











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.53 supl.1 San José Mai. 2005
Patterns of sexual recruitment of acroporid coral populations on the West Fore Reef at Discovery Bay,Jamaica
Norman J.Quinn1 & Barbara L.Kojis2
1 Discovery Bay Marine Laboratory,P.O.Box 35,University of the West Indies,Discovery Bay,St.Ann,Jamaica.Fax: (876)973-3091;norman.quinn@uwimona.edu.jm
2 Division of Fish and Wildlife,6291 Estate Nazareth,Red Hook,St.Thomas,U.S.Virgin Islands 00802; bkojis@vitelcom.net
Received 15-I-2004. Corrected 03-VIII-2004. Accepted 29-III-2005.
Abstract: Coral recruitment was examined on terracotta tiles deployed for four six-month periods between March 2001 and April 2003 on the West Fore Reef at Discovery Bay,Jamaica.During each sampling period, four tiles were deployed on each of two arrays at six depths ranging from 3 m to 33 m.Only three Acropora spat recruited to the tiles over the sampling period.The Acropora spat recruited during only one of the four six-month sampling periods and at only one depth,3m.That represents a density of 8 spat m-2 at 3 m depth for one six-month sampling period.Acropora recruitment represented <1 %of the total spat recruiting to the tiles deployed at 3 m during the four sampling periods.Density of acroporids on the West Fore Reef is low.Only one Acropora colony (an A.palmata )was recorded during Point-Quarter surveys of coral cover and density at depths of 3 m,9 m,14 m and 19 m.Considering the paucity of acroporid colonies and the infrequent settlement of acroporid spat on the West Fore Reef,it is unlikely that the historic abundance of A.palmata and A. cervicornis will return soon.
Key words :Setting plates,planulae,Caribbean,Scleractinian,sexual recruitment,Acropora.
Several decades ago a complex sequence of interacting events resulted in a phase shift in the shallow water acroporid coral community on the north coast of Jamaica (Hughes et al.1987,Lessios 1988,Hughes 1994).This community changed from a coral to an algal dominated system.
Many hypotheses have been proposed to explain the lack of recovery of the coral assemblages (e.g. Knowlton et al.1990,Hughes 1994).One of these is the lack of sexual recruitment of the major reef building corals in the family Acroporidae (Kojis and Quinn 1993,2001).This study sought to determine the density,depth distribution and seasonal variability of sexual recruitment of acroporid corals along the West Fore Reef at Discovery Bay.It was hypothesized that owing to a paucity of mature acroporid colonies there would be low levels of larval recruitment.
Recruitment patterns of juvenile corals are important to the overall community structure of coral reefs (Bak and Engle 1979,Harrison and Wallace 1990).Studies frequently focus on juvenile corals >1 cm diameter called "visible " recruitment.Since the post-settlement process may change the number of recruits (Keough and Downes 1982),recruitment onto tiles is considered to be a better indication of the availability of planktonic planulae than the study of "visible " corals (Harrison and Wallace 1990).As well, early settlement stages are more vulnerable than adults to changes in nutrient (Ward and Harrison 1996)and sediment levels (Gilmore 1999).
Materials and methods
Site Description
Benthic arrays were located at the CARICOMP coral reef site (18º 28.397 N, 77º 27.831 W)situated at the West Fore Reef of Discovery Bay,Jamaica in a mixed /buttress zone at six depths:3 m,9 m,14 m,19 m,26 m and 33 m.
Habitat characterization
In the late 1960 s this zone had high diversity and abundance of coral (Goreau and Wells 1967)and was dominated by large colonies of Acropora palmata surrounded by A.cervicornis , Montastrea annularis ,Porites spp.and large, towering colonies of Dendrogyra cylindrus (Woodley and Robinson 1977).After Hurricane Allen in 1980 and Hurricane Gilbert in 1988 the coral abundance and diversity decreased (Woodley et al.1981,Woodley 1989)and populations of A.palmata and A.cervicornis dropped almost to 0%coral cover (Knowlton et al.1981). In 2002-2003,the results from surveys estimated the percentage coral cover at a depth of 3 m -9 m at ~15%(Linton and Fisher 2004).
Sampling technique
Coral recruitment:Coral recruitment arrays were constructed by attaching four 208 cm2 unglazed terracotta tiles to a PVC array (Fig.1).The tiles were smooth on one side and had 12 ridges on the other side.Two of the tiles were arranged horizontally and two vertically on the array.The tiles were ~0.8 m above the substrate.

The arrays were initially installed in late March /early April 2001 and were replaced in October 2001 ("summer 2001 " sampling period),April 2002 ("winter 2001/2002 "),October 2002 ("summer 2002 ")and in April 2003 ("winter 2002/2003 ").The scleractinian and milleporan corals were counted (standardized to number recruits m-2)and identified to family where possible,using a binocular microscope.Because the tiles were deployed for only six months,fleshy algal cover was low and,therefore considered only a minor competitor for space.
Coral Community Survey: Hard coral communities around the benthic settlement arrays were quantitatively sampled in May 2003 using the plotless Point-Centered Quarter or Point-Quarter method (POCQ)(Cox 1972). At the depths where acroporids were historically common and arrays were in place (3 m, 9 m,14 m,and 19 m),scuba divers swam along an imaginary line and dropped a hammer at approximately 2 –5 m intervals.A sample point was established at the intersection of the hammer handle and head.Four quadrants were then formed by establishing one axis along the hammer handle and another at right angles to it along the hammerhead.The coral nearest the sample point in each quadrant was located and the species,size (maximum length and width) and distance from the center of the corallum to the sample point were recorded.Twenty sample points were surveyed at each depth.The overall density of colonies m-2 ,percentage coral cover and relative percentage cover for each species was calculated according to the formulae in Randall et al.(1988).
Results
Coral recruitment
Over a two-year period,three acroporid spat settled during only one of the four sampling periods (summer 2001)(Fig.2), and only on the tiles at 3 m depth (Fig.3).No Acropora recruitment occurred on tiles at 9 m, 14 m,19 m,26 m or 33 m.Acroporid spat density at 3 m during the summer 2001 sampling period was 8 spat m-2 .This was the sampling period and depth with the highest spat density. Acroporid settlement only represented 1%of the coral settlement during this sampling period and depth.A subsequent paper will analyze the settlement of other coral recruits.
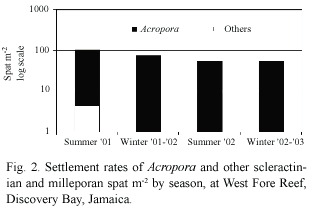
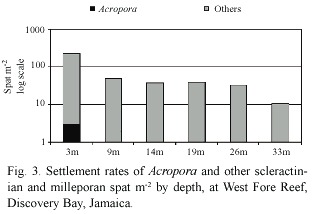
Coral Community Survey
Twenty-five hard coral species were recorded in the POCQ surveys (Table 1).The percentage live coral cover ranged from 8.9% at 9 m to 16.7%at 19 m.Adult Acropora palmata was only recorded at 3 m representing 0.3%of the live coral cover at this depth.No acroporid sexual recruits and no adult A.cervicornis were recorded in the POCQ surveys or observed at this site.However,approximately 10 km to the west at Braco Reef A.palmata recruits in the density of about 0.3 m-2 were observed on reef sections recently cleared by Diadema (Kojis and Quinn,unpublished).The density of coral colonies at the West Fore Reef was 0.85 m-2 at 3 m,1.03 m-2 at 9 m,1.41 m-2 at 14 m and 3.70 m-2 at 19 m.The density of A. palmata was only 0.01 m-2 at 3 m,and 0 m-2 at all other depths.
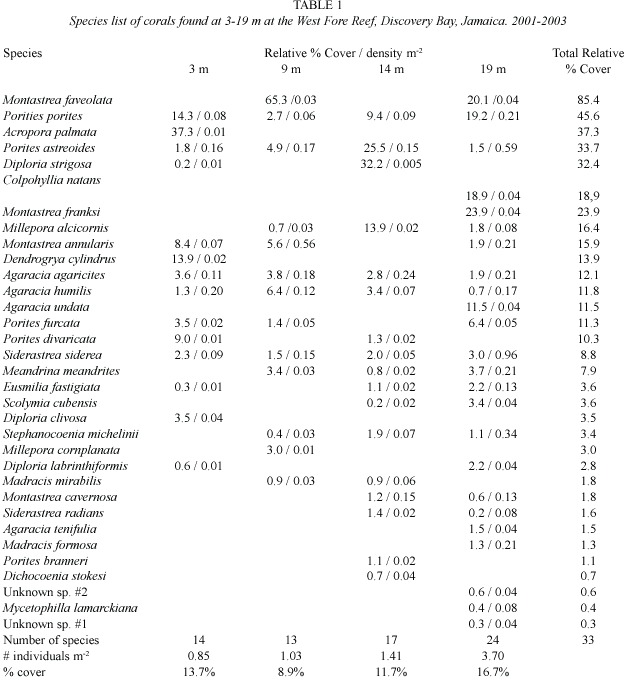
Discussion
It has been recognized that many factors such as hurricanes,reduced water quality associated with terrestrial runoff and poor watershed management,diseases,thermal stress, overgrowth by macro algae, boat groundings and anchor damage are involved in the continued deterioration of abundance,health and overall percent cover of Acropora cervicornis and A.palmata (Precht et al.2002).
Dense,nonspecific high relief thickets of Acropora palmata (elkhorn coral)and A. cervicornis (staghorn coral)were once characteristic of shallow and intermediate depth coral communities in the Caribbean prior to the late 1970 s (Goreau 1959,Tunnicliffe 1983).In the early 1980 s,A.cervicornis live coral cover was >21%at several sites around Discovery Bay but was nearly extirpated by 1987 (Knowlton et al. 1990).The dense thickets of A.palmata and A. cervicornis have not reestablished in nearly 25 years and,based on the low recruitment rates of acroporids recorded in this study,are unlikely to reestablish on the West Fore Reef of Discovery Bay for some time.By comparison,acroporid settlement on undisturbed reefs in Fiji are typically several hundred spat m-2 during the summer period (Kojis and Quinn 2001).
Acropora palmata and A.cervicornis exhibit patchy abundance and distribution at other sites near Discovery Bay and at other locations in the West Atlantic (Vargas-Angel et al.2003).On the northern Jamaican coast near Discovery Bay,colonies of A.cervicornis are common at 9 m at Dairy Bull, approximately 3 km from Discovery Bay,where they have been recorded to contribute more than 10% of the live coral cover in a small area (Quinn and Kojis,unpublished).Several small (<1 m diameter) A.palmata colonies are present at 10 m at west Rio Bueno reef and the density at this location is higher than at the West Fore Reef. Some of the colonies are >2.5 m at their longest horizontal dimension.
The lack of recovery of acroporids can either be linked to localized factors or a general paucity of planktonic recruits.It is important to identify the mechanism at work.If high algal density is thwarting community recovery by occupying planulae settlement space, then a reduction in algal cover would free up settlement space and coral recruitment should increase.If there are few coral planulae being produced by acroporids,then available space will have little bearing on the potential for acroporid recovery.In that case,transplanting of acroporid fragments may provide the best chance of improving the rate of recovery in the immediate future (Kojis and Quinn 1981).
It has been suggested that predators were preventing the recovery of staghorn populations as predation rates increased to levels beyond a predicted threshold as the result of a phase shift (Knowlton et al.1990).While the level of juvenile mortality from predation may be a key factor in the restructuring of some coral communities (Smith 1992,Smith 1997, Miller et al.2000),we suggest that the recruitment failure of acroporid planulae is more influential than juvenile mortality.
Because Caribbean acroporid reproductive strategy emphasized asexual fragmentation over sexual recruitment (Precht et al.2002)and the existing populations are so small it is considered unlikely that they will rapidly recover through growth and asexual fragmentation.The evidence from this study supports Kojis and Quinn s (2001)hypotheses that sexual recruitment of acroporids is currently low because there are few adult colonies to produce gametes and because of the potential lack of fertilization success when the distance between colonies is so great (Allee effect)(Allee 1931).With the additional impediment to settlement of a macro algal overgrown substrate,the possibility of successful sexual recruitment is further reduced.Currently,sexual reproduction is not likely to contribute to rapid recovery of acroporid communities, although the presence of some planulae recruits does indicate that sexual recruitment is available to reestablish populations on reefs with no acroporids.
The lack of sexual recruits is likely a function of both the present paucity of large populations of Acropora and the poor health of the existing populations.Several decades ago acroporid species were the dominant genus at depths <12 m (Goreau 1959, Goreau and Wells 1967).Currently at the West Fore Reef at Discovery Bay,they represent <1%of the coral individuals present depths <10 m. Furthermore, existing populations are under attack by various coral diseases (Aronson and Precht 2001), which are likely to reduce the fecundity of individual colonies.Considering the massive decline in acroporid populations Caribbean wide,it is likely that the larval pool has considerably decreased.
Kojis and Quinn (1993)noted that recovery of acroporid populations in the Caribbean may be hindered by high growth rates and frequent fragmentation which may reduce the amount of energy available for sexual reproduction and result in low gamete production rates and,therefore,low larval recruitment rates.Even when there were larger acroporid populations their sexual recruits rarely, if ever,settled on plates (Rogers et al.1984, Tunnicliffe 1983)or experimental substrata of dead Acropora cervicornis (Rylaarsdam 1983) or were rarely found in permanent quadrants (Porter and Meier 1992).Metabolic resources may be used to enhance growth at the expense of sexual reproduction.
Agariciid and poritid spat are the most abundant taxa on settlement plates in the Caribbean e.g .Barbados (Tomascik 1991,Hunte and Wittenberg 1992);Bermuda (Smith 1992); Curacao (van Moorsel 1985);East Flower Garden (Bagget and Bright 1985);Florida Keys (Smith 1997)and St.Croix (Rogers et al.1984).The brooding life history strategy of these genera and their relative abundance are considered important factors in establishing this dominance (Chiappone and Sullivan 1996). Planulae from brooding corals are ready to settle upon release.In contrast,acroporids are broadcast spawners and the gametes must first successfully fertilize and develop before they are ready to settle.This can occur as early as 2-3 days post fertilization (Heyward et al.1987),but is more likely to occur after several days.Broadcast spawning corals represent 4%of the recruits to plates in the US Virgin Islands (Kojis and Quinn 2001)and 2% in Jamaica (Sammarco 1980).Consequently, larval recruitment is likely rare for the main framework builders such as Acropora spp.and Montastrea spp.on Caribbean reefs (Kojis and Quinn 2001).While acroporid populations are nearly extirpated in some locations, healthy individuals still do exist and increasing numbers of colonies with sexual recruits have been observed at a few locations (Zubillaga et al.2003).Surprisingly,at the high latitude reef off Fort Lauderdale,Florida,an extensive, healthy population of Acropora cervicornis is thriving (Vargas-Angel et al.2003).As recommended in a recent Acropora workshop, experimental acroporid mariculture and other propagation techniques,along with transplantation,and reattachment of dislodged Acropora fragments may provide a feasible strategy to rebuild degraded population and require further study (Bruckner 2002).
Acknowledgments
We gratefully acknowledge the use of the facilities at the Discovery Bay Marine Laboratory (DBML),University of the West Indies and are also grateful for support through the Tropical Discoveries Fund.K.Purser kindly provided some diving assistance.This is DBML publication #667.
Resumen
El reclutamiento de corales fue estudiado utilizando placas de cerámica colocadas en cuatro períodos de seis meses,entre marzo de 2001 y abril de 2003,en el Arrecife Frontal Oeste en Discovery Bay,Jamaica.Durante cada período de muestreo se colocaron dos grupos de cuatro placas a seis profundidades,en un ámbito de 3-33 m.Se encontraron tres reclutas de Acropora durante todo el estudio, únicamente a 3 m de profundidad y durante uno de los cuatro períodos.Esto representa una densidad de 8 reclutas m-2 a 3 m,para un semestre de muestreo.Acropora representó <1%del total de reclutas de las placas colocadas a 3 m durante los cuatro períodos de muestreo.Se encontró una densidad baja de acropóridos en el Arrecife Frontal Oeste. Únicamente se encontró una colonia de Acropora palmata durante los censos de cobertura coralina y densidad realizados a profundidades de 3,9,14 y 19 m.Considerando la escasez de colonias de acropóridos y el asentamiento infrecuente de reclutas en el Arrecife Frontal Oeste,la recuperación de la abundancia histórica de A.palmata y A. cervicornis,es poco probable a corto plazo.
Palabras clave:Placas de asentamiento,planula,Caribe, Scleractineos,reclutamiento sexual,Acropora.
References
Alee,W.C.1931.Animal Aggregations.A study in General Sociology.Univ.Chicago Press,Chicago.437 p. [ Links ]
Aronson,R.B.&W.F.Precht.2001.White-band disease and the changing face of Caribbean coral reefs. Hydrobiol.460:25-38. [ Links ]
Bagget L.S.&T.J.Bright.1985.Coral recruitment at the East Flower Garden Reef (northwestern Gulf of Mexico).Proc.5 th Int.Coral Reef Symp.,Tahiti 4: 379-384. [ Links ]
Bak,R.P.M.&M.S.Engle.1979.Distribution,abundance and survival of juvenile hermatypic corals (Scleractinia)and the importance of life histories strategies in the parent coral community.Mar.Biol. 54:341-352. [ Links ]
Bruckner,A.W.2002.Proceedings of the Caribbean Acropora workshop:Potential application of the U.S. Endangered Species Act as a conservation strategy. NOAA Tech.Memor.NMFS-OPR-24,Silver Spring, MD.199 p. [ Links ]
Chiappone,M.&K.M.Sullivan.1996.Distribution,abundance and species composition of juvenile scleractinian corals in the Florida reef tract.Bull.Mar.Sci.58: 555-569. [ Links ]
Cox,G.W.1972.Laboratory Manual of General Ecology. W.C.Brown,Dubuque.195 p. [ Links ]
Gilmore.J.1999.Experimental investigation into the effects of suspended sediment on fertilization,larval survival and settlement in a scleractinian coral.Mar. Biol.135:451-462. [ Links ]
Goreau,T.F.1959.The ecology of Jamaican coral reefs I.Species composition and zonation.Ecology 40: 67-90. [ Links ]
Goreau,T.F.&J.W.Wells.1967.The shallow –water Scleractinia of Jamaica:revised list of species and their vertical distribution range.Bull.Mar.Sci.17: 442-453. [ Links ]
Harrison,P.&C.C.Wallace.1990.Reproduction,dispersal and recruitment of scleractinian corals,p.133-207.In Z.Dubinsky (ed.).Ecosystems of the World.Vol.25. Elsevier,New York. [ Links ]
Heyward,A.,K.Yamazato,T.Yeemin &M.Minei.1987. Sexual reproduction of coral in Okinawa.Galaxea 6:331-343. [ Links ]
Hughes,T.P.1994.Catastrophes,phase shifts,and large-scale degradation of a Caribbean reef.Science 265: 1547-1551. [ Links ]
Hughes,T.P.,D.C.Reed &M.J.Boyle.1987.Herbivory on coral reefs:community structure following mass mortality of sea urchins.J.Exp.Mar.Biol.Ecol. 113:39-59. [ Links ]
Hunte,W.&M.Wittenberg.1992.Effects of eutrophication and sedimentation on juvenile corals.II. Settlement. Mar. Biol. 114:625-631. [ Links ]
Keogh,M.J.&B.J.Downes.1982.Recruitment of marine invertebrates:the role of active larval choices and early mortality.Oecologia 54:348-352. [ Links ]
Knowlton,N.,J.C.Lang,M.C.Rooney &P.Clifford.1981. Evidence for delayed mortality in hurricane damaged Jamaican staghorn corals.Nature 294:251-252. [ Links ]
Knowlton,N.,J.C.Lang &B.D.Keller.1990.Case study of natural population collapse:Post hurricane predation of Jamaican staghorn corals.Smithson.Contrib. Mar.Sci.31:1-25. [ Links ]
Kojis,B.L.&N.J.Quinn.1981.Factors to consider in transporting coral to accelerate regeneration of damaged coral reefs.Proc.Environ.Eng.Conf.,1981. Townsville,Queensland.Inst.Eng.Publ.8(81): 183-187. [ Links ]
Kojis,B.L.&N.J.Quinn.1993.Biological limits to Caribbean reef recovery.A comparison with western South Pacific reefs,p.35-41.In R.N.Ginsburg (Comp.).Global Aspects of Coral Reefs,Health, Hazards,and History.RSMAS,Univ.Miami, Florida. [ Links ]
Kojis,B.L.&N.J.Quinn.2001.The importance of regional differences in hard coral recruitment rates for determining the need for coral restoration.Bull. Mar.Sci.69:967-974. [ Links ]
Lessios,H.A.1988.Mass mortality of Diadema antillarum in the Caribbean:what have we learned?Ann.Rev. Ecol. Syst. 19:371-393. [ Links ]
Linton,D.&T.Fisher (eds.).2004.CARICOMP.Caribbean Coastal Marine Productivity Program:1993-2003. CARICOMP, FIU/UWI.91 p. [ Links ]
Miller,M.W.,E.Weil &A.M.Szmant.2000.Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA.Coral Reefs 19:115-123. [ Links ]
Porter,J.W.&O.W.Meier.1992.Quantification of loss and change in Floridian reef coral populations.Amer. Zool.32:625-640. [ Links ]
Precht,W.F.,A.W.Bruckner,R.B.Aronson &R.J.Bruckner. 2002.Endangered acroporid corals of the Caribbean. Coral Reefs 21:41-42. [ Links ]
Randall,R.H.,S.D.Rogers,E.E.Irish,S.C.Wilkins,B.D. Smith &S.S.Amesbury.1988.A marine survey of the Obyan-Naftan reef area,Saipan,Mariana Islands. Univ.Guam Mar.Lab.Tech.Rep.No.90:56 p. [ Links ]
Rogers,C.S.,H.C.Fitz III,M.Gilnak,J.Beets &J.Hardin. 1984.Scleractinian coral recruitment patterns at Salt River submarine canyon,St.Croix.U.S.Virgin Islands.Coral Reefs 3:69-76. [ Links ]
Rylaarsdam,K.W.1983.Life histories and abundance patterns of colonial corals on Jamaican reefs.Mar.Ecol. Prog. Ser. 13:249-260. [ Links ]
Sammarco,P.W.1980.Diadema and its relationship to coral spat mortality:Grazing competition and biological disturbance. J.Exp.Mar.Biol.Ecol.45:45-272. [ Links ]
Smith,S.R.1992.Patterns of coral recruitment and post-settlement mortality on Bermuda s reefs;comparisons to Caribbean and Pacific reefs.Amer.Zool.32: 663-673. [ Links ]
Smith,S.R.1997.Patterns of coral recruitment,recruitment and juvenile mortality with depth at Conch Reef,Florida.Proc.8th Int.Coral Reef Symp., Panama 2:1197-1202. [ Links ]
Tomascik,T.1991.Settlement patterns of Caribbean scleractinian corals on artificial substrata along an eutrophication gradient,Barbados,West Indies.Mar. Ecol.Prog.Ser.77:61-69. [ Links ]
Tunnicliff,V.1983.Caribbean staghorn coral populations: Pre-Hurricane Allen conditions in Discovery Bay, Jamaica. Bull. Mar Sci.33:132-151. [ Links ]
Anonymous.1984.Comparing Coral Reef Survey Methods. Rep.Reg.UNESCO /UNEP Workshop.Phuket Marine Biological Centre,Thailand,13-17 December 1982.UNESCO Rep.Mar.Sci.21:1-170. [ Links ]
van Moorsel,G.W.N.M.1985.Disturbance and growth of juvenile corals (Agaricia humilis and Agaricia agaricites, Scleractinia) in natural habitats on the reef in Curacao.Mar.Ecol.Prog.Ser.24:99-112. [ Links ]
Vargas-Angel,B.,J.D.Thomas &S.M.Hoke.2003. High-latitude Acropora cervicornis thickets off Fort Lauderdale, Florida, USA. Coral Reefs 22:465-473. [ Links ]
Ward,S.&P.L.Harrison.1996.The effects of elevated nutrients on settlement of coral larvae during the ENCORE experiment,Great Barrier Reef,Australia. Proc.8th Int.Coral Reef Symp.,Panama.2:891- 896. [ Links ]
Woodley,J.D.1989.The effects of Hurricane Gilbert on coral reefs at Discovery Bay,p.79-82.In P. Bacon (ed.).Assessment of the Economic Impacts of Hurricane Gilbert on Coastal and Marine Resources in Jamaica.CEP Tech. Rep. No. 4 ,UNEP.Caribbean Environment Programme, Kingston,Jamaica. [ Links ]
Woodley,J.D.&E.Robinson.1977.Modern reef localities –West Discovery Bay.Field Guidebook to the Modern and Ancient Reefs of Jamaica.3rd Int.Coral Reef Symp.Atlantic Reef Committee,Univ.Miami, Miami,FL.Pp.18-26. [ Links ]
Woodley,J.D.,E.A.Chornesky,P.A.Clifford,J.B.C. Jackson,L.S.Kaufman,N.Knowlton,J.C.Land,M.P. Pearson, J.W. Porter,M.C.Rooney,K.W.Rylaarsdam, V.J.Tunnicliffe,C.M.Wahle,J.L.Wulff,A.S.C.Curtis, M.D. Dallmeyer,B.P.Jupp,M.A.R.Koehle,J.Niegel &E.M.Sides.1981.Hurricane Allen s impact on Jamaican coral reefs.Science 214:749-755. [ Links ]
Zubillaga,A.L.,C.Bastidas &A.Cróquer.2003. Estandarización de un protocolo para la estimación del estatus poblacional de Acropora palmata.Scient. Meeting Asso.Mar.Lab.Caribbean,Port of Spain, Trinidad.Abstract.Pp.125. [ Links ]

