











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.53 no.1-2 San José jun. 2005
Floral biology and reproductive system of enantiostylous Senna corymbosa (Caesalpiniaceae)
Cecilia Laporta
Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Lab. Plantas Vasculares. Pab. 2, 4º piso, Ciudad Universitaria, 1428, Buenos Aires, Argentina. Fax:5411-4576-3384; laporta@uni-bremen.de; cecilialaporta@hotmail.com
Received 18-X-2003. Corrected 06-III-2003. Accepted 15-IV-2004.
Abstract
The genus Senna (K.Bahuin) Miller (Cassieae) is represented in Argentina by 35 species and several varieties distributed in temperate, tropical and subtropical regions, and presents a high degree of endemism. Some taxa are used for medicine, animal foraging and ornamental purposes. Floral morphology, phases, rewards, attractants, visitors, pollen, reproductive system, P/O ratio, OCI and ISI indexes of enantiostylous Senna corymbosa were analyzed for morphological androecial differentiation and possible related functional differences between stamens groups. The study was carried out over three consecutive flowering seasons in March of 1999, 2000 and 2001, in two populations near the city of Buenos Aires, Argentina. The species has a buzz-pollination syndrome. The pollinators are Bombus atratus that vibrate the stamens, leading to nothotribic or sternotribic deposition of pollen. P/O ratio and pollen production were high thanks to the precise pollinating mechanism that needs a large delivery of pollen to ensure effective pollen deposition. P/O ratio also indicates that the species is xenogamous, although geitonogamy and autogamy (only induced, not spontaneous)were also recorded. The species is self-compatible. There were differences in hand-pollination treatments between long and medium stamens in fruit set, as well as in vitro differential fertility between their pollen grains. Hence, there are morphological and functional androecium differences: the medium stamens play a nutritional role while the long ones play a reproductive one. These differences are reflected in the breeding system. Moreover, differential fertility and enantiostyly may diminish the effects of self-compatibility by partially reducing the contribution of geitonogamy to selfing. Rev. Biol. Trop. 53(1-2):49-61. Epub 2005 Jun 24.
Keywords: Floral biology, reproductive system, Senna, Cassieae, enantiostyly, buzz-pollination, Bombus.
The genus Senna (K.Bahuin) Miller belongs to the T. Cassieae, ST. Cassiinae (Irwin & Barneby 1982). In Argentina it is represented by 35 species and several varieties distributed in temperate, tropical and subtropical regions and presents a high degree of endemism. Some taxa are used for medicine, animal foraging and ornamental purposes. The majority of the species of the subtribe, comprised of the genera Cassia, Senna and Chamaecrista ,show poricidal anther dehiscence that implies buzz-pollination mediated by large bees. In addition, the androecium is often differentiated into middle length stamens with nutritive anthers (medium), that provide food for pollinators, and long length stamens with pollinating anthers (long), that provide pollen that is deposited on the stigma (Van der Pijl 1954, Faegri and van der Pijl 1966, Proctor and Yeo 1973). The flowers also show enantiostyly; the stylar deflection results in left or right-handed flowers and pollen deposition is produced to one side of the bee s abdomen when it vibrates the stamens. Therefore, the stigma pointing in the opposite direction can only receive pollen that remains on the same side of the insect during a previous visit to the opposite floral morph (Bahadur et al.1990). In Cassia, this floral dimorphism has been interpreted by some authors as a device for promoting cross-pollinations (Todd 1882, Knuth 1906) or for promoting pollination between left and right-handed morphs, increasing geitonogamy (Bahadur et al.1990). Other authors considered enantiostyly as enigmatic (Thorp and Estes 1975).
The present study provides information on floral biology, reproductive system and androecial differentiation of Senna corymbosa (Lam.) Irwin &Barneby. Notwithstanding its medicinal properties and its ornamental and foraging qualities, it has not been widely brought into cultivation (Burkart 1959,1987).Therefore, It is useful to know the reproductive system, the floral biology and the factors that affect effective natural pollination, especially when it depends mostly on insects. We analysed the morphology, floral phases, rewards, attractants, visitors, reproductive system, pollen features and pollen/ovule ratio in order to establish the floral syndrome, the effective pollinators in the study area and the presence of morphological and functional androecial differentiation.
Material and methods
Plant species and study area: Senna corymbosa is a yellow-flowered shrub grown as an ornamental in several areas. It is native of Argentine Mesopotamia, southern Uruguay, the Plate estuary and the neighboring Atlantic shore and coastal lowlands, being the 35 ºS the southern limit of its distribution. The populations in Tucumán and Córdoba provinces are probably secondary (Irwin and Barneby 1982). Flowering phenology, reproductive system, visitor frequencies and pollination events over three consecutive flowering seasons (1999- 2001) were studied in two Senna corymbosa populations. One of them was a natural one placed at the Ecological Reservation "Ribera Norte "(RN) (Bs. As., Acassuso, 34º 29 S, 58º 30 E), consisting of 5 plants. The other population was placed in the Experimental Campus of Universitary City (Buenos Aires, 34º 38 S, 58º 28 E), from seeds of native plants belonging to the Ecological Reservation "Costanera Sur "(CS) (Bs.As.), consisting in 10 plants. The study area comprises the southern limit of distribution of the species.
Flowering phenology and morphology: General morphology was studied from fresh and fixed material (FAA) using a stereosmicroscope. Photographs with a Scanning Electronic Microscope (Jeol Scanning Microscope JSM II) were taken to analyse stigma and anther morphology. Coloring patterns of the corollas were identified with standard color charts (Kelly 1965).
Floral phases: From the begining of anthesis, stigma receptivity was tested taking as a positive result the bubbling in presence of hydrogen peroxide (Osborn et al.1988), as well as the anthers changes in order to distinguish floral phases. The evolution of the anthesis was also followed with hand lenses and the relative frequency of each floral phase in two plants of both populations was registered (n=60 flowers (fl)/plant). It was also calculated the approach ratio between right to left handed flowers, through three random countings in three samples of each population (n=180 fl).
Black and white photographs were taken with a Kodak 18A filter to determine the reflection pattern of the flowers and the presence of reward guides in the UV spectrum of vision of pollinators.
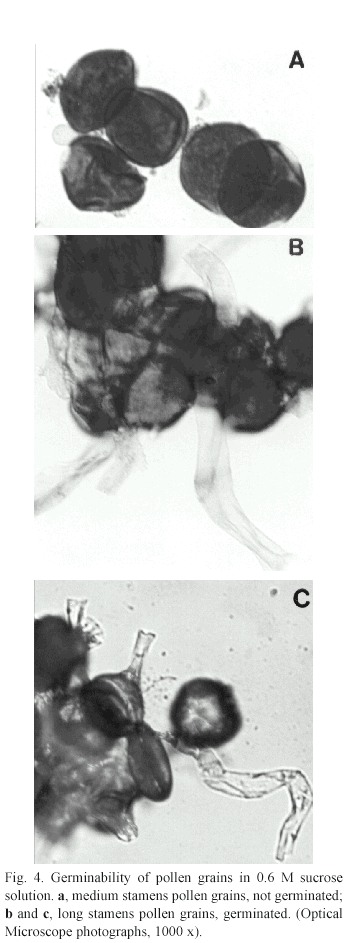
Pollen analysis,production and pollen/ ovule ratio: Pollen viability was tested (Greissl 1989) and stained pollen grains were observed under epifluorescence microscope with a UV- filter (380-425 nm). Pollen tube growth of each type of stamens was analysed in 0.2M, 0.4M and 0.6M sucrose concentrations (Dafni 1992). The existence of starch, lipidic and proteic reserves was tested by staining fresh pollen grains with iodine solution,sudan IV and picric acid, respectively. The number of pollen grains per anther and P/O (pollen/ovule ratio) for both enantiostylious right and left handed morphs and both long and medium stamens were calculated (Cruden 1977).
Reproductive system: During March of 1999, 2000 and 2001, 70 buds of three different plants of each population, prior to anthesis, were bagged with tulle bags. When the flowers started to open, they were unbagged and hand-pollinated using pollen from long and medium stamens, contacting the stigmas five times. In 2001, hand-pollination experiences were also performed with a diapason in population two. It vibrates at the same frequency as bumblebees do.The vibrating diapason contacted the pistile, which had half of an open anther on the stigma.Two kind of tests were carried on over each flowering season:
a)Spontaneus. (1) Autogamy, ten flowers were bagged and not hand-pollinated. (2) Free pollination, ten marked buds were offered to foraging visitors and bagged after 72 hr to avoid damage by frugivores.
b)Induced. (1) Autogamy, 20 flowers were hand-pollinated, half of them with pollen of long stamens and the other half with pollen of medium ones, of the same flower.(2)Geitonogamy:the same as autogamy but with pollen of the same plant. (3) Xenogamy: the same as autogamy but with pollen of three different plants.
All the flowers were bagged again after each treatment to avoid damage by frugivores. After 30 days fruit set (number of fruits per number of flowers) and seed set (number of seeds per fruit) were recorded. ISI (Self-incompatibility Index) (Dafni 1992) as well as OCI (Outcrossing Index) (Cruden 1976) were calculated.
Floral visitors, pollinators and pollen load: The spectrum, frequency and behavior of floral visitors were recorded on 20 previously marked flowers in two plants of each population. Observations were conducted over 1999 and 2000 flowering seasons during five consecutive days, in March of each year, from the begining of the anthesis (07 hr) until visits were stopped completely (15 hr). Herbarium vouchers of the flowering vegetation on a 1km2 surrounding area were collected, determined and their pollen characterized after having been acetolised (Erdtman 1952). Both plant organism and palynological vouchers were deposited at the BAFC Herbarium. Pollinators were identified among all the floral visitors by regarding their behavior, their successful contact with the reproductive cycles of the flowers and their pollen load (Genise et al. 1990). One specimen of each species captured was deposited at the Museo Argentino de Ciencias Naturales "B.Rivadavia".
Within each plant, pollinators flight movements were observed to determine if they discriminated among floral types or, alternatively, randomly moved between them, as well as among long and medium stamens. Observations were conducted for four days in two plants of each population, between 10:45 and 11:15 hr, during the highest pick of activity of the insects. Their behavior in each flower was followed until they completely left the plants. It was registered if they buzzed only the long stamens, only the medium ones, first the long and afterwards the mediums or vice versa. Temperature and relative humidity were registered daily with a digital hygrothermometer.
Statistical analysis: Student t-test was used to determine if there were differences between Right handed-Left handed morphs and long-medium stamens with respect to the pollen grain production. Chi square tests were performed to determine whether insects moved randomly among floral morphs, and to check the probabilities of an 1:1 ratio between both enantiomorphs in the population.
Results
Flowering phenology and morphology: Senna corymbosa is a shrub that reaches from 1 to 3 m height. At the places of study it blooms from October to April, with two flowering peaks in November and March. Inflorescences are composed of racemes that are 8-10 flowered, each day 1-2 flowers are open and remain open for 2 days. Floral diameter is about two to three cm, with five subequal and imbricate petals. The biggest and ourtermost is a dish-like petal and the two smallest are posterior and partially protect the anthers and gynoecium (Fig.1c). The flowers are perfect, without nectar, apparently odourless, pollen being the main reward.Coloration of corolla and anthers are primary attractants (Fig.5).
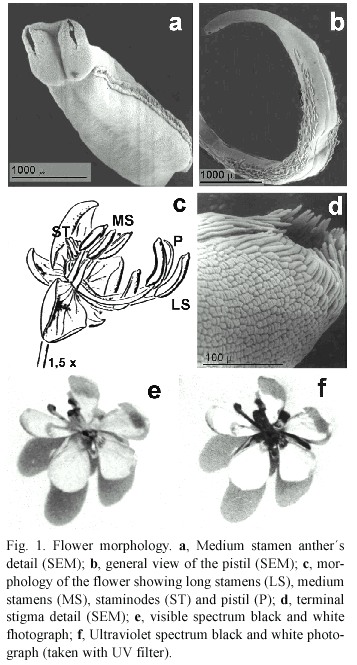
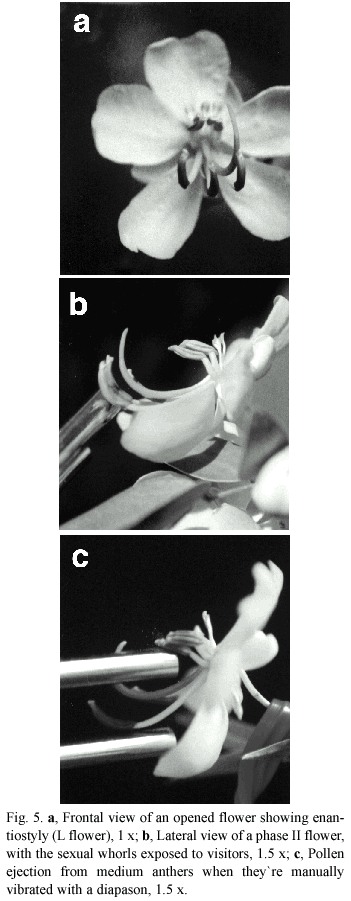
Androecium. The anthers are appendiculated and dehisce by apical slits which open or close according to ambient humidity (Fig.1a). The three uppermost stamens have sterile anthers. There are three types of fertile stamens, arranged in two groups. The first is a central group of four stamens with short filaments and large "feeding " anthers, directed with the apertures towards an approaching floral visitor (medium). The second group comprises three curved stamens with long filaments and also large ´pollinating ´ anthers placed in front of the ourtermost dish-like petal (long) (Fig.1c). The two lateral stamens of this group of three are inclined each one to the right and to the left of the midline of the flower. The third stamen is centric, abaxial, lowermost and curved whose filament is shorter than those of its neighbours, reaching half of the height of them (Figs.1c,5a).
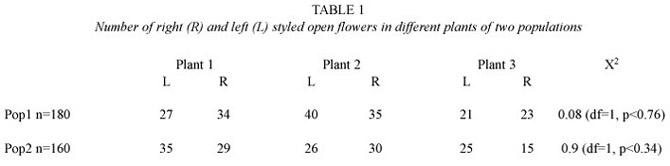
Gynoecium. The unique and whole carpel is deflected and the style is bent down. The stigma is terminal, minute, dry and chambered (Fig. 1d). The sickle-shaped pistil emerges between the bases of the two longest stamens and above the low remaining one. It projects to the right or to the left of the middle line of the flower, and is curved upwards (Fig.1b). It s less than twice as long as the longest stamens (Fig.5b). According to the direction of deflection of the pistil base, the flowers are right-styled (R) or left-styled (L) (Fig.5a). Those flowers borne in the same node show somatic inverse enantiostyly. Right and left handed flowers are also borne in the same raceme in different nodes but a regular pattern in their sequence in the inflorescence could not be discerned. The enantiomorphs approached 1:1 ratios in random countings of newly open flowers (Table 1).
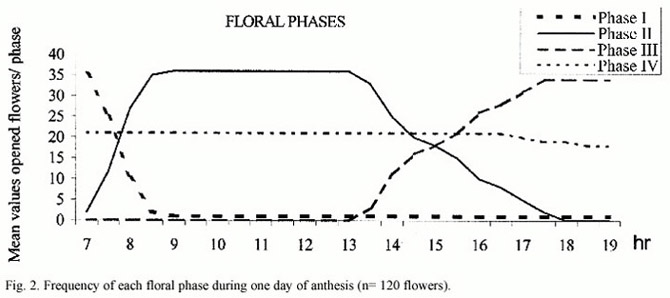
Floral phases: The anthesis exhibits four floral phases and lasts two days. Phase I is the shortest in time, from 06.30 to 08.30 hr, showing a partially open flower, orange-yellow coloured with positive stigmatic receptivity. At 08.30 hr in the morning, almost all the flowers reached phase II, in which they are completely opened, the stigmatic receptivity is positive and the sexual whorls are exposed to visitors (Fig.5a,b). The anthers of the two types of stamens show different colors: yellow for the medium stamens and brownish yellow for the long ones. In this phase the anthers present the maximum amount of pollen, they are turgid and remain stiff up to the begining of phase III, although their pollen load diminished considerably after being forraged by insects. The shift between phase II and III occurs after 14.00 hr. Anthers, almost empty, became darker in the early afternoon, changing their color from yellow to pale brown. The corolla looses turgency and becomes lighter coloured, the stigmatic receptivity is still positive. The flower remains on the plant even if it has not been fertilized. During the second day of anthesis, it starts phase IV. The stigmatic receptivity is negative, the color of the corolla turns to light orange and the anthers became drier and darker. Relative frequency of floral phases changes through the day, with phase II the predominant one between 09.00 and 14.00 hr.(Fig.2). Black and white photographs show a spectral contrast in the open flower between the center and the margins. It is evident in the UV range, visible to the insects (Fig.1a ,b). It may act as a pollen guide.
Pollen analysis and production:Viability of pollen grains was about 98% and there were not differences between both groups of stamens, nor in pollen grain reserves (proteins and carbohydrates). On the other hand, in vitro germinability showed strong differences between long and medium stamens (Fig.4) and also between the different sucrose concentrations tested: only pollen grains of long stamens did germinate, mostly at 0.6 M concentrations of sucrose. Anthers of long stamens carried higher number of pollen grains (aprox.60%) than those of medium ones (Table 2).There were not found significant differences between R flowers and L flowers in long stamens' pollen grain production (t=0.25,8 df,p=0.995)as well as in medium stamens pollen grain production (t=0.086,8 df,p=0.995).There were not also found significant differences between pollen grain production of long stamens and medium stamens of R flowers (t=2,34,8 df,p=0.995) nor even of L flowers (t=1,819,8 df,p=0.995).
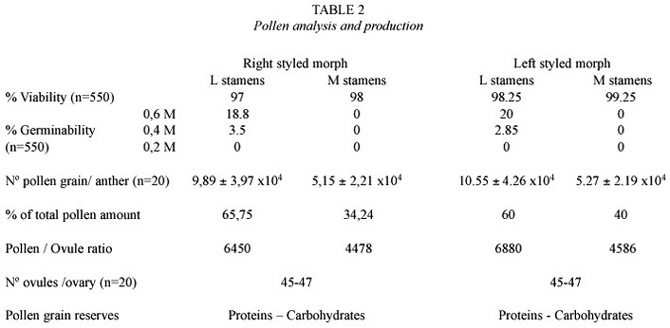
Reproductive system and P/O ratio: Hand pollination experiences showed that successful crosses were carried out with pollen grains of medium and long groups of stamens. The species is self compatible,since autogamy and geitonogamy were registered.There were differences on fruit set and seed set between free pollination and artificial one as well as between medium and long stamens in fruit set (Table 3).The OCI index indicates that the species is adapted for outcrossing,mostly self compatible and pollinator demanding. ISI index shows self-compatibility.P/O ratio was extremelly high for both group of stamens, indicating obligate xenogamy as the main reproductive system (Table 2).

Visitors: The main visitors and successful pollinators were individuals of Bombus atratus Franklin. They vibrated the medium stamens during their visited and receive the pollen on the ventral side of their bodies, on the inner side of their legs and on their mouth parts (Table 4). The vibration of the medium anthers is transmitted by the bees body to the long stamens and causes the pollen to be ejected and deposited on the ventral part of their bodies and on the back or on the sides of them (pleurotribic deposition). Although this pollen is brushed off by the bees,it is only with difficulty collected by them. The pollen is deposited on the bees body just at the place where the stigma may touch or approximates closely during vibrations (Fig.6). They are transmitted to all the anthers, causing the ejection of pollen from all of them (Fig.5c).

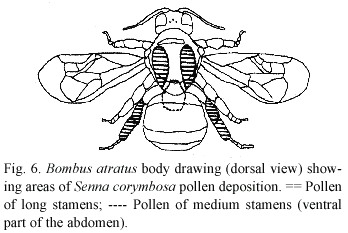
In fact, a direct contact of the stigma and a bees body could only rarely be seen, whereas close approximation was more common. The largest amount of pollen was found deposited only on the ventral side of the bees body and it also comes close to the stigma, even the pollinator touches it with its ventral body side. Then, individuals of Bombus atratus apparently also pollinate this species with pollen from the ventral side (sternotribic deposition) when entering or leaving the flower. The pollination may occur with pollen from both the medium anthers and the three longest ones. Pollen load observed in each part of the bee's and other visitor's bodies is shown in Table 4.
Individuals of Apis mellifera were also observed visiting the flowers. Its relatively small size does not allow them to contact or even approximate the stigma. They act as pollen thieves. Wasps were observed exploring buds. Ants explored buds and also open flowers:they entered them and collected pollen that remained on the petals due to previous visits of other buzzing visitors that can extract pollen. They also explored stipitate glands placed on the rachis and branches in general.
It was seen that insects do not discriminate between floral morphs but they move randomly (c 2 =2.4,df=1,p<0.12 in March 1999 and c 2 = 0.8,df=1,p<0.37 in March 2000)and that they predominantly buzz the median stamens or the median first and the long afterwards (Table 5).

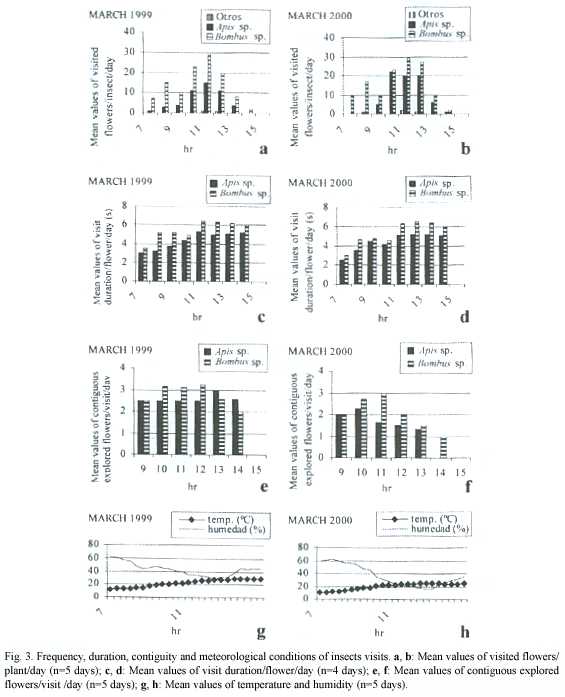
Visitor s activity and meteorological conditions: According to the graphics, a peak of maximum activity is registered between 11.00 and 12.00 hr. At this time was observed the highest frequency of visits and contiguous flowers explored during both 1999 and 2000 study seasons (Fig.3a,b,e,f). A previous peak of activity between 08.00 and 09.00 hr can also be seen. The duration of each visit (the time the visitor remained in the flower) was longer in the earlier peak than in the later one (Fig.3c, d). The number of contiguous flowers visited per plant is higher in the second peak of activity, that shows an increase of temperature and a decrease of humidity, than in the first one. (Fig.3g,h).
Discussion
Results of floral morphology, phases and visitor behavior allowed to conclude that Senna corymbosa presents a floral syndrome of melitophily and, more specifically, a buzz-pollination syndrome (Dulberger 1981,Buchman 1983). It comprises some floral features shown by the species treated, like the occurrence of nectarless flowers, poricidal dehiscense of anthers, minute terminal stigmas, curved styles and release of pollen when anthers are vibrated by insects (Buchman 1974).
The main visitors are bees able to vibrate while "milking" the stamens (medium ones). The low diversity of visitors observed presumably is due to the study site location, at the southern limit of the distribution of the species. Between the captured individuals, those of Bombus attratus presented pollen of S.corymbosa in the dorsal and ventral part of the thorax. The frequency of their visits was maximum and their large size allowed them to partially contact or approximate the stigmas when they vibrated the medium stamens. Electrostatic forces play an important role in pollen transfer via insects and enable pollen to jump from anther to bee and from bee to stigma (Corbet et al.1982,Buchman 1983).
Small bees, like Apis melifera individuals, do not effectively pollinate the flowers because of their small body size relative to the flower features.The observations revealed that often there was no real contact of the bees body with the stigma and/or the opening anthers.
Pollinating agents are necessary not only for carrying pollen and its later placement on the stigmas, but also for release of pollen from the anthers. Since insects buzz predominantly the medium stamens, a functional difference between them can be established: the long stamens produce pollen for the fecundation while the medium ones produce pollen that is mainly used by the visitors to nurture their larvae. Present results show that the medium stamens produce viable pollen, with reserves but can not germinate in vitro (Table 2). These kind of anthers are known as "feeding" ones. Regarding the insect's behavior, the orientation of the anthers leads to deposition of pollen from the "feeding "anthers on the ventral parts of the insect and pollen from the "pollinating" anthers on its back or side of the body. These last anthers contain the largest amount of total pollen produced (Table 2), accounting for the degree of precision of the pollination mechanism, that needs a large delivery of pollen. It was also seen in other self-compatible and enantiostylous species of the genus Cassia (Dulberger 1981) and Cyanella (Dulberger and Ornduff 1980).
In spite of the fact that medium stamens do not show in vitro germination of pollen grains, they do germinate in natural conditions, regarding for the sucessful hand-crosses obtained from pollen of both median and long anthers (Table 3). In Cassia auriculata mild incompatibility was reported (Bir Bahadur et al.1979), but it was later regarded as a differential fertility of pollen from long and medium stamens (Dulberger 1980). However, there are differences on fruit set when medium or long stamens are used as pollinating agents. It may suggest that this extreme specialization of the pollinating mechanism would evolve in a differential fertility of pollen grains that could dampen or diminish the effects of the self-compatibility, since pollinators also contact the stigmas with the ventral part of their bodies, where pollen of the medium stamens is mostly carried. Although these grains did not germinate in-vitro but in natural conditions, it can be suggested that pollen tubes from both long and medium stamens show a different growth rate. Hence, if both kind of pollen grains are transferred to the stigma, those of long stamens will reach the ovules faster than those of medium stamens, implying an adaptative advantage to the first ones. Then, the functional differences of the stamens are not quite fully reflected in the breeding system. The differences in the orientation of the style (enantiostyly) determine that the pollen deposited in the superior part of the thorax can only be captured by the style that presents the same position. It means that the insect may has previously visited the opposite floral morph to assure an effective pollination, transferring nototribically pollen from the long anthers to the stigmas, as it was seen in other Cassiinae species (Dulberger 1981, Gottsberger 1988, Fenster 1995). But even if the pollinator has previously visited the same floral morph, it will be able to pollinate because long anthers surround it from both sides of its body. Pollen grains may be deposited on the right and the left side simultaneously, since vibrations are transmitted to all the stamens, causing the ejection of pollen from all the anthers. In a recent study on Chamaecrista fasciculata, one of the genus of the subtribe Cassiinae, it was found that there were small differences in outcrossing rate between enantiostylous and non-enantiostylous plants, artificialy created (Fenster 1995). It may also suggest that, even when enantiostyly seems to be a way to avoid self-fertilization, its role is not clear enough in those cases when is associated with self-compatible systems. The fact that insects do not discriminate between floral morphs diminishes in half the probability of visiting the opposite floral morph. Present results allow to conclude that the genetic effect of pollination between the same floral morph in a plant is the same that of autogamy. It is clear that xenogamy is not specifically promoted by this enantiostylous mechanism, since all the plants of a population present both floral morphs in an approached ratio 1:1. In Cyanella lutea (Tecophilaeaceae), a self-incompatible species, enantiomorphy together with a relatively small number of flowers concurrently open on a plant, presumably promotes outcrossing (Dulberger and Ornduff 1980). In Solanum rostratum it was found that the ratio between geitonogamy to xenogamy is dependent on the flight pattern of the bee and the presence of a relatively small and equal number of R /L flowers simultaneously open on a plant (Bowers 1975).
The reproductive studies and the P/O ratio indicate that the species is xenogamous, as confirmed in field experiencies. But geitonogamy and autogamy were also registered, showing that the species is self compatible. The OCI and ISI indexes also support self-compatibility. The P/O ratio is extremelly high for the species treated compared to those estimated by Cruden (1977) for several entomophilous-xenogamous plants, even if it is calculated only from pollen of large anthers. Some explanations are possible: the pollen produced may serve as nutritive source as well as reproductive, so it must be high and specially when pollinators are large bees which needs large amounts of pollen. Another reason could be that, regarding the degree of precision of the pollination mechanism, it needs a large delivery of pollen to assure an effective pollen deposition. In the case of autogamy, it was succesfully produced only when induced, not spontaneous. It indicates that in the abscense of an agent that vibrates the anthers no pollen is discharged, showing a strong dependence on insects for pollination events. And it is also coincident with the OCI index, that indicates a pollinator- demanding species (Cruden 1976). Its necessary to point out that this method does not discriminate the pollen origin.
There were differences of fruit set and seed set between free and hand pollination. The lower numbers produced by hand pollination treatments may be the result of the random transfer of medium stamens pollen grains vs. long stamens pollen grains. In natural conditions, the flowers are visited repeatedly during the morning, promoted by "dummy" anthers that remained coloured and stiff even when they are empty. Pollen grains placed in stigmatic chambers by insects visits may be higher than when placed there through five or six manual events, as it was seen when free and artificial pollinated stigmas where observed under hand lenses (data not shown). Preliminary, present results indicate that the reproductive treatments performed with diapason (vibrating the stily while being pollinated) showed a higher number of fruit and seed set than those performed without it. It suggests that the style may play an important role in pollen grain germination. Its movements may also promote the placement of pollen grains in the stigmatic chamber that contains the stigmatic exudate allowing their germination.
According to the graphics, a peak of maximum visitors activity occurs between 11.00 and 12.00 hs. It is coincident with the greatest abundance of open flowers, mostly in the second floral phase,that show positive stigmatic receptivity (Fig.2).Theres a previous activity peak between 8.00 and 9.00 hr, when flowers begin the second phase. During the earlier peak xenogamy is favored, while during the latest one geitonogamy is favored. The "dummy" anthers play an importante role in this way. Early in the morning, the duration of the insect visits is shorter than at midday (Fig.4c,d) because the anthers are full of pollen grains and insects can easily collect them,visiting also fewer contiguous flowers (Fig.4e,f) and promoting the visit to others plants flowers. At midday, they have to visit more flowers to collect the same amount of pollen (Fig.4e,f), remaining more time in each flower (Fig.4c,d) and visiting more flowers of the same plant. After midday, the activity decays because of the presence of empty anthers (floral phase III) and because of water-stress and thermorregulation problems affecting bees (Fig.4g,h and Fig.2). The activity peaks are also coincident with an increase of temperature and a decrease of humidity, that allow a proper ejection of the pollen cloud by buzzing (Fig.4a,b,g,h). Phenological studies of some Cassiinae species (Delgado Salinas and Souza Sánchez 1977, Gottsberger 1988) demostrated that, even when the species flowering is mostly produced during the rainy season, bees approach flowers only during dry weather. Presumably, those visitors that collect pollen actively but dont pollinate (Apis sp.) produce a decrease of pollen that forces the effective pollinators (Bombus atratus ) to visit more flowers searching for pollen, increasing in this way the geitonogamy /xenogamy.
Whether enantiostyly promotes the increase of pollen dispersal phase of cross-pollination (Fenster 1995), reduces the probability of damage to a fragile gynoecium during buzz pollination (Dulberger 1981) or does not promote outcrossing itself, is being studied. But enantiomorphy in Senna corymbosa seems to diminish the effects of self-compatibility by partially reducing the contribution of geitonogamy to selfing.
Resumen
Se analizaron la morfología y fases florales, recompensa, atractivos, visitantes, polen, sistema reproductivo, índices OCI e ISI y relación polen/óvulo de la especie enantiostílica Senna corymbosa. El objetivo fue estudiar su biología floral y determinar la posible presencia de diferenciación morfológica en el androceo. Era importante saber si las diferencias morfológicas reflejaban diferencias funcionales entre los grupos de estambres. El estudio se llevó a cabo durante tres estaciones de floración consecutivas en marzo de 1999,2000 y 2001,sobre dos poblaciones presentes en zonas protegidas de la ciudad de Buenos Aires, Argentina. La especie presenta un síndrome de polinización vibrátil (buzz-pollination).Los polinizadores son individuos de Bombus atratus, capaces de vibrar los estambres y extraer polen, el que se deposita nototríbica y/o esternotríbicamente. La relación P/O fue extremadamente elevada, así como la producción de polen. Esto se relaciona con el comportamiento de los visitantes y con la alta precisión del mecanismo de polinización, que requiere una gran cantidad de polen para asegurar su efectiva deposición en el cuerpo del insecto. La relación P/O indica que la especie es xenógama, aunque se registraron también geitonogamia y autogamia inducidas. La especie, por lo tanto, es autocompatible. Se encontraron diferencias en los tratamientos de polinización manual entre estambres largos y medianos en cuanto al establecimiento de frutos, así como una fertilidad diferencial in vitro entre sus tipos polínicos. Existen, así, diferencias morfológicas y funcionales en el androceo: los estambres medianos tienen un papel nutricional mientras que los largos cumplen un papel reproductivo. Estas diferencias se reflejan en el sistema de cruzamiento. Más aún, la fertilidad diferencial y la enantiostilia disminuyen los efectos de la autocompatibilidad reduciendo parcialmente la contribución de la geitonogamia a la endogamia.
Palabras clave: Biología floral, sistema reproductivo, Senna , Cassieae, enantiostilia, polinización vibrátil, Bombus .
References
Bahadur B., P. V. Kumar & N. P. Reddy. 1979. Enantiostyly in Cassia L. (Caesalpinioideae) together with SEM studies of pollen and its possible significance. In Reproduction in flowering plants. Abstr. Int.Symp. Christchurch,New Zealand. [ Links ]
Bahadur B., A. Chatuverdi & N. Rama Swary.1990. S.E.M. studies of pollen in relation to enantiostyly and heteranthery in Cassia (Caesalpinaceae). Current perspectives in Palyn. l Research, J. Palynol. 1990- 1991:7-22. [ Links ]
Bowers, K. A. W. 1975. The pollination ecology of Solanum rostratum (Solanaceae). Amer. J. Bot. 62:633-638. [ Links ]
Buchmann, S. L. 1974. Buzz pollination of Cassia quiedondilla (Leguminosae) by bees of the genera Centris and Melopoma. Bull. South Cal. Acad. Sci. 73:171-173. [ Links ]
Buchmann, S. L. 1983. Buzz pollination in angiosperms, p. 73-113. In C.E. Jones & R. J. Little (eds.).Handbook of Experimental Biology, Scientific and Academic Editions, New York, NY. [ Links ]
Burkart, A. 1959. Leguminosas, p.463-465. In L. R. Parodi (ed.).Enciclopedia Argentina de Agricultura y Jardinería. Acme. Buenos Aires. [ Links ]
Burkart, A. 1987. Leguminosae, p.509-518. In Secretaría de Agricultura, Ganadería y Pesca (ed.).Colección Científica del Inta, Tomo VI, parte 3 ª. Flora ilustrada de Entre Ríos. Buenos Aires. [ Links ]
Corbet, S. A., J. Beament & D. Eisikowitch. 1982. Are electrostatic forces involved in pollen transfer? Plant Cell Environ.5:125-129. [ Links ]
Cruden, R. W. 1977. Pollen ovule-ratios: a conservative indicator of breeding systems in flowering plants. Evolution 31:22-46. [ Links ]
Delgado-Salinas, A. & S. M. Sánchez. 1977. Biología floral del género Cassia en la región de los Tuxtlas, Veracruz, México. Bol. Soc. Bot. Mex. 37:5-53. [ Links ]
Dulberger, R. & R. Ornduff. 1980. Floral morphology and reproductive biology of four species of Cyanella (Tecophilaeaceae). New Phytol. 86:45-56. [ Links ]
Dulberger, R. 1981. Biology of Cassia didymobotrya and C.auriculata. Am. J. Bot. 68:1350-1360. [ Links ]
Erdtman, G. 1952. Pollen Morphology and Plant Taxonomy. Angiosperms. (An introduction to Palynology). Almquist & Wicksell, Stockholm. [ Links ]
Faegri, K. & L. Van der Pijl. 1971. The Principles of Pollination Biology.Oxford. [ Links ]
Fenster, C. B. 1995. Mirror image flowers and their effect on outcrossing rate in Chamaecrista fasciculata (Leguminosae). Amer. J. Bot.82:46-50. [ Links ]
Genise, J., R. Palacios, P. Hoc et al. 1990. Observaciones sobre la biología floral de Prosopis II. Fases florales y visitantes en el Distrito Chaqueño Serrano. Darwiniana 30:71-85. [ Links ]
Gottsberger, G. & I. Silberbauer-Gottsberger. 1988. Evolution of flower structure and pollination in Neotropical Casiinae (Caesalpinaceae) species. Phyton 28:293-320. [ Links ]
Greissl, R. 1989. Vitality analysis of monadic and polyadic pollen grains using optical contrast-fluorescence microscopy. Sci. Tech. Info. 15:180-184. [ Links ]
Irwin, H. S. & R. Barneby. 1982. The American Cassiinae. A synoptical revision of Leguminosae tribe Cassieae subtribe Cassiinae in the New World. Mem. NY Bot. Garden 35:455-918. [ Links ]
Kelly, K. L. 1965. ISCC-NBS color-name charts illustrated with centroid colors. Suppl. to Nat. Bur. Standards Circ.553.U.S.Government, Washington. [ Links ]
Knuth, P. 1906. Handbook of flower pollination.Vol.1. Clarendon,Oxford. [ Links ]
Osborn, N. M., P. G. Kevan & M. Lane. 1988. Pollination biology of Opuntia polycantha and Opuntia phaecantha (Cactaceae) in Southern Colorado. Plant Syst. Evol.159:85-94. [ Links ]
Proctor, M. & P. Yeo. 1973. The pollination of flowers. Collins, London. [ Links ]
Thorp, R. W. & J. R. Estes. 1975. Intrafloral behavior of bees on flowers of Cassia fasciculata . J. Kans. Entom. Soc. 48:175-184. [ Links ]
Todd, J. E. 1882. On the flowers of Solanum rostratum and Cassia chamaecrista. Amer. Nat. 16:281-287. [ Links ]

