Moore 1982, Mattila and Clapham 1989, Mignucci-Giannoni 1998, Acevedo 2001, Félix and Haase 2001, Acevedo et al. 2003, Swartz et al. 2003, Frantzis et al. 2004, Kaschner 2004, Félix and Haase, 2005): that is, concentrating on or along the edges of shallow banks and islands (Silva et al. 2006) like Caño Island, in areas less than 200 m deep, with fewer whales found in deep water. In the southern Pacific coast of Ecuador, humpbacks are rarely seen offshore in breeding season, seeking depths less than 60m (Félix and Haase 2005).
What makes a contrast to other breeding and nursing sites is that segregation of whale groups within the study area is not quite evident at the spatial microscale. However, the time overlap is rare, since females with calves and competitive male groups were never seen at the same time. For instance, distribution of male singers seemed to follow a similar pattern described by Whitehead and Moore (1982), they report singers in shallow isothermal water over to smooth bottom, where song propagation will be virtually bi-dimensional. However, singers elsewhere had been located on deep waters (Frankel et al. 1995), where offshore locations provide different acoustics propagation conditions. Thus, the factor that determines which area is preferable may be social rather than oceanographic (Frankel et al. 1995). Hence humpback males could be in the whole study area, shown in the overlap with mothers and calves groups. Moreover, the pods without calves could be occurring in same the locations frequented by singers, similar to what Frankel et al. (1995) reported in Hawaii.
Implication for managements and the proposal of a MPAs network off the Osa Peninsula: Encounters of Southern and Northern Pacific humpback whales during transitional months, and the corresponding potential genetic exchange, could only happen in areas of major use as wintering grounds; such as the waters of the Osa Peninsula. Hence, local networking MPAs will certainly reassure the continuity of this interchange.
The latter analysis has described the ecogeographical factors crucial in the definition of nursing and breeding humpbacks whales realized niche in Osa Peninsula waters. When examining the spatial arrangement of M. novaeangliae in the study area, three considerations arise: 1) Whales (whether females and calves groups, lone whales or males singers) are aggregated in two focal locations; Caño islands east coast toward Drake Bay, and all along the west coast of the Peninsula from Drake Bay to Punta Salsipuedes, including waters within Corcovado National Park. Therefore strict protection measures, such as restriction of certain fishing gears or even certain fishing operations, can be equally focalized. 2) A limiting bathymetry of 100 m, plus a limiting slope of less than 10% restrict whales to a very important extension within the continental shelf, which cover a great portion of what would be a proposed new MPA (as well as the extension of the limits of Corcovado National Park) to conform a network. This could also be a strong argument for the restriction of certain pervasive consumptive activities, and reflect the need of management initiatives to other non consumptive ones such eco-tourism (whale-watching). 3) The fact that segregation by groups is not observed within the study area could reflect a lack of differential habitat use. The two focal areas mentioned before, where the whales are clumped in their distribution, have a considerable increased value of not only as breeding site, but as nursing area at the same time. Moreover, since habitat requirements according to the ecogeographical variables analyzed before equally overlap, the entire study area can potentially be considered as a key site in Central America for both nursing calves and reproductive whales.
The latter has strong effects in the value of humpback whales as a flagship-umbrella species, since its wide distribution increases the scope of co-occurrence with sympatric species within the shelf ecosystem. Those co-occurring species might benefit from any protective initiative granted to these migrating whales. The role of M. novaeangliae as a flagship-umbrella species would be greatly enhanced and complemented by a suite of species, that as previously considered, would sympatrically share the same habitat in different portions of it, for instance; one of the most important coral reef patches and associated communities in the Pacific coast of Costa Rica, overlap with humpback whales occurring in Caño island, while the most important population of Strombus galeatus; depleted elsewhere in the countrys coastline, share the same distribution along the southern coast of Corcovado National Park (Cortéz and Jiménes 1996, Alvarado et al. 2007). It is valuable to highlight the value of the study area for other major predators as: other cetaceans (>11 species), sharks and billfish species. The Seascape Species Approach could be an alternative to create such suite.
Migrating humpback whales along the Eastern Tropical Pacific basin (ETP) have been an effective means to promote and reinforce conservation measures in important protected areas along its distribution range, particularly in the Southeast Pacific coast of South America, around areas such as Gorgona National Park in Colombia and Machalilla National Park in Ecuador. As with any trans-zonal species, humpback whales greatly benefit not only with local conservation efforts, but with a cooperative regional conservation initiatives. Setting a priority in creating a network of MPAs that will enclose Drake Bay, Caño island and Corcovado National Park (using as one of the arguments the whales habitat use: breeding and nursing site), will comply with the conceptual framework, guidelines and priority activities established in the Conservation strategy of the South-eastern Pacific humpback whale (Flóres-González et al. 2007); particularly those addressing conservation in situ and the creation of new protected areas. Such initiative will be a great asset to regional conservation efforts in the ETP basin.
Acknowledgments
We thank the staff of Vida Marina del Sur Foundation, including Sierra Goodman, for her input during fieldwork and on the manuscript, as well as for her logistical support. Roy Sancho Rojas, Carlos Luis Rivas Lopez and Luis Taleno provided important support during fieldwork. Manuscript was improved thanks to the valuable insights of Trond Larsen, Alex Zerbini, D.K. Odell, Miguel iñiguez and F. Felix. This investigation has been funded by The Evergreen Foundation and Vida Marina Foundation.
Resumen
Las ballenas jorobadas viajan a aguas tropicales para reproducirse y criar a sus ballenatos. Es importante entender las características oceanográficas que influencian su distribución para lograr una planificación efectiva de áreas marinas protegidas con hábitats críticos para estos cetáceos. Este estudio examina la relación entre la profundidad, la pendiente del suelo oceánico y la distribución de estas ballenas, usando avistamientos del 2005 y 2006 en la costa Pacífica de la Península de Osa, Costa Rica (temporada de reproducción del sur y norte en el Pacífico Tropical Oriental). Usamos Análisis de Factores de Nicho Ecológico (ENFA por sus siglas en inglés), donde los índices de Marginalidad y Tolerancia ilustran las restricciones en uso de hábitat. En una escala local, factores físicos como la profundidad y la pendiente definen el hábitat crítico de reproducción y cría de M. novaeangliae. Las divergencias entre las medias de las sub-muestras y la media global del área de estudio en ambas variables eco-geográficas, determinan las limitaciones en requerimientos de hábitat en aspectos topográficos como la profundidad (>100 m) y la pendiente del fondo (>10%), localizando los hábitat críticos para reproducción y cría dentro de la plataforma continental. Los planes y propuestas para un sistema de áreas marinas protegidas deben de considerar la conectividad de la isla del Caño y la Bahía de Drake, así como la extensión de los límites marinos del Parque Nacional Corcovado.
Palabras clave: Hábitat crítico, zonas de reproducción, zona de cría, Megaptera novaeangliae, área marina protegida.
References
Acevedo, R., L. Oviedo & N. Silva. 2003. Identification of key areas to conservation of misticetes cetaceans off Margarita island. Paper presented at the Simposio Extinción en Margarita. V Congreso Venezolano de Ecología. Isla de Margarita, Venezuela. [ Links ]
Acevedo, J., K. Rasmussen, F. Felix, C. Castro, M. Llano, E. Secchi, M.A. Saborio, A, Aguayo-Lobo, B. Haase, M. Scheidat, L. Dalla Rosa, C. Olavaria, P. Forestell, P. Acuña, G. Kaufman & L. Pastene. 2007. Migratory destination of humpback whales from the Magellan Strait feeding ground, Southeast Pacific. Mar. Mamm. Sci. 23: 453-463. [ Links ]
Caldwell, D.K., M.C. Caldwell & C.M. Walker. 1970. Marine mammals from Puerto Rico to Antigua. J. Mamm. 51: 636-639. [ Links ]
Corkeron, P.J. 1995. Humpback whales (Megaptera novaeangliae) in Hervey Bay, Queensland: Behaviour and responses to whalewatching vessels. Can. J. Zool. 73:1290-1299. [ Links ]
Compton R.C. 2004. Predicting key habitat and potential distribution of northern bottlenose whales (Hyperoodon ampullatus) in the northwest Atlantic ocean. MSc Thesis. University of Plymouth, Plymouth, U.K. [ Links ]
Frankel, A.S., C.W. Clark, L.M. Herman & C.M. Gabriele. 1995. Spatial distribution, habitat utilization, and social interactions of humpback whales, Megaptera novaeangliae, off Hawaii, determined using acoustic and visual techniques. Can. J. Zool. (73): 1134-1146. [ Links ]
Frantzis, A., O.Nikolaou, J.M. Bompar & A. Cammedda. 2004. Humpback whale (Megaptera novaeangliae) occurrence in the Mediterranean Sea. J. Cetacean Res. Manage. 6:25–38. [ Links ]
Félix, F. & B, Haase. 2001. The humpback whale off the coast of Ecuador, population parameters and behaviour. Rev. Biol. Mar. Oceanog. 36: 61 – 74. [ Links ]
Félix, F. & B, Haase. 2005. Distribution of humpback whales along the coast of Ecuador and management implications. J. Cetacean Res. Manage. 7:21–31. [ Links ]
Flores-González, L., J.J.Capella, B.Haase, V.A. Bravo, F. Félix & T. Gerrodette. 1995. Movements of southern hemisphere humpback whales within Eastern Tropical Pacific waters – winters 1986 – 1993. Eleventh Biennial Conference on the Biology of Marine Mammals, Orlando, Florida. [ Links ]
Flores-González, L., I.C. Avila, J.J. Capella, P. Falk, F. Felíx, J. Gibbons, H. Guzmán, B. Haase, J.C. Herrera, V. Peña, L. Santillan, I.C. Tobon & K. Van Warebeek. 2007. Estrategia para la conservación de la ballena jorobada del Pacífico Sudeste. Lineamientos de un plan de acción regional e iniciativas nacionales. Fundación Yubarta. Cali, Colombia. [ Links ]
Gibbons, J., J.J. Capella & C. Valladares. 2003. Rediscovery of a humpback whale, Megaptera novaeangliae, feeding ground in the Straits of Magellan, Chile. J. Cetacean Res. Manage. 5:203-08. [ Links ]
Grerg, E.J. & A. W. Trites. 2001. Prediction of critical habitat for five whales species in the waters of coastal British Columbia. Can. . Fish. Aq. Sci. 58: 1265–1285. [ Links ]
Hirzel, A.H., V. Helfer & F. Metral, F. 2001. Assessing habitat-suitability models with a virtual species. Ecol. Model. 44:111-121 [ Links ]
Hirzel, A.H., J. Hausser, D. Chessel & N. Perrin. 2002. Ecological-niche factor analysis: How to compute habitat-suitability maps without absence data?. Ecology 83:2027-2036 [ Links ]
Hirzel, A.H., B. Posser, P. A. Oggier, I. Crettenand, C. Glentz & R. Arlettaz. 2004. Ecological requirements of reintroduced species and the implication for release policy: the case of the bearded vulture. J. Applied Ecol. 41:1103-1116 [ Links ]
Hoyt E. 2005. Marine Protected Areas for whales, dolphins and porpoises: a world handbook for cetacean habitat conservation. Earthscan, London, U.K. [ Links ]
Kaschner, K. 2004. Modelling and mapping resource overlap between marine and fisheries on a global scale. PhD Thesis. University of British Columbia, British Columbia, Canada. [ Links ]
Mattila, D.K. & P.J. Clapham. 1989. Humpback whales, Megaptera novaeangliae, and other cetaceans on Virgin bank and in the northern Leeward Islands, 1985 and 1986. Can. J. Zool. : 2201-2211. [ Links ]
May-Collado, L., T. Gerrodette, J. Calambokidis, K. Rasmussen, & I. Sereg. 2005. Patterns of cetacean sighting distribution in the Pacific Exclusive Economic Zone of Costa Rica, based on data collected from 1979-2001. Rev. Biol. Trop. 53: 249-263. [ Links ]
Mignucci-Giannoni, A. 1998. Zoogeography of cetaceans off Puerto Rico and the Virgin islands. Carib. J. Sci. 34: 173–190. [ Links ]
Oviedo, L., N. Silva, L. Bermúdez & D.K. Odell, D.K. 2005. Distribution of bottlenose dolphins (Tursiops truncatus) on the east coast of isla Margarita and the Los Frailes Archipelago, Venezuela. Aquatic Mamm. 31: 442-446. [ Links ]
Rasmussen, K., D.M. Palacios, J. Calambokidis, M.T. Saborio, L. Dalla Rosa, E. Secchi, G. Steiger, J.M. Allen & G. Stone. 2007. Southern hemisphere humpback whales wintering off Central America: insight from water temperatura into the longest mammalian migration. Biol. Let. 3: 302-305. [ Links ]
Reeves, Randall R., B.D. Smith, E. Crespo & G. Notarbartolo di Sciara (compilers). 2003. Dolphins, Whales and Porpoises: 2002–2010 Conservation Action Plan for the Worlds Cetaceans. IUCN/SSC Cetacean Specialist Group. IUCN, Gland, Switzerland. [ Links ]
Scheidat, M., C. Castro, J. Gonzalez, & R. Williams. 2004. Behavioural responses of humpback whales (Megaptera novaeangliae) to whalewatching boats near isla de la Plata, Machalilla National Park, Ecuador. J. Cetacean Res. Manage. 6:63-8. [ Links ]
Stevick, P.T., A. Aguayo, J. Allen, I.C. Avila, J. Capella, C. Castro, K. Chater, L. Dalla Rosa, M.A. Engel, F. Félix, L. Flórez-González, A. Freitas, B. Haase, M. Llano, L. Lodi, E. Muñoz, C. Olavarría, E. Secchi, M. Scheidat & S. Salvatore. 2004. Migrations of individually identified humpback whales between the Antarctic Peninsula and South America. J. Cetacean Res. Manage. 6:109–113. [ Links ]
Whitehead, H. & M. Moore. 1982. Distribution and movements of West indian humpback whales in winter. Can. J. Zool. (60): 2203-2211. [ Links ]
Internet reference
Rasmussen, K., Calambokidis, J. and Steiger, G. 2004. Humpback whales and other marine mammals off Costa Rica and surrounding waters, 1996-2003. Report of the Oceanic Society 2003 field season in cooperation with Elderhostel volunteers. Seattle, USA. [Available on http://www.cascadiaresearch.org/reports/rep-cr04.pdf]. [ Links ]
Silva, N., R. Acevedo & L. Oviedo. 2006. Preliminary observations on the spatial distribution of humpback whales off the north coast of Margarita island, Venezuela -Southeast Caribbean. JMBA2 Biodiversity Records. [Available on http://www.mba.ac.uk/jmba/pdf/5224.pdf]. [ Links ]

