










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 n.1-2 San José Jun. 2005
Caracterización espacial y biogeográfica de las asociaciones de macroalgas de Bahía del Rincón,Baja California Sur,México
R. Riosmena-Rodríguez, G. Hinojosa-Arango, J. M. López-Vivas, K. León-Cisneros & E. Holguin-Acosta
Programa de Botánica Marina, Departamento de Biología Marina, Universidad Autónoma de Baja California Sur, Apdo. Postal 19-B, La Paz B. C. S. 23080 México. Fax:+5211280880; riosmena@calafia.uabcs.mx
Recibido 30-VIII-2002. Corregido 22-VIII-2003. Aceptado 30-IX-2003.
Abstract
Spatial and biogeographic characterization of macroalgal assemblages from Bahía del Rincón, Baja California Sur, Mexico.Macroalgal studies in Baja California Sur have dealt mainly with occurrence and seasonality, but some areas are poorly known even for these basic data. Bahía del Rincón-La Rivera is an important high-productivity fisheries area where coastal infrastructure development is under way. A spatial characterization of the marine flora from Bahía del Rincón-La Rivera was done by intensive sampling at different depths and localities with skin and SCUBA diving. At least 500m2 were surveyed in each site. Additionally, quantitative sampling was done in ten random 25 cm2 quadrates per site. In the intertidal section, density and cover estimates were used. We also investigated the historical records and geographical affinities. A total of 72 species were identified (most were red algae: 62%). We found no general trend in the biogeographical affinities, which varied with each taxonomic group. Most brown algae species were tropical-endemic; red algae temperate- cosmopolite and green algae tropical-cosmopolite. In the spatial assemblage structure we found a high similarity between the intertidal areas, but a low similarity in shallow or deeper areas (3-5 m). This pattern was the same when we compared the abundance of the main species. We suggest that there are significant spatial differences in recruitment and development of the assemblages in relation to vertical distribution (depth)and position along the shore. There is a clear-cut Gelidium-Jania belt in the intertidal zone and a Padina-Dictyota belt below the low tide. Sporadic and year-round species occur in the intertidal zone, annual and perennial species below the low tide line. Sites differ in recruitment and this affects the abundance of other species (such as coraline and Caulerpa species). Temperature and sedimentation affect seasonality, but community structure is relatively constant throughout the year. Rev. Biol. Trop. 53(1-2):97-109. Epub 2005 Jun 24.
Key words: Gulf of California, seaweeds, ecology, Bahía Rincón, coastal development.
El conocimiento de la flora marina de Baja California Sur se ha desarrollado principalmente en Bahía de La Paz (Riosmena- Rodríguez y Paul-Chávez 1997), Bahía Magdalena (Sánchez-Rodríguez et al.1989), Bahía Concepción (Mateo-Cid.et al.1993), Bahía Tortugas (Mendoza-González y Mateo- Cid 1986), Bahía Asunción (Mateo-Cid y Mendoza-González 1994), el arrecife coralino de Cabo Pulmo (Anaya-Reyna y Riosmena- Rodríguez 1996) y Todos Santos (Mendoza- González y Mateo-Cid 1994). Los estudios han analizado básicamente la presencia y variaciones estacionales de la flora. Asimismo, se han incorporado extensiones de ámbito y nuevos registros para varias especies (Sánchez- Castrejón et al. 1995, Riosmena-Rodríguez et al.1998), lo que constata el carácter dinámico de la flora.
Además existe el desconocimiento de la composición específica y la forma de organización de comunidades de macroalgas dentro de otras áreas de Baja California Sur. Una de las que todavía no han sido bien caracterizadas es la zona entre Bahía de Muertos y Cabo Pulmo. Baynes (1999) ha buscado caracterizar las asociaciones de la zona de Cabo Pulmo pero el trabajo carece de una base taxonómica confiable (no determina a nivel de especie), por lo que sus resultados tienen que ser tomados con precaución. Por otro lado, la zona de Bahía del Rincón-La Rivera se ha caracterizado por su riqueza pesquera asociada a una alta productividad, pero ahora se pretende desarrollar inmobiliariamente la zona. Este estudio se desarrolló en el marco del estudio de impacto ambiental para la zona requerido por SEMARNAP y la legislación mexicana. Es por ello que los objetivos de la presente caracterización son: (1) describir la composición específica de las especies de macroalgas que se distribuyen en la región y sus afinidades biogeográficas y (2) determinar la complejidad espacial de las asociaciones de macroalgas, en particular su distribución vertical.
Material y métodos
Trabajo de campo: Se hizo una prospección a lo largo de la zona de Bahía del Rincón-La Rivera que comprende aproximadamente 10 km de costa entre las localidades de El Surgidero y Las Tachuelas. La mayoría del sustrato disponible fue arenoso y se observaron afloramientos rocosos submareales entre 1-3 m. por lo que se hicieron evaluaciones cada 0.5 km para localizar comunidades de algas. Se determinó trabajar en tres sitios rocosos con tres niveles de análisis: Las Tachuelas (23º 29 32" N,109º 27 25" W; intermareal, 1 m profundidad y 3-4 m), El Hotel (23º 30 N, 109º 30 W; intermareal y 1 m de profundidad) y El Surgidero (23º 37 10" N, 109º 36 04" W; a 3-4 m.). Se visitaron estas localidades ya que no existen otras áreas de sustrato rocoso en la zona somera debido a que la pendiente de la plataforma costera es pronunciada y existen dos arroyos que depositan una gran cantidad de sedimento en el área.
En cada localidad/ profundidad se hizo una revisión cualitativa y otra cuantitativa. Dependiendo de las características de cada localidad se subdividieron por ambientes: a) intermareal, b) submareal somero (1 m) y c) submareal profundo (3-4 m). La recolecta intermareal y submareal somera se realizó durante la marea alta y se utilizó buceo libre; mientras que para los sitios submareales más profundos se utilizó buceo autónomo tipo SCUBA. Para el muestreo cualitativo todos los sitios y ambientes visitados fueron revisados exhaustivamente (un área >500 m2), procurando recolectar el mayor número posible de ejemplares por especie. El número de ejemplares recolectados dependió de su abundancia y variabilidad encontrada, en los sitios con menos individuos la recolecta no fue mayor a 15 organismos. Los especimenes fueron removidos del sustrato con ayuda de una espátula convencional y depositados en bolsas negras de plástico.
Para los muestreos cuantitativos se utilizaron dos metodologías complementarias, en ambos casos la idea fue determinar el porcentaje de cobertura de las especies principales en cada hábitat. Para la zona somera (1 m) y la de profundidad (3-4 m) se utilizaron cuadrantes de 25 cm2 a lo largo de la franja de macroalgas donde se realizaron 10 cuadrantes por sitio de manera aleatoria por números. Para el caso de la zona intermareal se determinó que cualquier metodología in situ no tendría una interpretación adecuada y dado el reducido tamaño del cinturón algal (30 cm de altura) se optó por una metodología alterna. Por ello se utilizó como estimador de la densidad y cobertura de los organismos a las conchas de lapas (Patella mexicana) que son dominantes, además de que presentaron similitudes a la zona y no tienen movilidad extrema. Esto se realizó seleccionando diez individuos al azar por medio de números aleatorios por zona y estimando la cobertura por centímetro linear en cada caso, se estimo que diez organismos sería un buen estimador debido a una curva de rarefacción de Sanders para riqueza (Brower et al.1998) y densidad. En todos los casos se estimó el porcentaje de cobertura de cada una de las especies de la comunidad.
Trabajo de laboratorio: Posteriormente, las algas fueron trasladadas al laboratorio y se seleccionaron por división y morfología para su determinación taxonómica. Para la determinación y actualización nomenclatorial de los especimenes se utilizaron las referencias convencionales para la zona citadas por Riosmena-Rodríguez y Paul-Chávez (1997). Todos los ejemplares fueron procesados y catalogados como parte del Herbario Ficológico de la UABCS (# de catálogo del 2130 a 2207). En el caso de las recolectas intermareales se procesaron las evaluaciones de cobertura con la ayuda de un estereoscopio para determinar la cobertura por especie con un transecto a lo largo de la concha que se subdividió en diez puntos a lo largo del eje principal. Las especies fueron registradas por el número de contactos y registrándose inclusive si se tenía una cobertura múltiple.
Procedimientos de gabinete: Una vez identificados todos los organismos fueron depositados en la colección del Herbario Ficológico de la UABCS, después se procedió a hacer una cuantificación de las especies por división. Además se realizó una revisión exhaustiva de la literatura utilizando las referencias presentadas por Riosmena-Rodríguez y Paul-Chávez (1997) para conocer que especies presentan su ámbito de distribución que abarque esta zona. Además se consideró el número de especies (riqueza) para cada localidad por división.
Con la información de la presencia-ausencia de las especies por localidad se generaron los análisis y las gráficas de similitud, fenológico y el de afinidad biogeográfica. Para el análisis de disimilitud se utilizo el índice de Jaccard, posteriormente se realizó un agrupamiento de los resultados con el método UPGMA para el establecimiento de ligas y para establecer las distancias se utilizó el método de bloques (Manhattan)con el programa STATISTICAR . La graficación se da en distancia de similitud, donde 0 es completamente igual y 1 es totalmente disimilar. Se considera significativo una similitud entre 0 y 0.20.
Para determinar el estado fenológico de todas las especies reportadas se utilizó la nomenclatura de Mathieson (1989) que establece tres categorías por el grado de permanencia en el sistema de las especies donde las especies esporádicas (E) solo habitan momentáneamente el sitio, las anuales o pseduoperenes (A) presentan una generación cada año y las perenes (P) forman una fronda por año con el sujetador permaneciendo por más de uno. En el caso de la Biogeografía se determinó para cada especie reportada o colectada en la región su afinidad biogeográfica en cuatro categorías: Tropical (TR) especies solo distribuidas en los trópicos; Templada (TM) para especies solo distribuidas en las zonas templadas; Cosmopolita (CM) donde son especies con amplia distribución y Endémicas (ED) donde se consideraron especies sólo distribuidas en el Golfo de California.
En el caso de los muestreos cuantitativos se determinaron a especie cada uno de los elementos de los diez cuadrantes colectados. Para cada especie se determinó la media y desviación típica de la cobertura para cada sitio/ profundidad. Los datos de cobertura media se agruparon en cada zona (intermareal, submareal baja (o somera), submareal profunda), en cada caso se determinó que el análisis cuantitativo se requería hacer por género para ajustar los datos a la normalidad. Se realizaron pruebas de normalidad (Chi-cuadrada)y homocedasticidad (prueba de Barlett) a todos los grupos de datos (Zar 1996). Una vez establecidos estos parámetros, se realizó un ANDEVA de una vía (sitio como factor) para las especies de la zona intermareal y un ANDEVA de dos vías (sitio y profundidad como factor) para las especies de la zona somera (1m de profundidad) y profunda (3-4 m). Una vez realizado este análisis se hizo a posteriori una prueba Tukey, todos estos se realizaron con el programa STATISTICAR. Se tabularon todas las abundancias por especie y se graficó a las que sumadas representaran el 85-90% de la varianza.
Resultados
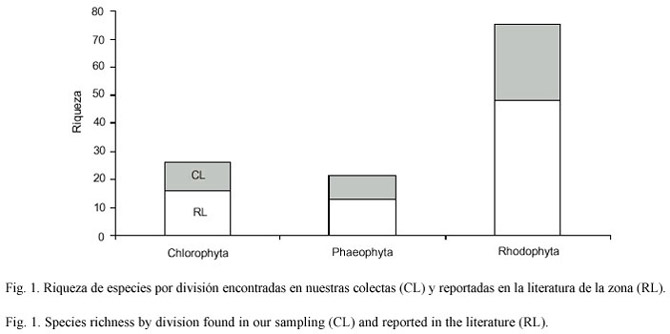
A partir de las recolectas realizadas y la revisión de la literatura se determinaron un total de 72 especies como flora potencial, de las cuales corresponden 45 a recolectas modernas. De todos los registros el 62% de las especies corresponden a la división Rhodophyta, mientras que las algas de la división Chlorophyta y Phaeophyta presentaron porcentajes muy similares con 16 y 13% respectivamente (Fig.1). Esta proporción se mantiene al analizar las colectas actuales y las referencias históricas (Cuadro 1).

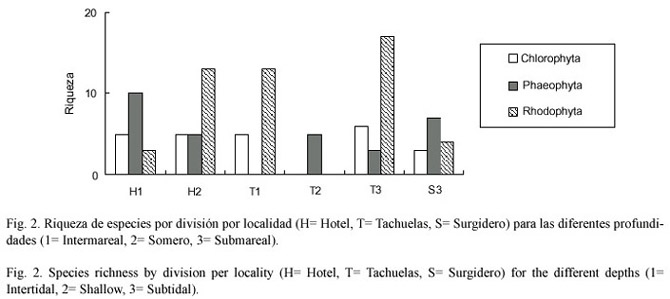
En el caso del elenco por sitio se determinó que la riqueza por división entre las localidades varió muy marcadamente (Fig.2). En el caso de las algas rojas se observó que las Tachuelas- submareal (3-4 m) se observó la mayor riqueza (18 especies), los sitios denominados como Tachuelas-intermareal y Hotel-somero presentaron riquezas similares (12 especies). Las menores riquezas se observaron en la zona submareal del Surgidero (3-4 m)y la del Hotel-intermareal. En el caso de las algas cafés sólo el Hotel-intermareal se mostró diferente (10 especies) a las demás localidades donde se presentan entre 5 y 7 especies. Esto se ve más claramente con las algas verdes, que se presentan con variaciones mínimas entre las localidades de 3 a 5 especies (Fig.2).
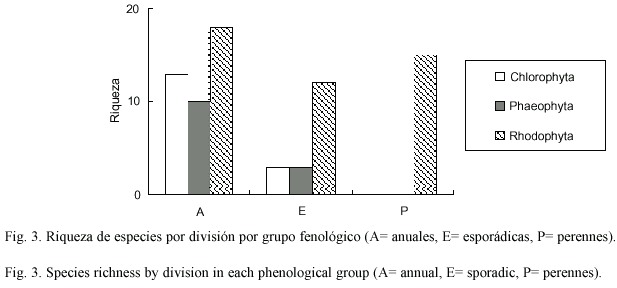
En el caso de las características fenológicas de las especies encontradas se observa que son preponderantemente especies anuales para las tres divisiones (Fig.3). En particular las algas verdes y cafés solo presentan elementos anuales y esporádicos (Fig.3); donde el número es ligeramente mayor en las verdes anuales pero igual en las esporádicas. El caso de las algas rojas es distinto ya que se presentan riquezas muy similares entre los tres elementos, siendo el principal las perennes (Fig.3).
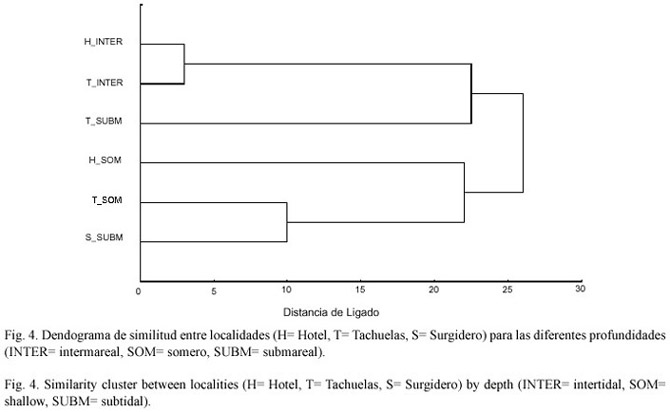
El análisis de similitud muestra un agrupamiento en la composición de especies altamente significativo entre las dos localidades intermareales visitadas (Fig.4). En el caso de las localidades someras y submareales el análisis de similitud encontró que la localidad de Tachuelas-somero, Hotel somero y Surgidero submareal el análisis las agrupa de manera significativa y en este agrupamiento se observa que Tachuelas-somero y Surgidero-submareal tienen mayor relación que Hotel-somero (Fig.4). La única localidad que parece no tener algún vínculo fuerte con la otra es Tachuelas-submareal que se agrupa con mayor similitud con las zonas intermareal que con la somera de cualquiera de las localidades visitadas (Fig.4).
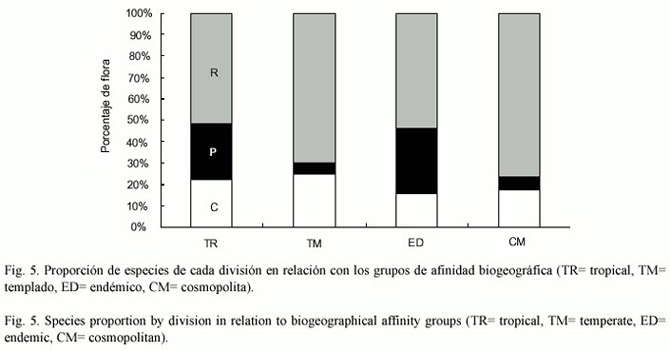
Las afinidades de las especies de la flora potencial presentan un comportamiento diferencial muy marcado entre las divisiones y la región de afinidad (Fig.5). Esto se observa claramente en las algas cafés donde la mayoría de los elementos son de naturaleza tropical/ endémica mientras que los cosmopolitas/ templados son una proporción mínima (Fig.5). Algo inverso se observa en algas rojas donde las templadas/ cosmopolitas representan una mayor proporción que las tropicales/ endémicas (Fig.5).En el caso de las algas verdes se observa que las especies se reparten en la misma proporción (~20%) entre los cuatro tipos de afinidades (Fig.5).
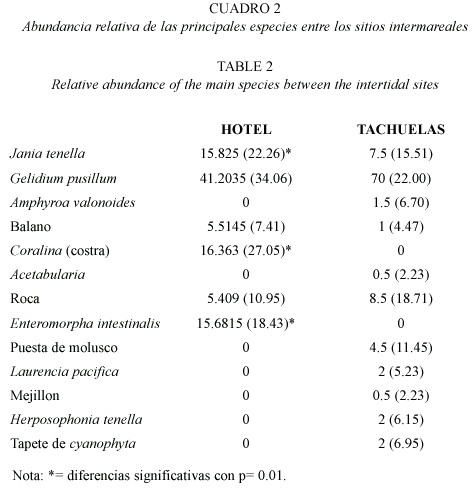
En el caso de los muestreos cuantitativos en la zona intermareal en general, se observó un patrón muy similar entre las algas de la zona del Hotel y Tachuelas (Cuadro 2). La cobertura es dominada por Jania tenella con una diferencia significativa (p<0.5,n=110) entre la zona de Tachuelas y el Hotel casi del 30% (Cuadro 2). Otro de los elementos dominantes fue Gelidium pusillum donde la mayor cobertura observa sobre la roca testigo sin presentar diferencias significativas entre las localidades consideradas. Además, para la zona del Hotel la cobertura de las coralinas costrosas fue significativa (p<.05,n=110) junto con la de Enteromorpha intestinalis y para las Tachuelas el espacio vacío tuvo una presencia detectable (Cuadro 2). Entre los sitios con mayor diversidad de especies representadas fue Tachuelas con 11 especies (Cuadro 2) con alguna cobertura particular y la zona del Hotel existieron seis especies que presentaron cobertura pero sin que esta fuera significativa.
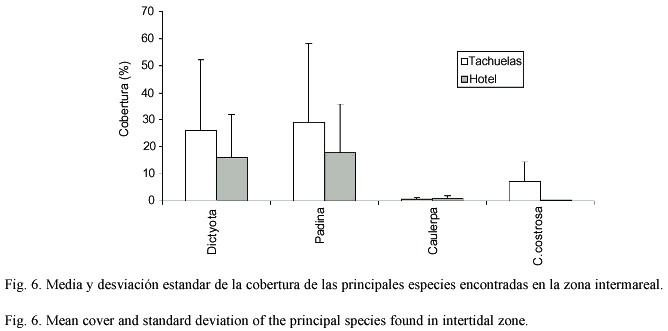
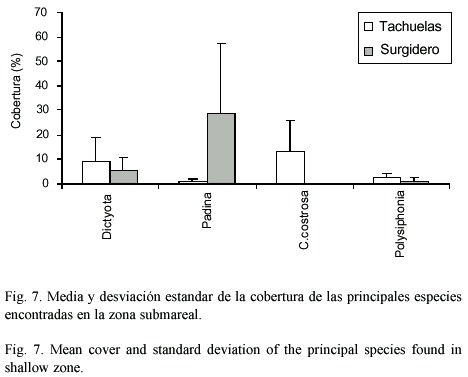
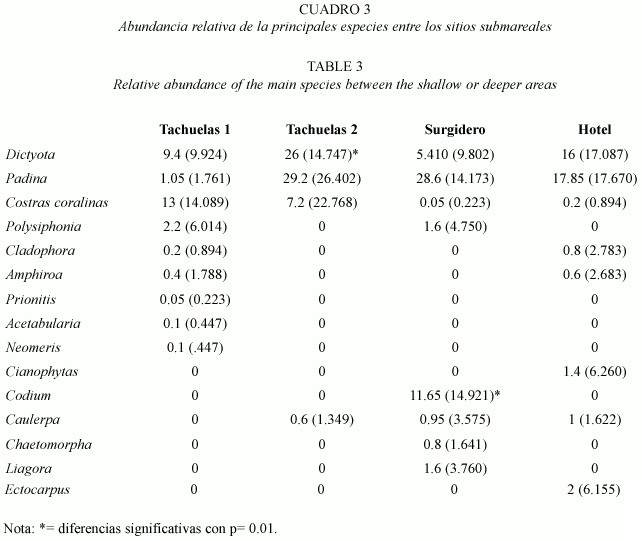
Cuantitativamente las dos zonas someras estudiadas (Tachuelas y Hotel) presentaron componentes principales similares (Dictyota, Padina, Caulerpa y Coralinas costrosas) donde la cobertura fue significativamente superior (p<.05,n=110), en la mayoría de los casos excepto en Caulerpa, para Tachuelas sin llegar a ser significativo estadísticamente a nivel de género pero no es así en el recubrimiento por especie donde si se puede decir que existieron diferencias no estadísticamente verificadas (Fig.6). Entre estos sitios no hay diferencias significativas que se observan en la abundancia de las especies determinadas (Cuadro 3). Mientras que en el caso de la zona profunda (3-4 m) se encontró un patrón similar con coralinas costrosas Dictyota y Padina como las especies principales entre Tachuelas y Surgidero (Fig.7). La única diferencia la constituye la presencia de Polysiphonia. Al aplicar el análisis de varianza se determinó que existen diferencias significativas entre Tachuelas y Surgidero pero no entre este último y el Hotel. Asimismo, la abundancia de Padina y Dictyota se ven afectadas por el sitio y las coralinas costrosas se ven afectadas por la abundancia de Padina y Dictyota.
Discusión
A partir de los resultados encontrados en este estudio se pueden establecer algunos aspectos importantes. La riqueza específica es similar a otras áreas de la región del Cabo (Anaya-Reyna y Riosmena-Rodríguez 1996, Baynes 1999), pero se nota una reducción consistente en la riqueza en relación a otras zonas (Mateo-Cid et al.1993, Paul-Chávez y Riosmena-Rodríguez 2000). Esto muy probablemente está asociado al incremento en la diversidad de peces e invertebrados herbívoros (Montgomery 1980a, 1980b, 1980c, 1980d, Baynes 1999). La variación en el número de especies por localidad (Fig. 2) y el grado de afinidad entre sitios sugiere que existe una gran relación e intercambio entre los sitios con excepción de la zona submareal de Tachuelas que muy probablemente este recibiendo una mayor influencia de la zona del arrecife de Cabo-Pulmo. La proporción de especies por división muestra un porcentaje mayor de algas verdes que pardas, esto no se había registrado en otras localidades en la región (Riosmena-Rodríguez y Paul-Chávez 1997, Paul-Chávez y Riosmena- Rodríguez 2000). Otro aspecto interesante es la presencia de una mayoría de especies anuales en vez de especies esporádicas (Paul-Chávez y Riosmena-Rodríguez 2000), lo que sugiere una mayor estabilidad temporal.
En lo que respecta a las evaluaciones de cobertura se puede determinar que existe una estructura vertical de las poblaciones con un cinturón muy marcado de Gelidium-Jania en la zona intermareal. En el caso de la zona submareal el cinturón lo establece Padina-Dictyota.
El hecho que éstas se vieran afectadas por el sitio, conforme al ANOVA realizado, muestra que existe diferencias marcadas en el reclutamiento en la zona lo que afecta la abundancia de otras especies (como las coralinas o Caulerpa ). Esta forma estructural no se había descrito para la región sur, ya que son pocos los estudios cuantitativos en el área. Las posibles variaciones estacionales estarán regidas por los patrones de temperatura de la zona (Bolton y Anderson 1990) y los eventos de sedimentación locales.
Aunque el estudio fue de tipo espacial, la dominancia de grupos fenológicos anuales muestra que no existe una gran diferencia en la estructura de la comunidad en el año. Las diferencias verticales se deben más a una estrategia de las especies: más esporádicas o anuales en el extremo intermareal, más anuales o perennes en la zonas submareales.
Agradecimientos
El presente trabajo se desarrolló gracias al convenio UABCS-Salinas y Rocha, a quienes agradecemos el auspicio. R. Riosmena-Rodríguez agradece el apoyo de A. Ángeles-Pérez para el presente trabajo.
Resumen
Se caracterizó la flora marina de la zona de Bahía del Rincón -La Rivera por medio de prospecciones intensivas en diferentes localidades a varias profundidades. Además, se realizó un análisis intenso de la literatura y por este mismo medio la determinación de las afinidades geográficas de las especies. En total se encontraron 72 especies, donde la mayor proporción (62%) correspondió a las algas rojas. En el caso de las afinidades biogeográficas se observó que la mayor proporción varió en función de la división, donde para algas cafés las predominantes fueron de naturaleza tropical/ endémica y para las algas rojas fueron las templadas/ cosmopolitas. Espacialmente se encontró una alta similitud entre las zonas someras estudiadas más no así entre las someras y profundas donde existen diferencias significativas. En el caso de la zona intermareal se determinaron diferencias significativas con áreas submareales y entre sitios de la zona somera y profunda basados en las abundancias de los principales taxa. Con base en los resultados se sugiere que existen diferencias verticales en reclutamiento y desarrollo de las poblaciones en relación a su distribución.
Palabras clave :Golfo de California, algas marinas, ecología, Bahía Rincón, desarrollo costero.
Referencias
Anaya-Reyna, G. & R. Riosmena-Rodríguez. 1996. Macroalgas marinas del arrecife coralino de Cabo Pulmo-Los Frailes, B. C. S., México. Rev. Biol. Trop. 44:861-864. [ Links ]
Baynes, W. T. 1999. Factors structuring a subtidal encrusting community in the southern Gulf of California. Bull. Mar. Sci. 64:419-450. [ Links ]
Bolton & Anderson. 1990. Correlation between intertidial seaweed community composition and sea water temperature patterns on a geographical scale. Bot. Mar. 33:447-457. [ Links ]
Brower, J. E., J. H Zar & C. N. Von Ende. 1998. Field and laboratory methods for general ecology. McGraw- Hill. Nueva York. 273 p. [ Links ]
Mateo-Cid, L. E. & A.C. Mendoza-González. 1994. Estudio florístico de las algas bentónicas de Bahía Asunción, Baja California Sur, México. Cien. Mar. 20:41-64. [ Links ]
Mateo-Cid, L. E., I. Sanchéz-Rodríguez, Y. E. Rodríguez- Montesinos & M. M. Casas-Valdez. 1993. Estudio Florístico de las algas marinas bentónicas de Bahía Concepción, B.C.S., México. Cien. Mar. 19:41-60. [ Links ]
Mathieson, A. C. 1989. Phenological Patterns of Northern New England Seaweeds. Bot. Mar. 32:419-438. [ Links ]
Mendoza-González, A. C. & L. E. Mateo-Cid. 1986. Flora marina bentónica de la costa noreste del Estado de Sonora, México. Phycologia 60:414-427. [ Links ]
Mendoza-González, A. C. & L. E. Mateo-Cid. 1994. Algas Marinas bentónicas de Todos Santos, B.C.S., México. Acta Bot. Mex. 29:31-48. [ Links ]
Montgomery, W. L. 1980a. Comparative feeding ecology of two herbivorous damselfishes (Pomacentridae: Teleostei) from the Gulf of California, Mexico. J. Exp. Mar. Biol. Ecol. 47:9-24. [ Links ]
Montgomery, W. L. 1980b. Effects of grazing by the yellowtail surgeonfish, Prionurus punctatus, in the Gulf of California, Mexico. Bull. Mar. Sci. 30:901-902. [ Links ]
Montgomery, W. L. 1980c. The impact of non-selective grazing by the giant blue damsel fish, Microspathodon dorsalis, on algal communities in the Gulf of California, Mexico. Bull. Mar. Sci. 30:290-303. [ Links ]
Montgomery, W. L. 1980d. Marine macroalgae as foods for fshes: an evaluation of potential food quality. Env. Biol. Fish.5:143-153. [ Links ]
Paul-Chávez, L. & R. Riosmena-Rodríguez 2000. Floristic and Biogeographical trends in seaweed assambleges from a Subtropical Insular Island Complex in the Gulf of California. Pac. Sci. 54:137-147. [ Links ]
Riosmena-Rodríguez, R. & L. Paul-Chávez. 1997. Sistemática y biogeográfia de macroalgas en Bahía de La Paz. In J. Urbán-Ramírez & Angeles (eds.). La Bahía de La Paz y su entorno. Investigación y Conservación. UABCS. La Paz. [ Links ]
Riosmena-Rodríguez, R., D. A. Siqueiros-Beltrones & G. Anaya-Reina. 1998. New localities in seaweed distribution in the Gulf of California. Rev. Invest. Cient. Ser. Cien. Mar., UABCS 8:34-58. [ Links ]
Sánchez-Castrejón, E., R. Riosmena-Rodriguez & D. A. Siqueiros-Beltrones. 1995. Nuevos registros de Berkeleya hyalina (Naviculares: Berkeleyaceae) para el Golfo de California, México. Rev. Biol.Trop. 43:323-330. [ Links ]
Sánchez-Rodríguez, I., M. C. Fajardo-León & C. O .Pantoja. 1989. Estudio florístico estacional de las algas en Bahía Magdalena, B.C.S., México. Invest. Mar. CICIMAR 4:35-48. [ Links ]
Zar, J. H. 1996. Bioestatistical analysis. Prentice Hall, Nueva Jersey.662 p. [ Links ]

