











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.54 no.1 San José mar. 2006
Una nueva especie de pez del género Hemibrycon (Characiformes:Characidae)
César Román-Valencia, Raquel I.Ruiz C.1 & Ramiro Barriga2
1 Universidad del Quindío, Laboratorio de Ictiología, A. A. 460, Armenia, Quindío, Colombia; croman@uniquindio.edu.co & zutana_1@yahoo.com
2 Escuela Politécnica Nacional, Depto. Ciencias Biológicas, Museo de Zoología, Apartado 17-01-2759, Quito, Ecuador; rbarriga@server.epn.edu.ec
Recibido 22-VII-2005. Corregido 01-VIII-2005. Aceptado 25-X-2005.
Abstract: Una nueva especie de pez del género Hemibrycon (Characiformes: Characidae). Hemibrycon pautensis (Characiformes, Characidae), a new fish species from Paute River, eastern Ecuador is described. Diagnostic characteristics: eight to nine branched rays in the dorsal fin (vs.six to seven), and 27 –28 in the anal fin (vs.16 –26, except in H. dariensis which presents 22 –27, in H. metae 26 –31 and H. jabonero 23 –28); a no occurrence of dorsal pharyngeal plate (vs. occurrence); a cartilaginous and divided-in-two basihial (vs. an osseous base and a cartilaginous upper part). Hemibrycon pautensis resembles H. metae by its oblique external edge of the pelvic fins. They can be distinguished by the position of the pectoral fins in relation to the snout (38.24-41.6% in H. pautensis vs. 21. 21-25.87) and by the position of the pectoral fins in relation to the origin of the dorsal fin (20.95-24.30 in Hemibrycon pautensis vs. 35. 89-42.63), and by the number of proximate radials in the pectoral girdle (five in Hemibrycon pautensis vs. three to four). In addition, the geographic distribution of H. metae is restricted to the upper part of the Meta River in Colombia and can be distinguished of H. boquiae by: the number of scales between the lateral-line and the origin of the dorsal fin (eight in H. pautensis vs. 5-7); the distance between the snout and the pelvic fins (38.00-42.90% in H. pautensis vs. 42.9-46.19%); the pelvic fins length (13.77-17.96% in H. pautensis vs. 10.72-13.21%); and the snout length (21.34-27.88 in H.pautensis vs.26.92-33.66%).Rev.Biol.Trop.54(1):209-217.Epub 2006 Mar 31.
Key words: Hemibrycon sp. nov. characid fish, Amazonian, Ecuador, South America.
El género Hemibrycon Günther está conformado por peces de mediano a gran tamaño, entre 100 y 200 mm de longitud estándar. Se caracterizan por presentar más de siete dientes en el maxilar y cuatro dientes en la hilera interna del premaxilar (Eigenmann 1927). Su color es predominantemente plateado con una mancha humeral oscura (Géry 1977). Sin embargo, el significado filogenético de estos caracteres usados en su definición no ha sido demostrado y no existe un análisis filogenético disponible para Hemibrycon. Estos únicamente se utilizan en la definición "tradicional" de Hemibrycon. Al igual que otros carácidos, (por ej. Hyphessobrycon Ellis, Astyanax Baird & Girard, Hemigrammus Gill, Moenkhausia Eigenmann), probablemente no representan una unidad monofilética y su taxonomía está mal estudiada (Lucena 1993,2003).
Actualmente en Hemibrycon se reconocen como válidas 19 especies (Román-Valencia 2001, 2004, Eschmeyer 2005), con distribución en Alto Amazonas en Bolivia, Perú, Ecuador y Colombia; cuenca de los ríos Atrato, León y drenajes Costeros del Pacifico y Caribe, drenajes andinos y alto río Meta en Colombia; cuenca del río Tuyra al oriente de Panamá; al occidente en la cuenca del lago de Maracaibo, pie de monte llanero y cordillera de la costa en Venezuela, isla de Trinidad, Guyanas Británica y Surinam.
Para Ecuador, se han reconocido tres especies: Hemibrycon coxeyi Fowler 1943 para la cuenca del río Pastaxa, H. orcesi Böhlke 1958 para el Alto río Morona y la especie tipo H. polyodon Günther 1864 para Guayaquil (Eschmeyer 2005).
En el presente artículo se describe una nueva especie de Hemibrycon de Ecuador, se utilizan caracteres tradicionales, y además, su osteología, para ilustrar mejor la diferenciación con sus congeneres.
Material y métodos
El material fue preservado en alcohol etílico al 70%, se depositó en el museo de Zoología de la escuela Politécnica Nacional de Ecuador, Quito (MEPN) y en el laboratorio de ictiología, Universidad del Quindío, Armenia, Colombia (IUQ). Además, se examinó material depositado en el museo de Ciencias Naturales de Filadelfia, USA (ANSP), Instituto de Investigaciones Biológicas "Alexander Von Humboldt", Villa de Leyva, Boyacá (IaVH), el Instituto de Ciencias Naturales-Museo de Histria Natural, Universidad Nacional, Bogotá (ICNMNH) y Museo Nacional del Instituto Smithsonian, Washington (USNM).
Las medidas de los ejemplares se tomaron con un calibre digital, hasta centésimas de milímetro. Todas las medidas fueron obtenidas punto a punto. Los recuentos de radios, escamas y dientes se realizaron bajo estereoscopio. Las medidas y recuentos (Cuadro 1) se realizaron sobre el lado izquierdo de los ejemplares, excepto cuando éstos estaban deteriorados en tal lado. Medidas y conteos siguen a Weitzman y Malabarba (1999).
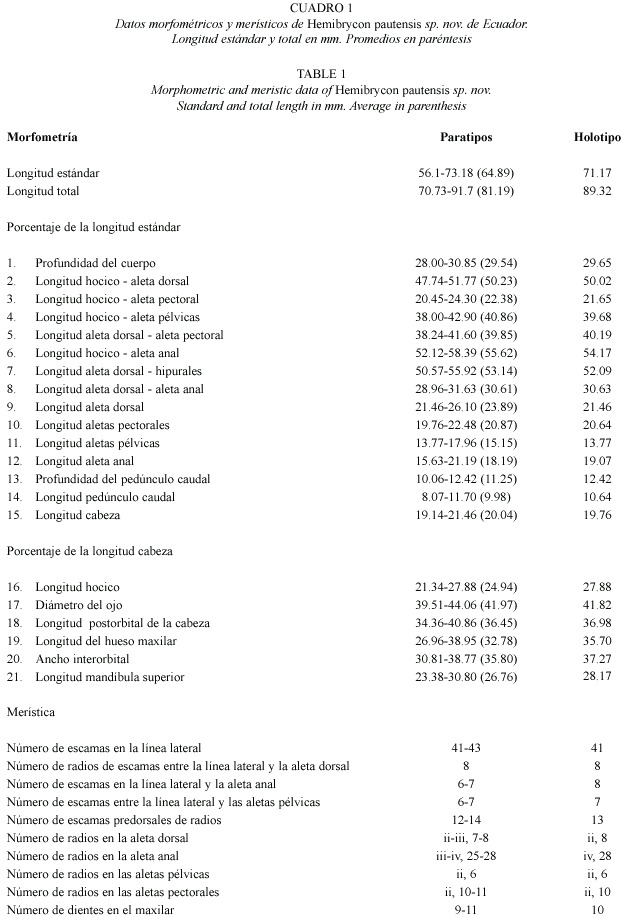
Las observaciones de las estructuras óseas y cartilaginosas se hicieron sobre ejemplares clareados y teñidos (C.&T.) de acuerdo a los métodos descritos por Taylor y Van Dyke (1985), Song y Parenti (1995) con modificaciones. La nomenclatura de los huesos se basó en Weitzman (1962) y Vari (1995). El conteo y las determinaciones de vértebras se hizo sobre material diafanizado y sigue a Helfman et al. (1997).
Material de comparación examinado: Hemibrycon boquiae (véase Román-Valencia 2001). Hemibrycon dariensis: USNM 293245, 28 ejemplares; Panamá, Darien, Río Tuira. 08º N y 77º O, provincia de Darien, río Pucuro acerca 3-4 Km. arriba confluencia con río Tuira, Pacifico, 17 feb 1985. IUQ 523, 26 ejemplares; Colombia, Departamento de Antioquia, río Zungo carretera, sistema río León, dic. 17 de 1990. IUQ 524, dos ejemplares; Colombia, Antioquia, quebrada Km. 25 vía Mutata-Chigorodo, dic. 1990. IUQ 525, 26 ejemplares; Colombia, Antioquia, sistema río León, río Villarteaga, dic. de 1990. ICNMNH 3057, cuatro ejemplares; Colombia, Antioquia, sistema río León, río Mutata, dic. de 1990. ICNMNH 3074, cuatro ejemplares; Colombia, Antioquia, Sistema río León, quebrada Murcia, nov. 13 de 1990. ICNMNH 804, 11 ejemplares; Colombia, Guajira, sistema río Rancheria, Quebrada Arroyo Mamón, Guajira, sept. 9 de 1981. Hemibrycon metae. IaVH 1934, un ejemplar; Colombia, Casanare Río Tua, Mpio de Monterrey. IAvH 3122, 10 ejemplares; Colombia, Casanare, municipio de Aguazul, afluente Río Cachiza, Quebrada Chichaca; marzo de 1994. IaVH 3124, 17 ejemplares; Colombia, Casanare, Cuencas ríos Pauto, Tocaria, Chave y Cravo Sur, noviembre de 1995. IaVh 3125, 33 ejemplares; Colombia,Casanare, cuenca Ríos Unete, Cravo Sur y Tua. IaVH 3127, dos ejemplares; Colombia, Casanare, Quebrada Barreña. IaVH 3129, 50 ejemplares; Colombia, Casanare, Quebrada Cupiagua-cuenca Río Unete. IaVH 3134, cuatro ejemplares; Colombia, Casanare Río Nunchia. IaVH 2986, tres ejemplares; Colombia, Casanare, Ríos Pauto, Tocaria, Charre y Cravosur, marzo 1996. IavH 2973, siete ejemplares; Colombia, Casanare, quebrada Palmicha, afl.Río Cachiza. IaVH 2991, tres ejemplares; Colombia, Casanare Ríos Unete, Cusiana y Tua. Hemibrycon coxeyi: ANSP 70155 (Holotipo), Ecuador, Hacienda Las Mascota, bocas del Río Pastaza, cuenca del Río Marañón. Hemibrycon polyodon: MEPN 001532, 14 ejemplares, Ecuador, provincia de Zamora-Chinchipe, quebrada Tunanza afluente Río Zamora, marzo 15 de 1979, col R. Barriga. MEPN 001533, 11 ejemplares, Ecuador, Provincia de Zamora, quebrada a Antonio-Guadalupe, marzo 21 de 1979, col. R. Barriga. MEPN 001534, 21 ejemplares, Ecuador, provincia de Zamora-Chinchipe, finca Torres Río Zamora, marzo 24 de 1979, col. R. Barriga. MEPN 001535, 15 ejemplares, Ecuador, provincia de Zamora-Chinchipe, quebrada Mayaycu a 2 km en la desembocadura en el Río Nangarifza a 200 m del destacamento militar Mayaycu, 18 de julio de 1993, col. R. Barriga. MEPN 001536, 10 ejemplares, Ecuador, provincia Morona-Santiago, Río Gualaquiza, marzo 8 de 1979, col. R. Barriga, R. Lema, M. Ibarra. MEPN 001537, tres ejemplares, Ecuador, provincia Zamora-Chinchipe, playa del Río Nangarifza al frente del destacamento Militar, julio de 1993, col. R. Barriga. Hemibrycon orcesi: ANSP 75904 (Paratipos con 33 vértebras en total), dos ejemplares, Ecuador,santiago-Zamora, cabeceras del Río Macuma,tributario Río Morona, altura 550-650 m. MEPN 001538, 117 ejemplares, Ecuador, Provincia Morona-Santiago, Río Tayusa afluente del Upano bajo el puente en la vía Méndez-Sucua, 4 de mayo de 1991, col. R. Barriga. MEPN 001539, 66 ejemplares, Ecuador, provincia de Morona-Santiago, Río Tayusa afluente del Upano bajo el puente en la vía Méndez-Sucula, 4 de mayo de 1991, col. R. Barriga. MEPN 001540, 10 ejemplares, Ecuador, provincia Pastaza, agua abajo de Sarayaco, agosto de 1956, col M. Olalla. MEPN 001541, cinco ejemplares, Ecuador, provincia Pastaza, brazo del Río Pastaza, proyecto Aquarrap, 22 de julio de 1999, col B. Chernoff y R. Barriga. MEPN 001542, dos ejemplares, paratipos. Provincia Pastaza, Río Macuma, abril 1953, col. R. Olalla. MEPN 001543, dos ejemplares, provincia de Pastaza, Río Bobonaza-Canelos cuenca Río Pastaza, col. M. Olalla abril 1953. B. orteguasae. ANSP 70504 (holotipo), Colombia, Departamento del Caquetá, Florencia, cuenca del Amazonas, Río Orteguaza; 1932. IUQ 408, 16 ejemplares, Colombia, Departamento del Putumayo, Orito, cuenca del Putumayo, Río Orito en el puente vía a Caldero, junio 28 de 1998. IUQ 425, un ejemplar, Colombia, Departamento del Caquetá, Florencia, cuenca del Orteguaza, Quebrada La Yuca a 1 km de la vía Florencia-Morelia (1º3613"N, 75º3823"O), diciembre 16 de 1998.
Resultados
Hemibrycon pautensis sp. nov. (Cuadro 1, Fig.1)

Holotipo. MEPN 001549, macho 71.7 mm LE; Ecuador, provincia de Morona-Santiago, cuenca del Amazonas, río Paute en la desembocadura del río Namangoza (aprox. 78º25 N, 2º32 S), 7 de mayo de 1991, R. Barriga.
Paratipos. MEPN 001548, 14 ejemplares colectados con el holotipo. IUQ 533, dos ejemplares (C.&T.) colectados con el holotipo. MEPN 001549, cuatro ejemplares; Ecuador, provincia de Morona-Santiago, cuenca del Amazonas, en la desembocadura del río Namangoza, 7 de mayo de 1991, R. Barriga.
Diagnosis. Hemibrycon pautensis se distingue de las demás especies del género por presentar: de ocho a nueve radios ramificados en la aleta dorsal (vs. seis a siete), y de 27 a 28 radios ramificados en la aleta anal (vs. 16-26, excepto en H. dariensis con 22-27, en H. metae con 26 a 31 y en H. jabonero con 23 a 28), placa faringea dorsal ausente (vs. presente), basihial cartilaginoso y dividido en dos (vs. óseo en su base, cartilaginoso en su extremo superior), último diente ventral del maxilar separado por un espacio sin dientes (vs. diente continuo con la fila). Hemibrycon pautensis es parecida a H. metae por el borde externo de las aletas pélvicas oblicuo. Se distinguen por la posición de las aletas pectorales con relación al extremo del hocico (38, 24-41, 6 en H. pautensis vs. 21, 21-25, 87) y por la posición de las aletas pectorales con relación al origen de la aleta dorsal (20.95- 24.30 en Hemibrycon pautensis vs. 35.89- 42.63), y por el número de radiales proximales en la cintura pectoral (cinco en Hemibrycon pautensis vs. tres a cuatro). Además, la distribución geográfica de H. metae está restringida a la cuenca alta del río Meta en Colombia. Se distingue de H. boquiae por el número de escamas entre la línea lateral y el origen de la aleta dorsal (ocho en H. pautensis vs. 5-7), por la distancia entre el hocico y el origen de las aletas pélvicas (38.00-42.90% en H. pautensis vs. 42.9- 46.19%), por la longitud de las aletas pélvicas (13.77- 17.96% en H. pautensis vs. 10.72-13.21%), y por la longitud del hocico (21.34- 27.88 en H. pautensis vs. 26.92- 33.66%)(véase cuadro 1 de Román-Valencia 2001 para datos de H. boquiae).
English diagnosis. Hemibrycon pautensis can be distinguished from its congeners by eight to nine branched rays in the dorsal fin (vs. six to seven), and 27 – 28 in the anal fin (vs.16 – 26, except in H. dariensis which presents 22 – 27, in H. metae 26 – 31 and H. jabonero 23 – 28); a no occurrence of dorsal pharyngeal plate (vs . occurrence); a cartilaginous and divided-in-two basihial (vs. an osseous base and a cartilaginous upper part); maxilla ventral teeth last separated (vs. tooth continuous). Hemibrycon pautensis resembles H. metae by its oblique external edge of the pelvic fins. They can be distinguished by the position of the pectoral fins in relation to the snout (38.24-41.6% in H. pautensis vs. 21.21-25.87) and by the position of the pectoral fins in relation to the origin of the dorsal fin (20.95-24.30 in Hemibrycon pautensis vs. 35.89- 42.63), and by the number of proximate radials in the pectoral girdle (five in Hemibrycon pautensis vs. three to four). In addition, the geographic distribution of H. metae is restricted to the upper part of the Meta River in Colombia and can be distinguished of H. boquiae by: the number of scales between the lateral-line and the origin of the dorsal fin (eight in H. pautensis vs. 5-7); the distance between the snout and the pelvic fins (38.00-42.90% in H. pautensis vs. 42.9-46.19%); the pelvic fins length (13.77- 17.96% in H. pautensis vs. 10.72-13.21%); and the snout length (21.34-27.88 in H. pautensis vs. 26.92- 33.66%).
Descripción. Cuerpo alargado; área dorsal de la cabeza convexo. Perfil dorsal de la cabeza y del cuerpo elevado desde el supraoccipital hasta el origen de la aleta dorsal, e inclinado hacia abajo desde el último radio de la aleta dorsal hasta la base de la aleta caudal. Perfil ventral del cuerpo convexo desde el hocico hasta la base de la aleta anal, cabeza y hocico corto; mandíbulas iguales en longitud; boca terminal; labios blandos y flexibles, no cubren la hilera externa de dientes del premaxilar; el premaxilar presenta dos procesos uno ascendente que se articula al mesetmoide y otro proceso lateral que sostiene los dientes y en que se articula lateralmente el proceso ascendente del maxilar; borde ventral de la premaxilar plano, se articula al proceso lateral del maxilar; extremo posterior del maxilar sobrepasa el borde anterior de la órbita.
Cinco a seis infraorbitales presentes, con canal latero sensorial; tercer infraorbital más largo, ancho, borde ventral y posterior en contacto con el preopérculo.Supraorbital presente. Premaxilar con proceso lateral corto y puntiagudo. Presenta dos hileras de dientes; hilera externa con tres a cuatro dientes tricúspides, sus bases dispuestas en línea recta. Hilera interna con cuatro dientes pentacúspides, con la cúspide central mucho mayor. Maxilar con siete a once dientes que cubren la mayor parte de su borde, dientes dorsales pentacúspides y ventrales tricúspides, último diente ventral del maxilar separado por un espacio sin dientes. Dentario con cuatro grandes dientes frontales pentacúspides, el cuarto se observa doblado o no recto, la cúspide central tiende hacia el extremo posterior, dientes con la cúspide central mucho mayor, seguidos por siete a nueve dientes pequeños con uno a tres cúspides.
La parte ventral del supraoccipital se une al arco exoccipital por una banda de cartílago, también en el extremo superior del complejo neural y entre el margen ventral del complejo neural y la superficie dorsal de la tercera vértebra. Rinoesfenoides óseo, pero en contacto con el orbitoesfenoides a través de cartílago. Borde anterior de la aleta dorsal recto, segundo radio simple y primeros dos radios ramificados más largos. Basihial cartilaginoso y dividido en dos. Placa faringea dorsal ausente. Pteriogióforos proximales dorsales insertados entre las espinas neurales 13 a 21. Seis a ocho supraneurales presentes con cartílago en los extremos superior e inferior.
Cintura pectoral articulada a la parte postero lateral del cráneo por la fusión del supracleitrum y el extremo ventral del postemporal, unidos al borde dorsal del cleitrum. El cleitrum se ubica bajo el borde ventral del opérculo. Presencia de cartílago en la unión de la escápula con la superficie interna del supracleitrum. Cinco radiales proximales. Aleta pélvica corta, su extremo no alcanza el origen de la aleta anal. El hueso pélvico es una estructura alargada,se localiza paralelo al área central del cuerpo; hueso pélvico corto, y recto, con cartílago en sus extremos anterior y postero lateral; el proceso isquial es una estructura corta, recta y con una apófisis puntiaguda que presenta cartílago filamentoso.
Aleta caudal bifurcada con lóbulos cortos y puntiagudos. Radios caudales principales 8-10/9-10. Aleta caudal sin escamas. Cuarenta y uno a 43 escamas con poros en la línea lateral, terminan sobre la aleta caudal. En cuatro ejemplares se registraron 14 escamas con poros en la línea lateral. Los poros de la línea lateral forman una leve curva entre la primera y la onceava escamas, el resto están dispuestas en línea recta. Número total de vértebras 40-41.
Dimorfismo sexual secundario. Los machos tienen una hilera de seis a 23 espinas cortas sobre los radios ramificados de la aleta pélvica sobre las dos ramas del radio ubicadas sobre cada segmento, una sola espina sobre cada porción. Los primeros ocho radios ramificados y el tercer radio simple de la aleta anal con seis a 18 con espinas puntiagudas.
Color en vivo. Descrita con base en foto de un ejemplar vivo en un acuario mantenido alrededor de dos meses, al cabo del cual murió y se procedió a su fijación en formalina al 10%. Cuerpo azul oscuro sobre fondo plateado en la parte dorso-lateral, área latero ventral a partir de la cabeza hasta el orificio anal blanco amarillento. Se observa mancha anaranjada en la parte dorsal del ojo. Con mancha humeral oscura verticalmente alargada y mancha peduncular oscura, la cual no se prolonga sobre los radios medios caudales. Aletas pectoral y pélvicas rosadas, aletas dorsal, anal y caudal azul oscura. Mancha rosada se ubica en el extremo posterior del pedúnculo caudal, en la base de los radios caudales inferiores.
Distribución. Conocida sólo de su localidad típica, el río Paute
Etimología. El epíteto específico alude a la localidad típica, el río Paute
Discusión
Varios aspectos observados en la nueva especie es necesario resaltar: la presencia de supraorbital con la formación del canal supraorbital, que se une medialmente dando origen a la barra epifiseal sobre la fontanela craneana y que se continua con el canal laterosensorial. De igual manera, se registra éste hueso y sus interrelaciones con otras estructuras mencionadas en Bryconamericus hypopterus y B. orteguasae (Román-Valencia 2003), en Hemibrycon boquiae (Román-Valencia et al. sometido1), aunque Weitzman (1962) lo describe en Brycon meeki generalizado para los carácidos y afirma:"... El borde antero-superior de la órbita está rodeado por un hueso alargado, no asociado con un canal laterosensorial". Además, Román-Valencia et al. (2003) cita la presencia de supraorbitales en Grundulus bogotensis y en G. cochae, Ruiz y Román-Valencia (sometido) describen el hueso supraorbital para Astyanax aurocaudatus (nombre válido). Sin embargo, Weitzman y Malabarba (1998), Malabarba y Weitzman (2003) plantearon que los tetragonopterinae (tal y como es definido por Géry (1977) carecen de supraorbitales. En éstas condiciones, se puede considerar los supraorbitales un carácter variable en carácidos y no sustentable para distinguir tetragonopterinae de Brycon. Contrario a lo planteado por Weitzman y Malabarba (1998). La observación de éste hueso en algunos géneros de Characidae (como se anota en éste trabajo) sugiere su presencia en los taxones mas primitivos.
El cuarto diente no recto presente en H. pautensis, se puede considerar una divergencia en otros grupos relacionados a Hemibrycon; por ejemplo, es un carácter presente y variable en especies de Astyanax (Ruiz y Román-Valencia sometido), como también en especies de Hemibrycon (H. jelski, H. metae, H. orcesi, y H. polyodon).
La parte ventral del supraoccipital se une al arco exoccipital por una banda de cartílago, no es tan usual en Characidae. La hemos observado en Bryconamericus orteguasae (Román-Valencia et al. sometido 2) y en Astyanax aurocaudatus (Ruiz y Román-Valencia sometido); y en otras especies de Hemibrycon: H. polyodon, H. orcesi, H. jelskii.
De las especies registradas para Ecuador, el nuevo taxón se diferencia: de H. orcesi por la posición de las aletas pectorales con relación al extremo del hocico (38.24- 41.60 en H. pautensis vs. 24.95-29.80), por la posición de la aleta anal con relación al extremo del hocico (52.12- 58.39 en H. pautensis vs. 59.18- 66.93), por la longitud de la cabeza (19.14- 21.46 en H. pautensis vs. 23.17-30.37), por el número de escamas en la línea lateral (41-43 en H. pautensis vs. 34-36), por el número de escamas desde la línea lateral hasta el origen de la aleta dorsal (ocho en H. pautensis vs. cinco a seis), por el número de escamas desde la línea lateral hasta el origen de la aleta anal (seis a siete en H. pautensis vs. cuatro a cinco), por el número de radios ramificados en la aleta anal (25-28 en H. pautensis vs. 17-20) y por el número total de vértebras (39-40 en H. pautensis vs. 35). De H. jelski el nuevo taxón se distingue por la longitud de la cabeza (19,40- 21,46 en H. pautensis vs. 21,10- 25,66), por el número de escamas en la línea lateral (38- 41 en H. pautensis vs. 41-43), por el número de escamas predorsales (12-14 en H. pautensis vs. 10-12) y por el número total de vértebras (39-40 en H. pautensis vs.37-38).
De H. polyodon el nuevo taxón se distingue por el número de escamas entre la línea lateral y el origen de las aletas pélvicas (seis a siete en H. pautensis vs. cinco a seis) y por el número de supraneurales (seis a ocho en H. pautensis vs. siete). De H. coxeyi, al comparar su descripción original (Fowler 1943) con el material examinado del nuevo taxón, se observaron diferencias en la altura del cuerpo (3.2- 3.6 en H. pautensis vs. 3.2), en la longitud de la cabeza (4.7-5.2 en H. pautensis vs. 4.0), en el hocico con relación a la cabeza (3.6- 4.7 en H. pautensis vs. 6.0), en el diámetro del ojo con relación a la cabeza (2.2- 2.53 en H. pautensis vs. 3.0), en el diámetro del ojo con relación al ancho interorbital (ojo mayor que el interorbital en H. pautensis vs. ojo igual al interorbital) y en el diámetro del ojo con relación a la cabeza (2.5- 3.2 en H. pautensis vs. 3.5), en el número de radios de las aletas pélvicas (ii, 6 en H. pautensis vs. i.7), en el número de escamas de la línea lateral (41-43 en H. pautensis vs. 39), en el número de escamas transversas entre el origen de la aleta dorsal y la línea lateral (ocho en H. pautensis vs. siete), en el número de escamas el origen de la aleta anal (seis a siete en H. pautensis vs. cinco) y en el número de escamas predorsales (12-14 en H. pautensis vs. 17).
Agradecimientos
La Universidad del Quindío (Comité Central de Investigaciones) financió un viaje de estudio al Ecuador (C.R.-V.y R.I.R.). Idea Wild suministró reactivos utilizados en la transparentación de material. Maria Mercedes Azpelicueta (MLP, Argentina), Francisco Langeani (UNESP, Brasil)y un revisor anónimo leyeron el manuscrito y plantearon valiosas sugerencias y correcciones.
Resumen
Se describe Hemibrycon pautensis (Characiformes: Characidae), una especie nueva de pez del río Paute al oriente de Ecuador. El nuevo taxón se distingue de las demás especies por las siguientes caracteristicas: ocho a nueve radios ramificados enaleta dorsal (vs. seis a siete), y 27 –28 en la aleta anal (vs.16 –26, excepto en H. dariensis la cual presenta 22 –27, en H. metae 26 –31 y H. jabonero 23 –28); no presenta placa faringea dorsal (vs. presenta); basihial dividido en dos y cartilaginoso (vs. óseo y cartilaginoso en la parte superior). Hemibrycon pautensis es parecida a H. metae es parecida a H. metae por el borde externo de las aletas pélvicas oblicuo. Se distinguen por la posición de las aletas pectorales con relación al extremo del hocico (38.24- 41.6 en H. pautensis vs. 21.21-25.87) y por la posición de las aletas pectorales con relación al origen de la aleta dorsal (20.95-24.30 en Hemibrycon pautensis vs. 35.89-42.63), y por el número de radiales proximales en la cintura pectoral (cinco en Hemibrycon pautensis vs. tres a cuatro). Además, la distribución geográfica de H. metae está restringida a la cuenca alta del río Meta en Colombia. Se distingue de H. boquiae por el número de escamas entre la línea lateral y el origen de la aleta dorsal (ocho en H. pautensis vs. 5-7), por la distancia entre el hocico y el origen de las aletas pélvicas (38.00-42.90% en H. pautensis vs.42.9-46.19%), por la longitud de las aletas pélvicas (13.77-17.96% en H. pautensis vs. 10.72-13.21%), y por la longitud del hocico (21.34-27.88 en H. pautensis vs. 26.92-33.66%).
Palabras clave: Hemibrycon sp. nov., Pez caracido, Amazonia, Ecuador, Suramérica.
Referencias
Eigenmann, C. H. 1927. The American Characidae. Mem. Mus. Comp. Zool. XLIII (4): 311-428+24 láminas. [ Links ]
Eschmeyer, W. 2005. CAS. Ichthyology-Catalog of fishes, California Academy of Sciences, San Francisco, CA, USA. 2905 p. (Tambien disponible en linea, http://www.calacademy.org/research/ichthyology/catalog//fishcatsearch.html. [ Links ]
Fowler, W. H. 1943. A collection of fresh-water fishes from Colombia, obtained chiefly by Brother Niceforo Maria. Proc. Acad. Nat. Sci. Phil. XCV: 223-266. [ Links ]
Géry, J. 1977. Characoids of the world. T. F. H., Neptune City, Nueva Jersey, EEUU. 672 p. [ Links ]
Lucena, C. A. S. 1993. Estudo filogenético da família Characidae com uma discussâo dos grupos naturais propostos (Teleostei, Ostariophysi, Characiformes). Tesis de doctorado, Universidade de São Paulo, São Paulo, Brasil. 158 p. [ Links ]
Lucena, C. A. S. 2003. New characid fish, Hyphessobrycon scutulatus, from the rio Teles Pires drainage, upper rio Tapajós system (Ostariophysi: Characiformes: Characidae). Neotrop. Ichthyol. 1: 93-96. [ Links ]
Malabarba, L. R. & S. H. Weitzman. 2003. Description of a new genus with six new species from Southern Brazil, Uruguay and Argentina, with a discussion of a putative characid clade (Teleostei: Characiformes: Characidae). Comun. Mus. Ciênc. Tecnol. PUCRS, Sér Zool., Porto Alegre, 16: 67-151. [ Links ]
Román-Valencia, C. 2001. Redescripción de Hemibrycon boquiae (Pisces:Characidae), especie endémica de la quebrada Boquía, cuenca Río Quindío, alto Cauca, Colombia. Dahlia. Rev. Asoc. Colomb. Ictiol .4: 27-32. [ Links ]
Román-Valencia, C. 2003. Sistemática de las especies Colombianas de Bryconamericus (Characifomes, Characidae). Dahlia (Rev. Asoc. Colomb. Ictiol.) 6: 17-58. [ Links ]
Román-Valencia, C. 2004. Redescripción de Bryconamericus tolimae (Pisces:Characidae), especie endémica del Río Combeima, cuenca del Río Magdalena, Colombia. Dahlia. Rev. Asoc. Colomb. Ictiol. 7: 23-27 [ Links ]
Román-Valencia, C., Hans-Joachim P. & F. Pantoja. 2003. Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae) para Colombia y redescripción de Grundulus bogotensis (Humboldt y Valenciennes, 1833). Mem. Fund. La Salle de Cienc. Nat., Caracas 155: 51-72. [ Links ]
Román-Valencia, C., R. I. Ruiz & L. M. López. Sometido1. Osteología comparada de Hemibrycon boquiae y Bryconamericus caucanus (Characiformes: Characidae). Rev. Mus. Arg. Cienc. Nat., n. s. [ Links ]
Román-Valencia, C., R. I. Ruiz C. & M. B. López. Sometido2. Osteología comparada de Bryconamericus hypopterus y B. orteguasae (Characiformes: Characidae). Rev. Mus. Arg. Cienc. Nat., n. s. [ Links ]
Ruiz, R. I. & C. Román-Valencia. Sometido. Sobre la validez de Astyanax aurocaudatus Eigenmann, 1913 (Pisces: Characidae) en Alto Cauca, Colombia. Animal Biodiversity and Conservation. [ Links ]
Song, J. & L. R. Parenti. 1995. Clearing and staining whole fish specimens for simultaneous demostration of bone, cartilage and nerves. Copeia 1995: 114-118. [ Links ]
Taylor, W. R. & G. C. Van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium 9: 107-119. [ Links ]
Vari, R. P. 1995. The neotropical fish family Ctenoluciidae (Teleostei: Ostariophysi: Characiformes) supra and intrafamilial phylogenetic relationships, with a revisionary study. Smith. Contr. Zool. 564: 1-96. [ Links ]
Weitzman, S. H. 1962. The osteology of Brycon meeki ,a generalized characid fish, with an osteological definition of the family. Stanford Icthyol. Bull. 8: 1-77. [ Links ]
Weitzman, S. H. & L. R. Malabarba. 1998. Perspective about the phylogeny and classification of the characidae (Teleostei: Characiformes), p. 161-170. In L. R Malabarba, R. E. Reis, R. P Vari, Z. M. S. Lucena & C. A. S. Lucena (eds.). Phylogeny and classification of neotropical fishes. Porto Alegre, Edipucrs, Brasil. [ Links ]
Weitzman, S. H. & L. R. Malabarba. 1999. Systematics of Spintherobolus (Teleostei: Characidae: Cheirodontinae) from eastern Brazil. Icthyol. Explor. Freshwaters 10:1-43. [ Links ]

