










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.55 n.3-4 San José Sep./Dec. 2007
Ecología poblacional del ratón Peromyscus mexicanus (Rodentia: Muridae) en el Parque Nacional Volcán Poás, Costa Rica
Licidia Rojas Rojas & Minor Barboza Rodríguez
1 Programa Regional de Posgrado en Biología, Sistema de Estudios de Posgrado, Universidad de Costa Rica, San José, Costa Rica; licidia.rojas@gmail.com, minorbr@costarricense.cr
Abstract: Population ecology of the mouse Peromyscus mexicanus (Rodentia: Muridae) in Poas Volcano National Park, Costa Rica. The Mexican Deer Mouse has been reported as an abundant wild mouse in Costa Rica; nevertheless, it has not been studied as well as other Peromyscus species. Thirty Sherman traps were placed on three habitats during six consecutive days of each month, from March 2002 through April 2003 in three sites of Poás volcano National Park, Costa Rica. A total of 2 393 mice were captured. Other species such as Reithrodontomys creper, R. rodriguezi, Scotinomys teguina and Oryzomys devius (Muridae) were also captured in Tierra Fría and R. creper, R. sumichrasti, S. teguina and O. devius in Potrero Grande. in Canto de las Aves we captured P. mexicanus, R. creper, R. rodriguezi and O. devius. Of the total mice collected, 34.77 % were P. mexicanus. For this species, the mean monthly capture per hectare was 34±2.15 in Tierra Fría and 11±1.85 in Potrero Grande. in the third site, Canto de las Aves, only four mice were captured throughout the study. The estimated population size did not change among months in Tierra Fría, but it did in Potrero Grande. No sex ratio variation was found in any habitat. In Potrero Grande, weight averages were 43.54±3.42 g for males and 42.08±3.4 5 g for females. Variation in population structure among habitats was not significant. The presence of oak trees (Quercus sp.) and the high understory density could explain the stability of the population in this area. Rev. Biol. Trop. 55 (3-4): 1037-1050. Epub 2007 December, 28.
Key words: Mexican deer mouse, Peromyscus mexicanus, population ecology, Poás volcano National Park, Costa Rica, density.
El género Peromyscus constituye el grupo más diverso y con distribución más amplia entre los mamíferos pequeños de Norteamérica (Chirhart et al. 2005) y Centroamérica (Reid 1997). Por esa razón, ha sido ampliamente utilizado en estudios de ecología, comportamiento, fisiología, sistemática y evolución (Chirhart et al. 2005). El ratón de patas blancas (Peromyscus mexicanus) es el miembro del género que presenta la distribución geográfica más amplia y se encuentra en los bosques de tierras medias y altas (600–3 000 msnm) desde México hasta el oeste de Panamá (Reid 1997, Mora 2000). Es un animal terrestre y nocturno que se alimenta de tallos semillas y frutas, así como de algunos insectos (Mora 2000). Según Wenny (2000), esta especie cumple un papel importante como depredador de semillas del bosque nuboso de Costa Rica.
El ratón de patas blancas se ha descrito como una de las especies de múridos silvestres más abundantes de Costa Rica. Aunque esta especie ha sido poco estudiada, en relación a otras especies del género Peromyscus, existen algunas investigaciones importantes en Costa Rica. Así, el ratón de patas blancas se ha descrito como una de las especies de múridos siltierras medias y altas (600–3 000 msnm) desde vestres más abundantes de este país (Anderson 1982, Van den Bergh y Kappelle 1998). De igual manera, otros estudios han determinado que P. mexicanus está asociado a zonas bosco sas, específicamente de crecimiento secundario (Johnson y Vaughan 1993, Van den Bergh y Kappelle 1998). El Parque Nacional volcán Poás (PNVP) es una de las áreas en el país donde se ha registrado alta abundancia de P. mexicanus (obs. pers.). Sin embargo, hasta la fecha no ha habido un estudio a largo plazo que documente estas observaciones.
En el presente estudio se evaluaron algunos aspectos de la ecología poblacional de P. mexicanus en el Parque Nacional Volcán Poás. Este trabajo permitió comparar variables como el tamaño de la población, la proporción de sexos y otros parámetros poblacionales con estudios realizados en otros sitios (Anderson 1982, Van den Bergh y Kappelle 1998). Además, se determinó la abundancia de esta especie en tres tipos de bosque dentro del parque. Por otro lado, se determinó el consumo relativo de algunos frutos silvestres propios de la zona, utilizando animales en cautiverio.
Materiales y métodos
Descripción del sitio de muestreo: el Parque Nacional Volcán Poás (PNVP) está ubicado en la Provincia de Alajuela y se encuentra en el extremo oeste de la Cordillera volcánica Central (latitud 10°0420" a 10º1633" N; longitud 84°1140" a 84º1607" W). El ámbito de temperatura osciló entre los 10 y 24 °C con un promedio de 14 °C y la precipitación anual varió entre 3 500 y 8 000 mm (Boza 2001). El clima del PNvP se caracteriza por tener una alta pluviosidad.
Para estudiar la ecología poblacional de P. mexicanus en ambientes distintos, se seleccionaron tres sitios de muestreo dentro del parque. Uno de ellos es conocido como Potrero Grande (Fig. 1). Esta es un área de pastizal que se formó hace unos 40 años, cuando se derribaron algunos parches de bosque para la creación de potreros, para el establecimiento de un hotel y un aserradero (M. Arias, com. pers.). Diez años después cesó la actividad antropogénica y comenzó un proceso de sucesión natural. Potrero Grande, se encuentra a una altitud de 2 500 msnm (Macey 1975). Algunas de las plantas más comunes de este sitio son: kikuyo (Pennisetum clandestinum), zacate (Paspalum boscianum), (Halenia rhyacophyla), el llantén (Plantago birtella), margarita amarilla (Hipochaerys radicata), juncia (Carex nogrum) y musgo de la turba (Sphagnum spp.). También es notable, la presencia de plantas introducidas como naranjo de monte (Hypericum strictum) y corona de Cristo (Ulex europaeaus) que han ido invadiendo la zona. En las orillas de la carretera que hay en esta zona, se pueden observar otras especie como escalonia (Escallonia poasana), lengua de vaca (Miconia myrtillifolia), arrayán (Vaccinium consanguineum) y bejuco muela (Pernettia coriacea).
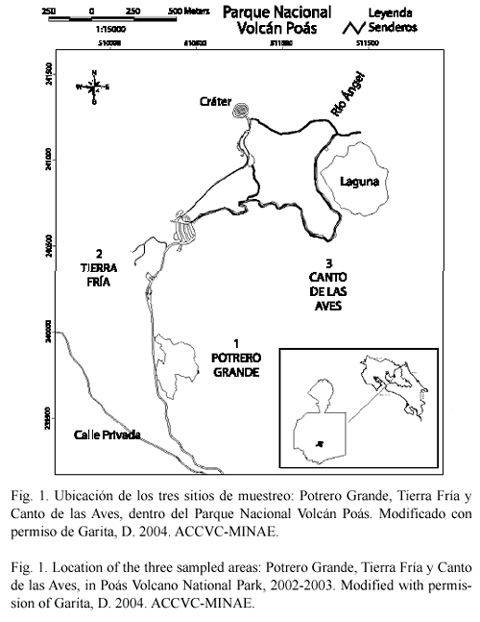
El segundo sitio de muestreo seleccionado se conoce como Tierra Fría. Este sitio es un bosque nuboso que está ubicado en el límite sur del parque (Fig. 1). Se encuentra a una altitud de entre 2 450 a 2 550 msnm (Macey 1975). En Tierra Fría, el dosel es abierto con pocos árboles grandes y un sotobosque relativamente denso. En este último se han desarrollado arbustos típicos de bosque secundario como lengua de vaca (Miconia spp.) y tocuico (Ardisia pleurobotrya), los cuales son más abundantes que las especies de dosel como el roble (Quercus spp.), papayillo (Schefflera rodriguesiana), fosforillo (Dendropanax sp.), escalonia (Escallonia myrilloides) y copey (Clusia odorata). En Tierra Fría se encuentra una gran variedad de plantas de crecimiento bajo, como carrizo (Chusquea spp.) y arete de india (Bomarea acutifolia).
El tercer sitio que se seleccionó es Canto de las Aves, el cual, se ubica al extremo este del cráter principal y es además el sitio de mayor elevación dentro de parque (Fig. 1). Canto de las Aves es un bosque nuboso de 2 600 a 2 700 msnm (Macey 1975). En este sitio las especies que dominan el dosel son copey (C. odorata), papayillo (S. rodriguesiana), cacho de venado (Dydimopanax pittieri), arrayán (Weinmannia trianea) y ratoncillo (Myrsine dependens), las cuales tienen unos 15 a 20 m de alto y con copas anchas por lo que la penetración de la luz es escasa en este sitio. Más abajo del dosel se pueden encontrar arbustos escasos de especies como tocuico (Ardisia pleurobotrya), arrayán (Vaccinium sp.) y miconia (Miconia biperulifera). También destacan algunas ericáceas epífitas como colmillo de perro (Cavendishia bracteata). Al acercarse a los bordes de la laguna, los carrizos (Chusquea spp.) se vuelven prominentes.
Determinación de algunos aspectos de la ecología poblacional: para determinar la ecología poblacional de P. mexicanus en cada uno de los tres sitios seleccionados, se realizaron muestreos mensuales mediante método de captura, marcado y liberación de animales. Para la captura de los ratones se colocaron 30 trampas Sherman (tamaño 23x9x8 cm) en cada sitio desde marzo de 2002 hasta abril de 2003. En cada sitio se colocaron tres filas (separadas por 10 m entre sí) de diez trampas cada una (espaciadas una de otra por 10 m). Se dejaron las trampas abiertas durante seis días consecutivos cada mes (excepto en los primeros dos meses del muestreo que se mantuvieron abiertas por cinco días). Cada día se revisaron las trampas y se les cambió el cebo. El cebo consistió en una mezcla de avena, mantequilla de maní y agua. Se marcó a cada individuo mediante un código de muescas en las orejas. Cada individuo capturado se identificó por marca, sexo y peso. El peso de determinó utilizando una balanza de mano de 100 g. La clasificación de individuos adultos e inmaduros se realizó siguiendo el protocolo de Anderson (1982).
Fluctuación en el tamaño poblacional de otras especies: también se determinó la importancia de P. mexicanus en la comunidad de múridos en cada sitio. Para ello, se registraron las capturas de las otras especies de múridos y se marcaron, se anotó el sexo y peso de cada individuo.
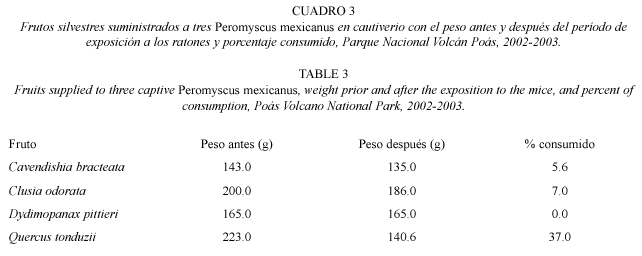
Proporción de frutos consumidos por individuos de P. mexicanus en cautiverio: para determinar el consumo relativo de algunos frutos silvestres en la dieta de P. mexicanus, se mantuvo a tres individuos en cautiverio durante diez noches consecutivas. Cada noche se colocó el mismo peso de los frutos de las cuatro especies más comunes en el sitio Tierra Fría; estas especies son roble (Quercus tonduzii.), colmillo de perro (Cavendishia bracteata), copey (Clusia odorata) y cacho de venado (Dydimopanax pittieri). Se tomó el peso de los frutos que se ofreció cada noche y en la mañana siguiente, por diferencia de peso se obtuvo el porcentaje de consumo de cada tipo de fruto. Cada mañana se verificó que la diferencia fuera producto del consumo (mordiscos) del ratón de patas blancas y no pérdidas por otros efectos.
Análisis estadístico: para estimar el tamaño de la población, el reclutamiento y la probabilidad de supervivencia relativa de la población de P. mexicanus, se utilizó el modelo de Jolli-Seber (Zar 1996). Se analizaron los datos con el Programa "Ecological Methodology" (versión 5.2. Krebs 2000). Cabe mencionar que al utilizar este modelo, se excluyen el primero y los dos últimos meses de muestreo por efectos de la técnica de captura-recaptura en los que se basa el mismo. Además, se usó el programa "Systat" (versión 9. SPSS 1998) para realizar todas las pruebas estadísticas. Para comparar la densidad entre sitios (excepto en Canto de las Aves donde no se obtuvieron suficientes datos), el tamaño de la población a través del tiempo y las proporciones de sexo, se utilizó la prueba de Chi cuadrado. Para comparar la diferencia en el peso corporal entre machos y hembras se utilizó una prueba U de Mann-Whitney y para comparar la variación en el peso corporal a través del tiempo se usó una prueba de Kruskal-Wallis. Para determinar las diferencias del tamaño poblacional de los individuos adultos con respecto a los inmaduros a través del tiempo se utilizó una prueba de Kolmogorov-Smirnov. Esta última prueba se utilizó además, para analizar las diferencias entre el tamaño poblacional mensual de P. mexicanus con respecto al tamaño poblacional mensual de otras especies. También se utilizó una correlación de Pearson para examinar la relación entre la precipitación mensual (datos de la estación meteorológica del ICE en el PNVP) y el tamaño poblacional de P. mexicanus. Para analizar la relación entre la fluctuación poblacional de P. mexicanus y R. creper en Tierra Fría se aplicó una correlación gradual (Spiegel 1971), este análisis sólo se llevó a cabo con estas dos especies y en este sitio, porque fue el único caso donde los datos fueron suficientes. Finalmente, se comparó el número de capturas de P. mexicanus cerca (de 0 de 5 metros) del borde del bosque con las capturas lejos (más de 25 metros) del borde del bosque en Potrero Grande, con una prueba de Chi cuadrado.
Resultados
Riqueza de especies: en todo el periodo de muestreo se hicieron 2 393 capturas, de las cuales 832 (34.77 %) correspondieron a P. mexicanus. En Tierra Fría, se capturaron cinco especies de múridos. En Potrero Grande también se capturaron cinco especies de la familia Muridae. En Canto de las Aves se capturaron cuatro especies de múridos (Cuadro 1).
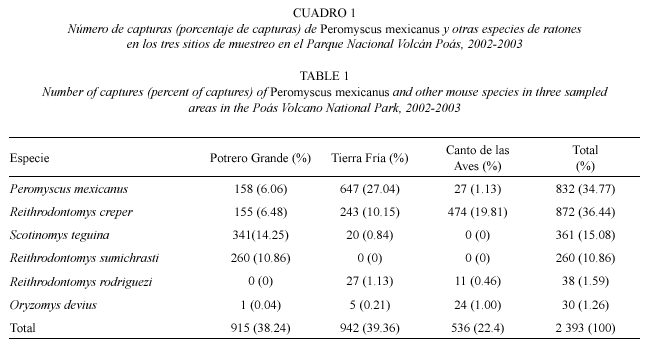
Densidad: La densidad de P. mexicanus mostró variación entre los tres sitios de estudio. En Tierra Fría se capturaron, en promedio 34±2.15 individuos por mes por hectárea, en Potrero Grande se capturaron 11±1.85 individuos por mes por hectárea y en Canto de las Aves sólo se capturaron cuatro individuos (dos machos y dos hembras) en febrero a abril de 2003. La densidad de P. mexicanus fue mayor en Tierra Fría que en Potrero Grande (x2=8.95; gl=1; p=0.003).
Efecto de la cercanía del bosque: en Potrero Grande se observaron más capturas de P. mexicanus cerca del bosque (139 capturas), que lejos de este (16 capturas), a pesar de que sólo nueve trampas (de un total de 30) se encontraban cerca del bosque (x2=260.03; gl=1; p<0.001).
Tamaño poblacional: el tamaño poblacional estimado de P. mexicanus no varió a lo largo del tiempo en Tierra Fría (x2=14.56; gl=11; p=0.20) (Fig. 2). En Potrero Grande se encontraron diferencias en el tamaño estimado de la población a través del tiempo (x2=34.21; gl=10; p<0.001); donde agosto y octubre presentaron el mayor aumento de la abundancia. Además, se obtuvo una correlación positiva entre la distribución de los promedios mensuales de lluvia y la distribución del tamaño poblacional mensual en Potrero Grande (r2=0.689; p=0.028). En Tierra Fría la relación entre la precipitación y el número de individuos también fue positiva (r2=0.565; p=0.070) (Fig. 2).
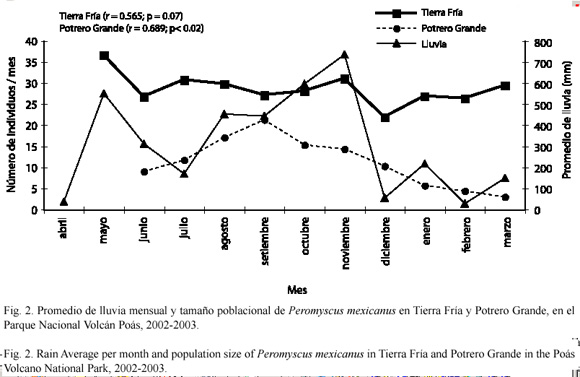
Índices de reclutamiento y supervivencia: en Tierra Fría se observó un mayor reclutamiento de P. mexicanus en mayo a julio, con un segundo aumento en octubre y noviembre. En Potrero Grande se encontraron los valores más altos de reclutamiento en julio y setiembre, sin embargo, en este caso los valores de los índices fueron variables (Cuadro 2). En resumen la tasa de reclutamiento en Tierra Fría es mayor que en Potrero Grande.
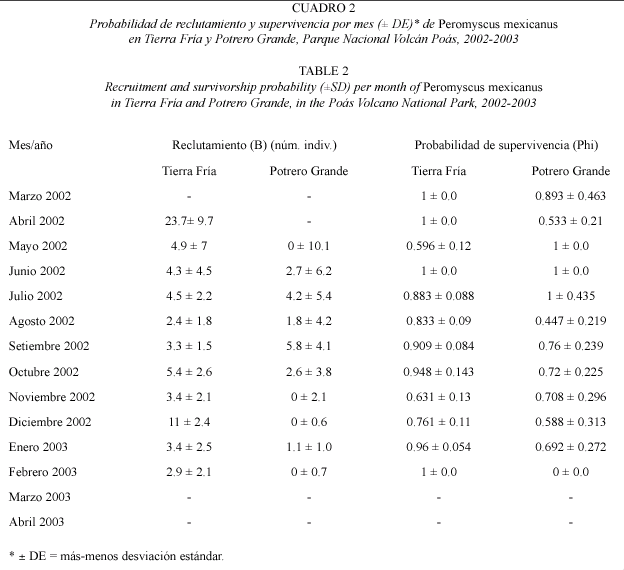
En general, la probabilidad de supervivencia de P. mexicanus en Tierra Fría varió entre 0.596 y 1. En Potrero Grande, los valores del índice de supervivencia presentaron variación en el tiempo, con un ámbito que varió desde 0.447 a 1 (Cuadro 2). Es importante señalar que los primeros y últimos meses del análisis presentan valores extremos debido a efectos propios del modelo de Jolli-Seber (Zar 1996).
Proporción de sexos: la proporción de sexos (machos: hembras) de P. mexicanus en Tierra Fría fue de 0.98:1 (41 machos y 42 hembras) y en Potrero Grande fue de 0.76:1 (13 machos y 17 hembras). La proporción de machos y hembras en cada sitio fue igual (Tierra Fría: x2=0; gl=1; p=1; Potrero Grande: x2=0.30; gl=1; p=0.58). También fueron iguales las proporciones de machos y hembras entre los dos sitios (x2=0.22; gl=1; p=0.64).
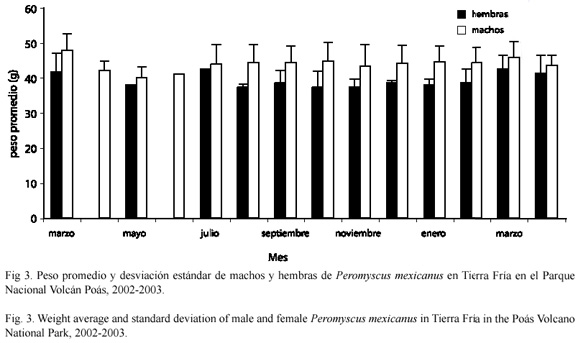
Distribución de el peso corporal: el promedio y la desviación estándar del peso de P. mexicanus en Tierra Fría para los machos fue 43.83+1.93 g y para la hembras fue 39.29+2.07 g. Existe diferencia en el peso promedio mensual entre machos y hembras para todos l estudio (U=1 213.5; gl=1; p<0.001). Además, el peso promedio mensual no varió a lo largo del tiempo ni en las hembras (H = 18.42; gl = 11; p = 0.07), ni en los machos (H = 8.06; gl = 13; p = 0.84) (Fig. 3).
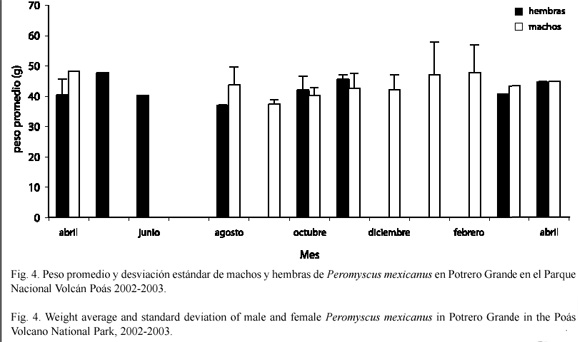
El promedio y la desviación estándar del peso de P. mexicanus en Potrero Grande para los machos fue de 43.54+3.42 g y para las hembras fue de 42.08+3.45 g. No hubo diferencias en el peso promedio mensual de los machos y las hembras (U=263.5; gl=1; p=0.16), pero esto puede deberse al bajo número de capturas que se obtuvo en algunos meses en este sitio. La distribución mensual del peso de las hembras fue igual a través del tiempo (Kruskal-Wallis=11.29; gl=9; p=0.25), lo mismo se observó para los machos (KruskalWallis= 10.68; gl=10; p=0.38) (Fig. 4).
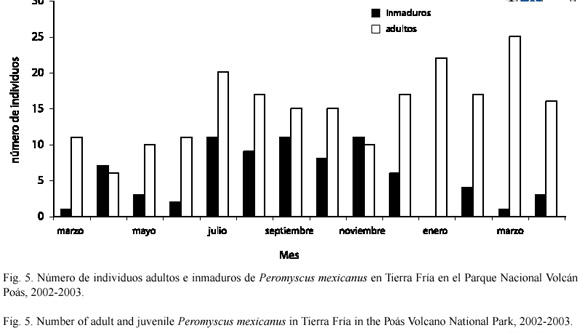
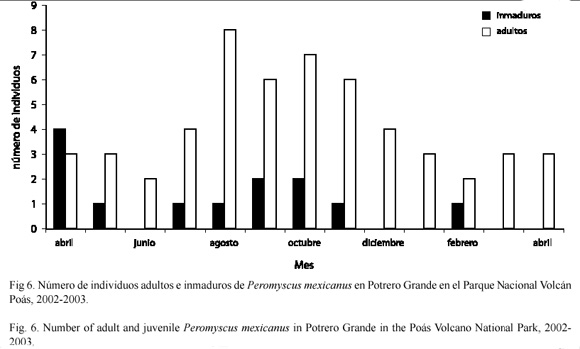
Distribución de individuos adultos e inmaduros: en Tierra Fría la distribución de los individuos adultos con respecto a la distribución de los inmaduros, fue diferente a través del tiempo (D=0.643; p=0.002) (Fig. 5). En general la cantidad de adultos fue mayor que la de inmaduros a través de este período (U=180.5; gl=1; p<0.001). Sin embargo, no se encontraron diferencias en la cantidad de inmaduros a través del tiempo (x2=3.81; gl=13; p=0.99). En Potrero Grande la distribución de adultos e inmaduros fue comparable a la observada en Tierra Fría (Fig. 6). La distribución de individuos adultos con respecto a la distribución de los inmaduros varió a través del tiempo (D=0.769; p<0.001). En general la cantidad de adultos fue mayor que la de inmaduros a través del período de muestreo (U=159.0; gl=1; p<0.001). La cantidad de inmaduros fue igual a través del tiempo (x2=9; gl=12; p=0.703).
Fluctuación en el tamaño poblacional de otras especies: en Tierra Fría la única especie adicional capturada todos los meses fue R. creper y la fluctuación poblacional de esta especie a través del tiempo difirió de la de P. mexicanus (D=0.917; p<0.001) (Fig. 7). No se encontró relación entre la variación del tamaño poblacional de una especie con respecto a la otra a través del tiempo (r=0.16; p=0.66).
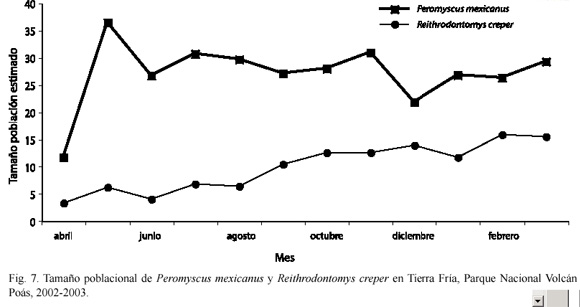
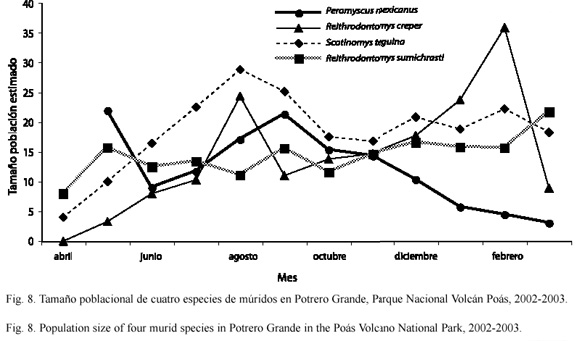
En Potrero Grande no hubo diferencias en la fluctuación poblacional de P. mexicanus con respecto a la de Reithrodontomys sumichrasti a través del tiempo (D=0.455; p=0.185), ni a la de Reithrodontomys creper (D=0.279; p=0.768), pero si hubo diferencias con Scotinomys teguina (D=0.636; p=0.017) (Fig. 8).
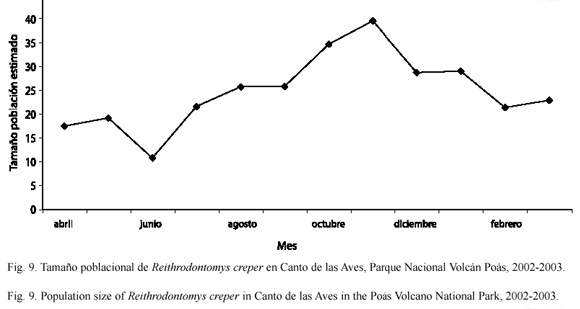
En el sitio Canto de las Aves, la abundancia de R. creper fue mayor que en Tierra Fría (U=138; gl=1; p<0.001) (Fig. 9). Ya que sólo se capturaron cuatro individuos de P. mexicanus, no fue posible establecer ninguna relación respecto a las fluctuaciones en el número de individuos entre estas dos especies.
Proporción de frutos consumidos por individuos de P. mexicanus en cautiverio: al analizar el porcentaje de frutos silvestres consumidos se observó que P. mexicanus prefirió consumir el fruto de roble (Quercus tonduzii) (x2=165.7; gl=3; p<0.05) (Cuadro 3). Estos resultados indican que el consumo de frutos de roble representó un 85 % del peso consumido durante los días de observación. Los restantes frutos silvestres incluidos en el análisis no fueron consumidos o se consumieron en proporciones muy bajas. Por ejemplo de los frutos de copey (Clusia odorata) los ratones sólo consumieron los arilos en ciertas ocasiones.
Discusión
Abundancia: Peromyscus mexicanus fue una de las dos especies que se capturaron con mayor frecuencia (Cuadro 1). En otras zonas de Costa Rica, como Monteverde y Talamanca, P. mexicanus ha sido reportado como la especie de múrido más abundante (Anderson 1982, Johnson y vaughan 1993, Van den Bergh y Kapelle 1996, McCain 2004). Otros representantes del género Peromyscus, también han presentado una mayor abundancia con respecto a otras especies de múridos, en diferentes sitios estudiados (Anthony y Niles 1981, Horváth et al. 2001).
En Tierra Fría y en Potrero Grande las dos especies son capaces de coexistir con poblaciones relativamente abundantes, esto podría explicarse gracias a la diferenciación morfológica que presentan estos dos ratones (Reid 1997), lo cual les podría conferir diferencias en sus hábitos de forrajeo (Smartt 1978). En Potrero Grande, en la mayor parte del tiempo se observa que cuando una de estas especies aumenta su tamaño poblacional, la otra disminuye, en Tierra Fría este patrón no es claro (Fig. 7 y 8). Hace falta más investigación para dilucidar los factores que regulan el tamaño poblacional de estas especies en estos sitios. Ya que otros factores ambientales pueden contribuir a modular la interacción entre ambas especies, favoreciendo el aumento en el número de individuos de una especie a expensas de una disminución en el número de individuos de la otra especie.
Los mayores tamaños poblacionales de P. mexicanus se observaron en Tierra Fría. Este resultado puede ser explicado con varios argumentos. Primero, en dicho sitio de estudio es el único en el que hay roble (Quercus tonduzii). Según los resultados de este estudio (Cuadro 2) los frutos de roble fueron proporcionalmente más consumidos por el ratón de patas blancas que los otros frutos estudiados, por lo que la presencia de roble puede ser importante para la abundancia del ratón patas blancas en Tierra Fría. Además, en Tierra Fría existe una mayor densidad de sotobosque que en los otros sitios estudiados (Macey 1975, obs. pers.) por lo que P. mexicanus puede encontrar mejores condiciones para su supervivencia y reproducción (Johnson y vaughan 1993, Van den Bergh y Kappelle 1998). Por ejemplo, MCloskey (1976), encontró que el aumento en el volumen de arbustos, mejora la actividad de forrajeo, movimiento, escape a depredadores y, en algunos casos, la construcción de madrigueras en especies como P. eremicus, P. californicus, P. megalotis.
En Tierra Fría P. mexicanus no mostró variaciones en el tamaño poblacional y presentó una supervivencia alta a través del tiempo. Este resultado es contrario a lo observado en otros estudios con Peromyscus. En sitios donde la estacionalidad es marcada, las variaciones en el tamaño poblacional a través del tiempo han sido correlacionadas con las fluctuaciones en la disponibilidad de alimento en algunas épocas del año (Fleming 1974, MCloskey 1976, Lewellen y Vessey 1998, Lamberth y Adler 2000). En Tierra Fría se observaron potenciales fuentes de alimento para P. mexicanus durante todo el año, particularmente frutos de roble en el suelo (obs. pers.). Estas observaciones, aunadas a la estabilidad en el tamaño poblacional y la alta tasa de supervivencia de P. mexicanus en Tierra Fría, sugieren que en este sitio la disponibilidad de alimento se mantiene relativamente estable a lo largo del año.
En Potrero Grande se observó una fluctuación en el número de individuos y en la supervivencia de P. mexicanus a través del tiempo. Además, el aumento en la población de P. mexicanus en algunos meses en este sitio, se correlacionó positivamente con el aumento en las lluvias. Potrero Grande es un sitio con menor diversidad de especies vegetales que Tierra Fría y donde no se encontraron frutos de roble (Quercus spp.). Según estos datos, este sitio podría resultar menos estable en cuanto a la disponibilidad de recursos alimentarios y espaciales para esta especie. Esto es particularmente importante sobre todo durante los meses secos, pues se sabe que el período lluvioso propicia la producción y caída de frutos y semillas (Lamberth y Adler 2000). Estos factores y el hecho de que se capturaron a P. mexicanus principalmente en las trampas cercanas al bosque, sugieren que esta especie es oportunista en Potrero Grande, movilizándose a esta zona en los meses más favorables desde el bosque cercano.
Distribución de individuos adultos e inmaduros: en Tierra Fría el reclutamiento de P. mexicanus fue relativamente constante en todo el período de muestreo, mientras que en Potrero Grande los valores de reclutamiento variaron considerablemente (Cuadro 2). A su vez, se observó la presencia de inmaduros en todo el período de muestreo en Tierra Fría y en algunos meses en Potrero Grande, particularmente entre julio a noviembre. Los datos obtenidos en este estudio sobre reclutamiento y número de adultos e inmaduros en Tierra Fría parecen apoyar lo observado por Heideman y Bronson (1993). Estos autores encontraron que P. mexicanus es oportunista en su reproducción. Según ellos, en sitios o años favorables, el ratón de patas blancas es capaz de madurar rápidamente y reproducirse con mayor frecuencia y no estacionalmente.
Proporción de sexos: no se observaron diferencias en la proporción de sexos de P. mexicanus en el Parque Nacional volcán Poás. Este resultado puede sugerir que no ha habido una presión selectiva que promueva la producción de un sexo más que el otro en esta población. Havelka y Millar (1997) han sugerido que las condiciones ambientales pueden alterar la proporción de sexos en Peromyscus, de acuerdo con las diferencias en las respuestas fisiológicas de los machos y las hembras, o por la inhabilidad de uno de los dos sexos en establecerse o adquirir los recursos según las condiciones climáticas. Asimismo, sugieren que en zonas templadas se presenta variación en la proporción de sexos debido a las limitaciones ambientales presentes en estos sitios, las cuales favorecen la acción de la selección natural hacia la mayor producción de uno de los sexos.
Distribución del peso corporal: el mayor peso de los machos con respecto a las hembras en Tierra Fría (Fig. 3) concuerda con los datos de Van den Bergh y Kapelle (1996), en San Gerardo de Dota, Costa Rica. En Potrero Grande no se observó este mismo resultado, posiblemente por el tamaño de la muestra, que fue relativamente bajo en este sitio.
No se encontraron variaciones en el peso dentro de los sexos a través del periodo de muestreo en Tierra Fría, ni en Potrero Grande. En un estudio realizado por Heideman y Bronson (1992) en Monteverde (Costa Rica), se encontró una reducción del peso corporal en la estación seca. Según los autores mencionados, esto es un efecto de la escasa disponibilidad de recursos en los meses secos. El ratón de patas blancas no mostró esta variación en Tierra Fría, posiblemente porque los recursos ahí parecen ser relativamente constantes (obs. pers.). Aunque en Potrero Grande al parecer, no hay refugio y alimento permanente, los ratones tienen la posibilidad de utilizar los bosques aledaños en los tiempos de escasez (obs. pers.).
En conclusión, P. mexicanus fue una de las especies que se capturó con mayor frecuencia en el Parque Nacional volcán Poás y Tierra Fría fue el sitio donde se observó mayor estabilidad de la población en términos de su tamaño poblacional, fluctuación poblacional a través del tiempo, reclutamiento, supervivencia y variación en el peso corporal. Así, es posible que la presencia de roble (Quercus sp.) y de una alta cantidad de refugios en el sotobosque en Tierra Fría, sean factores fundamentales en la estabilidad de la población. Por otro lado, la utilización estacional de Potrero Grande, un sitio con variaciones en la disponibilidad de recursos a través del tiempo, apoya la idea de que esta especie es oportunista.
Agradecimientos
A José Manuel Mora, Bernal Morera y Jorge Azofeifa; además a María Eugenia Zaldívar, Oscar Rocha, Alejandro Alvarado, Alexander García. A la Escuela de Biología y al Sistema de Estudios de Posgrado de la Universidad de Costa Rica por el equipo y financiamiento facilitado para el proyecto. A IDEA WILD por las trampas donadas para este proyecto. Finalmente al MINAE y al Instituto Costarricense de Electricidad (ICE).
Resumen
Se estudió el tamaño poblacional, la proporción de sexos, el peso y el número de adultos e inmaduros de Peromyscus mexicanus en tres sitios, durante 14 meses, en el Parque Nacional volcán Poás, Costa Rica. Se colocaron 30 trampas Sherman en cada sitio, para un estudio de captura-recaptura durante seis días consecutivos de cada mes, desde marzo de 2002 hasta abril de 2003. Se hicieron 2 393 capturas, hallándose en Tierra Fría, además de P. mexicanus, Reithrodontomys creper, R. rodriguezi, Scotinomys teguina y Oryzomys devius (Muridae). En Potrero Grande se capturaron P. mexicanus, R. creper, R. sumichrasti, S. teguina y O. devius. En Canto de las Aves se capturaron P. mexicanus, R. creper, R. rodriguezi y O. devius. El 34.77 % del total de ratones capturados fueron P. mexicanus. Se capturaron en promedio 34 individuos por mes por hectárea en Tierra Fría y 11 en Potrero Grande; en Canto de las Aves sólo se capturaron 4 individuos en todo el muestreo. El tamaño estimado de la población de P. mexicanus no varió mensualmente en Tierra Fría, pero si en Potrero Grande. No se observó variación en la proporción de sexos en ninguno de los dos sitios. El valor promedio de peso en Tierra Fría fue de 43.83+1.93 g machos 39.29+2.07 g hembras. En Potrero Grande fue 43.54+3.42 g machos y 42.08+3.45 g hembras. En ninguno de los dos sitios se encontraron diferencias en la cantidad de individuos inmaduros a través del tiempo.
Palabras clave: ratón de patas blancas, Peromyscus mexicanus, ecología poblacional, Parque Nacional volcán Poás, Costa Rica.
Referencias
Anderson, S.D. 1982. Comparative population ecology of Peromyscus mexicanus in a Costa Rican wet forest. Tesis de Doctorado, University of Southern California, Los Angeles, California, EEUU. [ Links ]
Anthony, R. & L. Niles. 1981. Small mammal associations in forested and old field habitats, a quantitative comparison. Ecology 62: 955-963. [ Links ]
Boza, M. 2001. Parques Nacionales de Costa Rica. INCAFO San José, San José, Costa Rica. [ Links ]
Chirhart, S., R. Honeycutt & I. Greenbaum.2005. Microsatellite variation and evolution in the Peromyscus maniculatus species group. Mol. Phylog. and Evol. 34: 408-415. [ Links ]
Fleming, T.H. 1974. The population ecology of two species of Costa Rican Heteromyd rodents. Ecology 55: 493-510. [ Links ]
Havelka, M. & J. Millar. 1997. Sex ratio of offspring in Peromyscus maniculatus borealis. J. Mammal. 78: 626-637. [ Links ]
Heideman, P. & F. Bronson. 1992. A pseudoseasonal reproductive strategy in a tropical rodent, Peromyscus nudipes. J. Repr. Fert. 95: 57-67. [ Links ]
Heideman, P. & F. Bronson. 1993. Potential and realized reproduction in a tropical population of Peromyscus (Rodentia). J. Mammal. 74: 261-269. [ Links ]
Horváth, A., I. March & J. Wolf. 2001. Rodent diversity and land use in Montebello, Chiapas, México. Stud. neotrop fauna & environm. 36: 169-176. [ Links ]
Johnson, W. & C. Vaughan. 1993. Habitat use of small terrestrial rodents in the Costa Rican highlands. Rev. Biol. Trop. 41: 521-527. [ Links ]
Lambert, T. & G. Adler. 2000. Microhabitat use by a tropical forest rodent, Proechimys semispinosus, in Central Panama. J. Mammal. 81: 70-76. [ Links ]
Lewellen, R.H. & S.H. Vessey. 1998. Modeling biotic and abiotic influences on population size in small mammals. Oecologia 113: 210-218. [ Links ]
Macey, A. 1975. The vegetation of volcán Poás National Park, Costa Rica. Rev. Biol. Trop. 23: 239-255. [ Links ]
McCain, C. 2004. The mid-domain effect applied to elevational gradients: Species richness of small mammals of Costa Rica. J. Biogeogr. 31: 19-31. [ Links ]
M`Closkey, R. 1976. Community structure in sympatric rodents. Ecology 57: 728-739. [ Links ]
Mora, J.M. 2000. Los mamíferos silvestres de Costa Rica. EUNED San José, San José, Costa Rica. [ Links ]
Reid, F. 1997. A field guide to the mammals of Central America y Southeast Mexico. Oxford University, Nueva York, EEUU. [ Links ]
Smartt, R. 1978. A comparison of ecological and morphological overlap in a Peromyscus community. Ecology 59: 216-220. [ Links ]
Spiegel, M.R. 1971. Estadística. McGraw Hill, México D.F., México. [ Links ]
Van den Bergh, M.B. & M. Kapelle. 1998. Diversity and distribution of small terrestrial rodents along a disturbance gradient in montane Costa Rica. Rev. Biol. Trop. 46: 331-338. [ Links ]
Wenny, D. 2000. Seed dispersal, seed predation, and seedling recruitment of a neotropical montane tree. Ecol. Monogr. 70: 331-345. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. Prentice Hall, Nueva York, EEUU. [ Links ]

