










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 suppl.2 San José Dec. 2006
Ultraestructura de bambúes del género Dendrocalamus (Poaceae: Bambusoideae) cultivados en Costa Rica IV: Dendrocalamus asper, clones Taiwán y Tailandia
Mayra Montiel1,2 & Ethel Sánchez2
1 Facultad de Ciencias Agroalimentarias, Escuela de Zootecnia, Universidad de Costa Rica. Ciudad Universitaria Rodrigo Facio, San Pedro de Montes de Oca. Apdo. Postal 2060, San José, Costa Rica; mmontiel@cariari.ucr.ac.cr
2 Centro de Investigación en Estructuras Microscópicas (CIEMIC), Universidad de Costa Rica. Apdo. Postal 2060, San José, Costa Rica. Fax: (506) 207-3182; ethels@cariari.ucr.ac.cr
Recibido 15-VI-2006. Corregido 15-VII-2006. Aceptado 20-VIII-2006.
Abstract: An ultraestructural analysis of the Taiwan and Thailand clones of Dendrocalamus asper made with the scanning electron microscope showed that the Taiwan clone had low dome stomata while the Thailand clone has high dome stomata in the leaf lamina. Cuticular wax accumulations in the Taiwan clone has two shapes, against four in the Thailand clone. Both have the same distribution of vascular and surrounding tissues. Macroscopic or microscopic observation of culm bracts are not recommended to differentiate these clones: a reliable identification requires observation of the ultrastructure of stomata and cuticular wax accumulations. Rev. Biol. Trop. 54(Suppl. 2): 65-75. Epub 2006 Dec. 01.
Key words: ultraestructure, scanning electron microscopy, anatomy, bamboo, Poaceae, Dendrocalamus asper, Taiwan clon, Thailand clon.
Las especies de bambú son muy importantes en el ámbito mundial por la variedad de sus productos. Dentro de éstas, el género Dendrocalamus es uno de los más relevantes por sus numerosos usos en la construcción,la indus- tria farmacéutica,en la producción de muebles, industrial del papel, artesanías y otros.
El género Dendrocalamus está compuesto por 27 especies en el subcontinente indio (Seethalakshmi y Kumar 1998) y dos especies más en el sureste de Asia y en la Península de Malasia. Se encuentra en altitudes de 0-1 500 msnm y se adapta a suelos secos y húmedos, creciendo mejor en suelos ricos; además tolera bajas temperaturas de hasta -33ºC.
Dentro de las especies de este género, Dendrocalamus asper (Schultes f.) se destaca por la excelente calidad y resistencia de su "madera", así como por el delicado sabor de sus retoños y raíces, que constituyen un platillo delicioso.
D. asper se encuentra en Asia tropical y ha sido introducido a varios países tropicales de América, así como en las estaciones experimentales de la Universidad de Costa Rica, jardines botánicos y fincas privadas en Costa Rica. El origen de esta especie puede ser el sureste asiático.
D. asper es también conocido como Bambusa aspera Schultes f.(1830), Gigantochloa aspera (Schultes f.) Kurz (1876), Dendrocalamus flagellifer y Dendrocalamus marrillianus Elmer (1915).
Dentro de los nombres comunes están "bambú gigante", "bambú betung", "phai-tong" en Indonesia, "sweet bambú" en Tailandia y "manh-tong" en vietnam (Widjaja 1995).
Respecto a la identificación de los bambúes, esta es un poco diferente a la que se realiza para la mayoría de las plantas con flores, debido a que el proceso de floración tarda muchos años y suele ser esporádico. Por esta razón, se identifican géneros y especies mediante sus características anatómicas. Las brácteas que recubren el culmo son un instrumento importante y de acuerdo con la especie presentan formas y estructuras diferentes entre sí. Pueden ser glabras o pubescentes y son únicas para cada especie. Esto, aunado al conocimiento de la anatomía de la epidermis de la lámina foliar, los tipos de tricomas, papilas céricas y células de sílice, ayuda a establecer diferencias significativas para cada especie, variedad o clon (Ghosh y Negi 1959, Tateoka et al. 1959, Metcalfe 1960, Grosser y Liese 1971).
Para contribuir a los conocimientos taxonómicos de D. asper, el objetivo de este trabajo fue analizar ultraestructuralmente los clones Taiwán y Tailandia de esta especie.
Materiales y métodos
Se recolectaron muestras de culmo, hojas, rama, y brácteas del culmo de D.asper, clon Taiwán, de dos años de edad en la finca Río Frío, Heredia, Costa Rica, a 100 msnm. En este sitio el suelo es tipo areno-limoso y la precipitación anual es de 4 120 mm, la temperatura promedio es de 25 ºC y la humedad relativa de 85-95%.
Los especímenes de dos años de edad del clon de Tailandia se recolectaron en Pedregocito de Pedregal, San Isidro del General, San José, Costa Rica, a 400 msnm. Esta zona posee un suelo tipo areno-limoso y la precipitación anual es de 2 050 mm, la temperatura promedio es de 27 °C y la humedad relativa de 80-85%.
Las muestras fueron fijadas en una solución de glutaraldehído (2.5%) y paraformaldehído (2%) en amortiguador de fosfato de sodio (0.1 M, pH 7.4), por 24 horas a 4°C. Luego se lavaron con el amortiguador y se posfijaron con tetraóxido de osmio (2%), por una hora. Posteriormente se lavaron tres veces con agua destilada, se deshidrataron en un gradiente ascendente de etanol (30-50%) y se secaron utilizando terbutanol en un secador por sublimación. Una vez secas, se montaron en bases de aluminio y utilizando un cobertor iónico (Eiko I-D 2) se cubrieron con 30 nm de oro, y se observaron con un microscopio electrónico de barrido (Hitachi S-570).
Resultados
Clon Taiwán
Lámina foliar: La superficie adaxial (haz) de la lámina foliar tiene la nervadura central o zona costal que corresponde al haz vascular de esa nervadura, así como las zonas intercostales (laterales a ella). A mayor magnificación se observa el detalle de las células reticuladas de la zona costal. En la zona intercostal hay estomas, células en forma de gancho, tricomas unicelulares, tricomas bicelulares, células de sílice y puntos blancos que corresponden a las papilas céricas (Fig. 1A, 1B). En la figura 1C se pueden observar estomas y dos cordones de células buliformes.
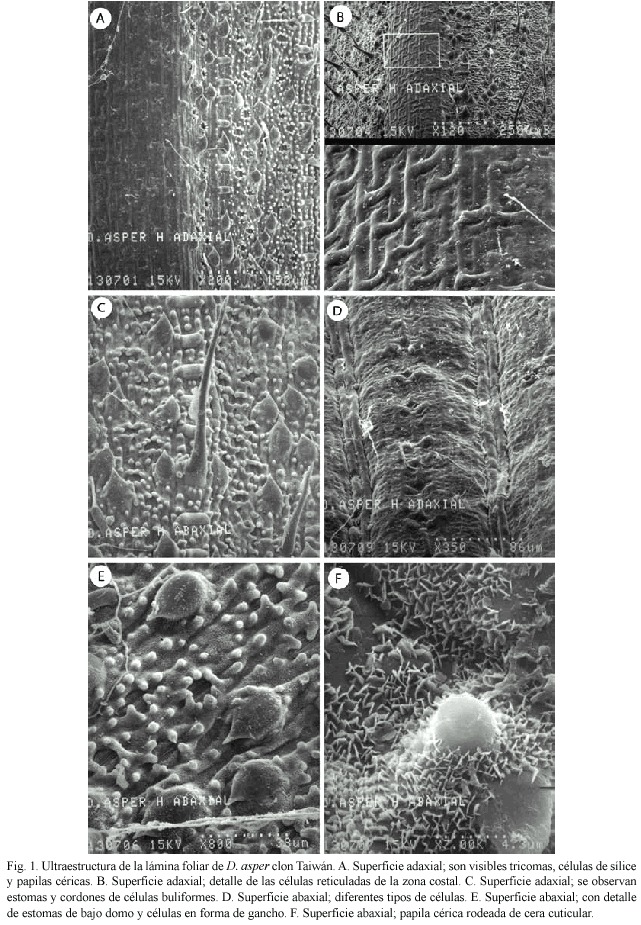
En la superficie abaxial (envés) (Fig. 1D), observamos células en forma de gancho, estomas, tricomas unicelulares largos, células de sílice y papilas céricas, como se observa un patrón muy similar al de la zona adaxial. En la figura 1E, a alta magnificación se observan con más detalle los estomas de bajo domo, las células en forma de gancho y el patrón de distribución de la cera cuticular. En la figura 1F, una papila cérica y cera cuticular a su alrededor.
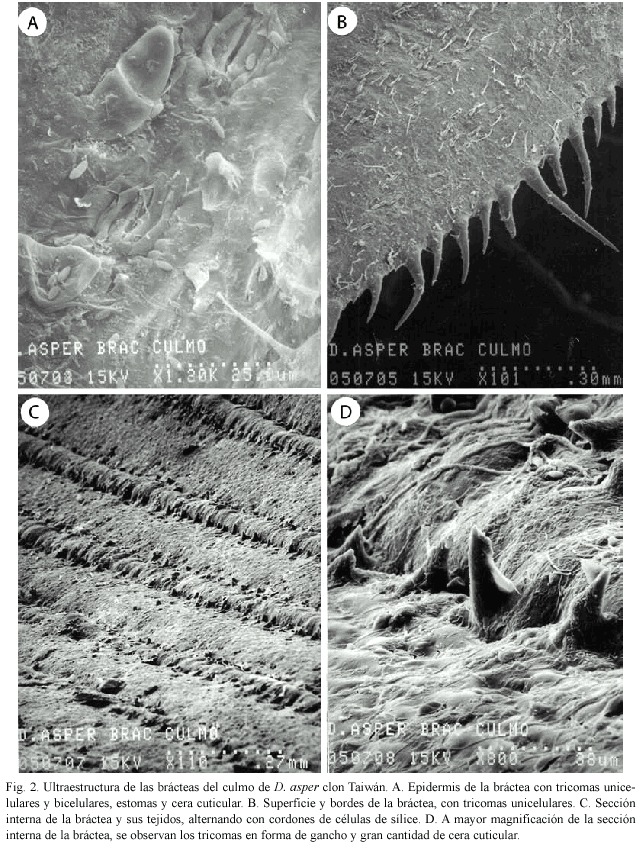
Bráctea del culmo: La epidermis de la bráctea presenta tricomas pequeños bicelulares, estomas, cera cuticular y tricomas unicelulares (Fig. 2A). Los bordes y la superficie de las brácteas presentan tricomas unicelulares de varios tamaños, lo que le da una sensación áspera al tacto (Fig. 2B). La parte interna de la bráctea (Fig. 2C) tiene filas de células de sílice que alternan con otros tejidos. A mayor magnificación (Fig. 2D) se pueden apreciar tricomas en forma de gancho y grandes cantidades de cera cuticular.
En los bordes de las brácteas también se presentan estructuras similares a "aurículas" con tricomas unicelulares muy largos (Figs. 3A, 3B). La figura 3C tiene el inicio o unión de la bráctea con el nudo del culmo, y su aspecto coriáceo.
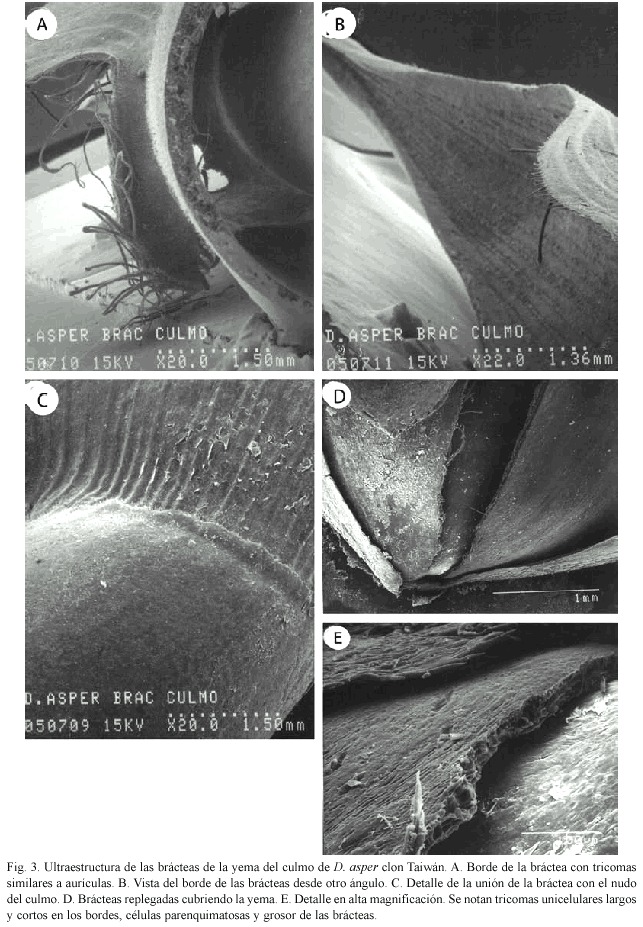
Yema: Las brácteas que cubren las yemas en el culmo, están replegadas una sobre otra y son de textura coriácea (Fig. 3D). En la figura 3E aparece el detalle de estas estructuras, se observa el grosor de cada bráctea y sus células parenquimatosas, también tricomas unicelulares largos y otros cortos en menor número, así como parte de la ornamentación de la bráctea.
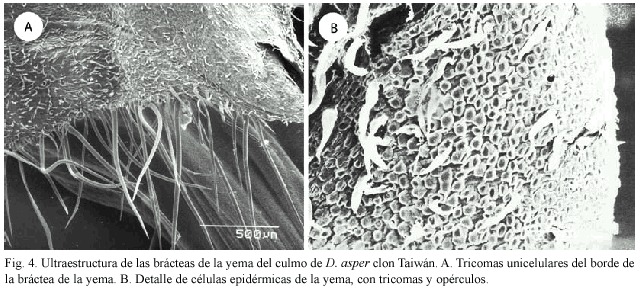
En el borde de la bráctea se observan tricomas unicelulares muy largos y otros pequeños (Fig. 4A). El detalle de las células epidérmicas de la yema tiene gran cantidad de tricomas unicelulares pequeños, así como gran cantidad de opérculos, producto del desprendimiento de los tricomas (Fig. 4B).
Clon Tailandia
Lámina foliar: La epidermis abaxial presenta la vena central y las venas laterales (zonas costales e intercostales) bien definidas. La vena central presenta células largas, pocas papilas céricas, tricomas unicelulares cortos y largos (Fig. 5A). Las nervaduras laterales tienen múltiples hileras de papilas céricas, así como estomas de alto domo (Fig. 5B). La figura 5C tiene las papilas céricas cuticulares de forma alargada y en la figura 5D se pueden apreciar muy bien los estomas, los tricomas unicelulares largos, las células de sílice, células en forma de gancho y el patrón de distribución de la cera cuticular.
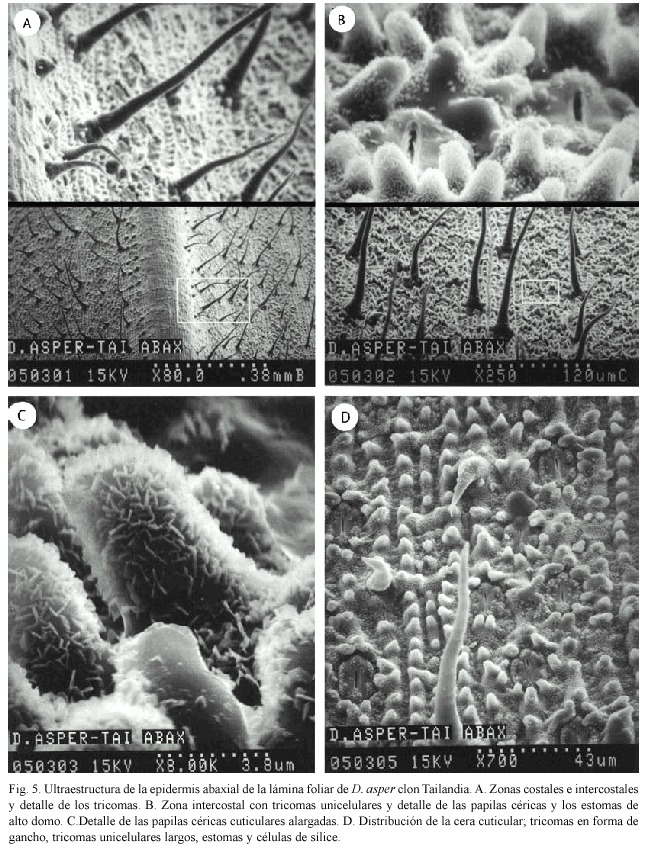
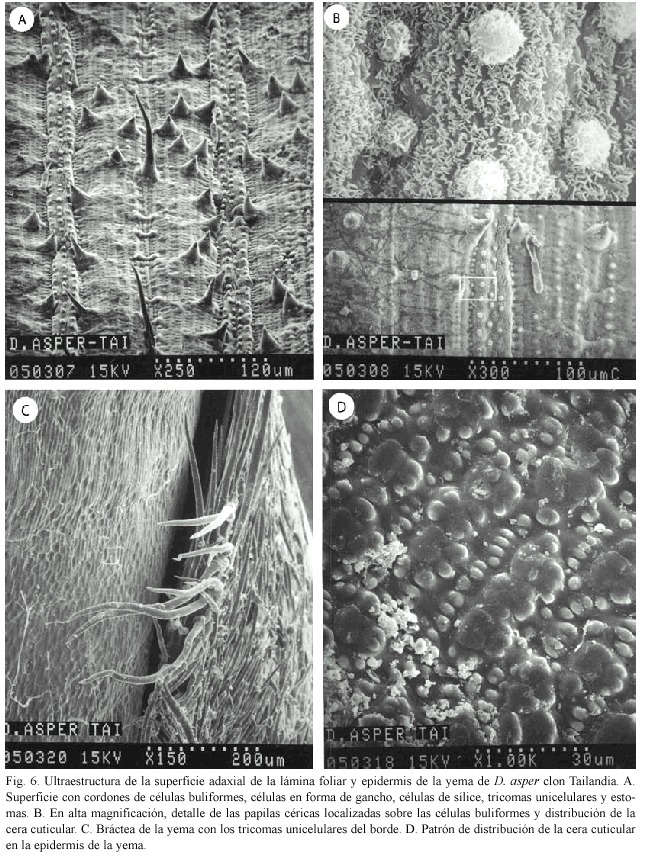
En la zona adaxial, las zonas intercostales presentan tricomas en forma de gancho, tricomas largos unicelulares, células de sílice y dos largos cordones de células buliformes (Fig. 6A). También se observan células en forma de gancho y las hileras de papilas céricas, las cuales a mayor aumento, se presentan esféricas (Fig. 6B).
Bráctea del culmo: Las características estructurales son iguales a las del clon Taiwán, razón por la cual se omite su descripción y no se incluyen figuras.
Yema: Esta estructura se encuentra protegida por brácteas al igual que en el clon Taiwán y también hay tricomas unicelulares en los bordes de ellas (Fig. 6C). En la epidermis se tiene un arreglo bastante lineal de la cera cuticular, en grupos de tres acumulaciones y en medio de ellas, hileras de cera cuticular bien definidas. En otras zonas del mismo tejido, la distribución es desordenada, presentando grandes acumulaciones de cera (Fig. 6D).
Culmo y rama: La distribución celular es igual para ambos clones y no presentan diferencias significativas. La descripción proporcionada se fundamenta en las observaciones hechas en cortes longitudinales y transversales del clon Tailandia. También es importante aclarar que rama y culmo son similares en su corte.
En el corte transversal de la rama se observan los haces vasculares con dos vasos de metaxilema y uno de protoxilema; la sección floemática y el tejido parenquimatoso con contenido celular. Conforme los tejidos avanzan en su retracción hacia la periferia del culmo o rama, se observa más tejido esclerenquimatoso (Fig. 7A).
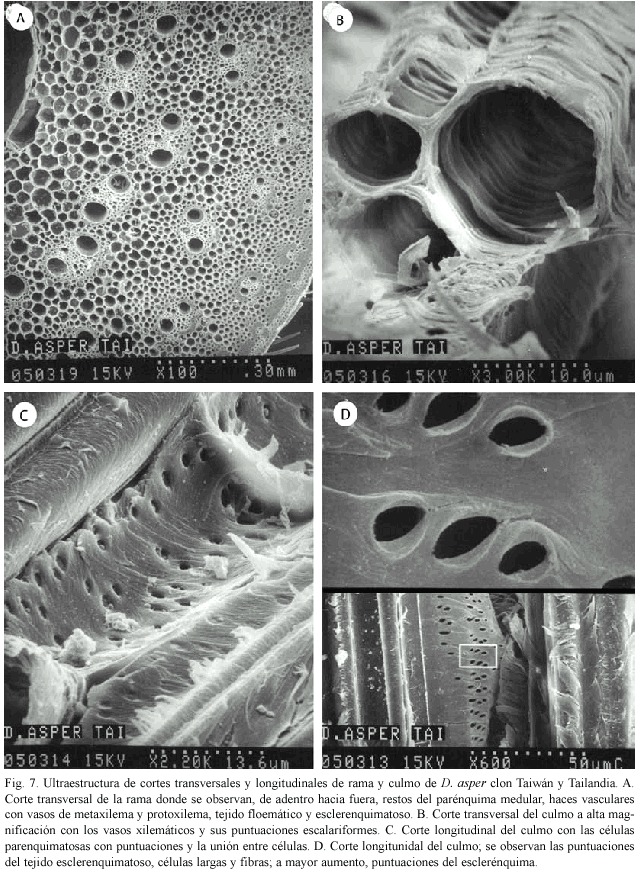
A mayor magnificación es posible observar los vasos xilemáticos con perforaciones escalariformes (Fig. 7B).
En corte longitudinal se observa el tejido parenquimatoso que rodea los haces vasculares, con células que presentan puntuaciones y acumulaciones, que posiblemente corresponden a almidón y a la unión entre células (Fig. 7C). El mismo tipo de corte permite apreciar las puntuaciones del tejido esclerenquimatoso, células largas y fibras. A mayor aumento se observan las puntuaciones del esclerénquima con paredes engrosadas (Fig. 7D).
Discusión
Al comparar estructuralmente ambos clones se encuentran similitudes y diferencias, siendo éstas últimas resultado quizá del aislamiento geográfico de los clones en el continente asiático.
Diferencias: en la lámina foliar el clon Taiwán presenta estomas de bajo domo y en el clon Tailandia son de alto domo. Además, la cera cuticular es escasa y las papilas céricas son esféricas en el clon Taiwán y no están presentes sobre los cordones de células buliformes de la superficie adaxial. Por el contrario, el clon Tailandia presenta abundante cera cuticular y las papilas céricas son alargadas, excepto las que se encuentran sobre los cordones de células buliformes.
El patrón de acumulación de la cera cuticular también difiere, en el clon Taiwán hay dos tipos: en las brácteas en forma de hilos y sobre la lámina de la hoja en forma esférica; en el clon Tailandia hay cuatro tipos: en las brácteas en forma de hilos, sobre las células buliformes en forma esférica, sobre la superficie de la lámina de la hoja en forma alargada y en la superficie de la yema en grupos de tres o en forma desordenada.
Las yemas del clon Taiwán presentan gran cantidad de tricomas alargados pero no acumulación de cera, mientras que en las yemas del clon Tailandia la cera es abundante y presenta los patrones de acumulación descritos anteriormente.
Similitudes: La superficie de las láminas foliares de ambos clones presenta tricomas unicelulares largos y tricomas espinosos; además, los estomas se encuentran rodeados por papilas céricas.
En lo que respecta a la nervadura central, los haces vasculares y las células epidérmicas también son muy parecidos (Grosser y Liese 1971).
Las brácteas protectoras de las yemas tienen gran cantidad de tricomas unicelulares largos, característica común al género Dendrocalamus. Las brácteas del culmo son prácticamente iguales en ambos clones.
Se espera que este estudio sea de utilidad para taxónomos y cultivadores de estos clones, los cuales son de gran importancia económica pero se confunden con gran facilidad.
Agradecimientos
Se agradece a la vicerrectoría de Investigación de la Universidad de Costa Rica por el apoyo que ha dado a este proyecto, así como al Centro de Investigación en Estructuras Microscópicas de la Universidad de Costa Rica por las facilidades brindadas. Este trabajo se realizó con la ayuda financiera de la Comunidad Europea, en el marco del Proyecto Guadua: Research for Sustainable Management and Markets of Bamboo in Colombia and Costa Rica (EU INCO ICA4-CT-2001-10091).
Resumen
El análisis ultraestructural de los clones taiwanés y tailandés de Dendrocalamus asper, realizado al microscopio electrónico de barrido, mostró que el clon de Taiwán tiene estomas de bajo domo en la lámina foliar, mientras que el de Tailandia tiene estomas de alto domo; las acumulaciones de cera cuticular en el clon de Taiwán son de dos formas (cuatro en el de Tailandia). La distribución de los tejidos vasculares es igual en ambos, así como los tejidos que los rodean. Sería difícil identificar estos clones por la observación macroscópica o microscópica de las brácteas del culmo, pues no presentan diferencias significativas: una identificación fiable requiere considerar la ultraestructura de los estomas y las acumulaciones de cera cuticular.
Palabras clave: ultraestructura, microscopia electrónica de barrido, anatomía, bambú, Poaceae, Dendrocalamus asper, clon Taiwán, clon Tailandia.
Referencias
Ghosh, S. S. & B. S. Negi. 1959. Anatomical of Indian bamboo. Part I. Indian For. 86: 719-727. [ Links ]
Grosser, D. & W. Liese. 1971. On the anatomy of Asian bamboos with special reference to their vascular bundles. Wood Sci. Technol. 5: 290-312. [ Links ]
Metcalfe, C. R. 1960. Anatomy of the monocotyledons. I. Gramineae. Oxford Clarendon, Oxford, Inglaterra. 731 p. [ Links ]
Seethalakshmi, K. K. & M. S. Muktesh Kumar. 1998. Bamboos of India: A compendium. Brill, Leiden, Los Países Bajos. 342 p. [ Links ]
Tateoka, T., S. Inoue & S. Kawano. 1959. Notes on some grasses IX: Systematic significance of bicelular microhairs of leaf epidermis. Bot. Gaz. 121: 80-991. [ Links ]
Widjaja, E. A. 1995. Dendrocalamus asper (Schult f.) Backer ex Heyne, p. 80-93. In Dransfield, S. & E. A Widjaja (eds.). Plant Resources of South-East Asia N º 7. Bamboos. Backhuys, Leiden, Los Paises Bajos. 189 p. [ Links ]

