










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 n.1-2 San José Jun. 2005
Secondary stem anatomy and uses of four drought-deciduous species of a tropical dry forest in México
Alejandra Quintanar Isaías 1 ,Mariana Velázquez Núñez 1 ,Fortunato Solares Arenas 2 ,Carmen de la Paz Pérez Olvera 1 & Alfonso Torre-Blanco 3
1 Departamento de Biología, Universidad Autónoma Metropolitana-Iztapalapa. Av. Michoacán y Purísima, Colonia Vicentina. C.P. 04690.
2 Campo Experimental, INIFAP, Zacatepec Morelos.
3 Laboratorio de Bioquímica, Facultad de Ciencias, UNAM, México. Fax (55)5804-4688; aqi@xanum.uam.mx
Received 06-VI-2003. Corrected 18-XI-2004. Accepted 10-XII-2004.
Abstract
Wood and bark anatomy and histochemistry of Acacia bilimekii Humb. & Bonpl., Acacia cochliacantha Mcbride., Conzatia multiflora (Rob) Stand. and Guazuma ulmifolia Lam.are described from stem samples collected in a tropical dry forest (Morelos,Mexico). Enzyme activities were tested in tangential, radial and transverse cuts of fresh material. Histochemistry and stem anatomy were studied on similar cuts previously softened in a solution of water-glicerol-PEG. Our results show that the anatomical patterns of bark and wood, as well as the histochemical patterns and specific gravity, are influenced by water accessibility and climate; these patterns could guarantee mechanical and anti-infection strategies to support extreme conditions. Enzyme cytochemistry reveals biochemical activities probably related to lipid utilization routes for the lignification processes and for synthesis of extractives; these results suggest that the formation and maturation of woody tissue is very active at the beginning of the rainy season. These species are widely used by the local population. Traditional uses include firewood, dead and live fences, fodder, construction, supporting stakes, handcrafts, farming tools, extraction of tanning products, and medicine. There is no relationship between use and abundance. Alternative uses are proposed according to a density index. Rev. Biol. Trop. 53 (1-2): 29-48. Epub 2005 Jun 24.
Key words: Bark and wood anatomy, histochemistry, enzyme cytochemistry, uses, drought-deciduous species, dry forest, Mexico.
Tropical dry forests are ecosystems widely distributed throughout Mexico. However, in spite of their wide distribution, no species of these ecosystems is used by the lumber industry. Up to the seventies, these ecosystems covered about 8% of this country (Miranda and Hernández-X.1963, Flores et al.1971, Rzedowsky 1978) but, by the mid nineties, at least 45% of the former 110 000 km2, were completely spoiled (Anonimous 1975, 1994, Boyás et al . 1993a).
The State of Morelos possessed ample stretches of tropical dry forests but these plant communities have been exploited by the local population, with little effort directed to their regeneration, or converted to other uses. At present, only 670 km2 of these ecosystems survive in this State with a moderate degree of conservation and 1100 km2 with a high degree of alteration (Solares 1997a).
From about 400 useful plant species listed in Morelos, 80% grow in tropical dry forests, including the ten most important species, all of them trees (Boyás et al.1993a,1993a,b). There is an intense exploitation of many plant species from these ecosystems, but only for traditional uses. In the State of Morelos rural communities have a rich ethnobotanical knowledge and many people, directly or indirectly, make their living on it, for this reason is important to implement conservation projects and recovery of degraded surfaces. In a recent report, Solares (1999) considers that about 45% of the species present in these ecosystems have a potential use as timber and 75% have other potential uses. For instance, the cuachalalate (Amphipteyigium adstringens Schiede.ex Schlecht.) an the cirián or cuatecomate (Crescentia alata H.B.K.) are wild resources now exploited on a sustentable basis (Solares 1997b,1999).
A.bilimekii, A.cochliacantha and G.ulmifolia are species typical of the tropical dry forest of Morelos, already used by the local population in several ways. The Acacia species are used for construction, poles, dead fences and as firewood. G ulmifolia is abundant in this ecosystem and is the main source of energy in this region as firewood. C. multiflora, on the other hand,is only marginally used but is the most abundant species in these communities (4700 trees/km2 )and has a high potential as an ornamental tree (Solares 1997a).
Anatomical descriptions of secondary phloem structures (Esau 1965, Fahn, 1990), revisions of their nomenclature and directions for the description of these tissues (Trokenbrodt 1990, Junikka 1994), as well as a proposal of classification based on their anatomical features (Furuno 1990) have been published.In addition to anatomy, the chemical composition of secondary phloem of Pinus pinaster has been studied (Nunes et al.1996) and, more recently, the anatomical and histochemical features of these tissues have been described in several species from a tropical dry forest (Paredes et al.2001).
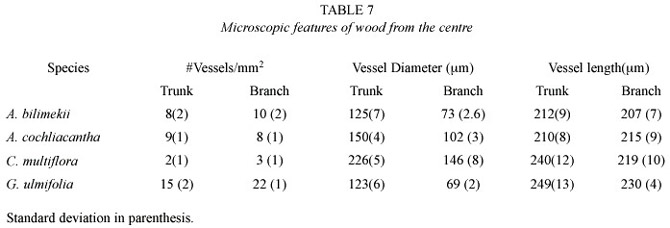
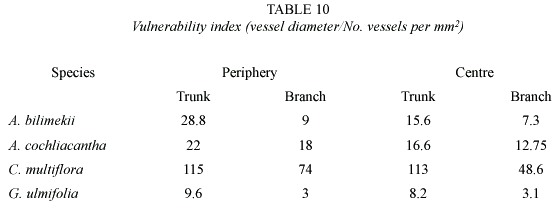
The study of the anatomical, histochemical and enzymatic features of the stem and primary branches contribute to the understanding of survival strategies of plants in different climates. Comparative studies of the anatomy of stems and branches can describe dimensional trends in cellular elements. In particular, vessel diameter and number of pores per mm2 are characters useful for calculating the vulnerability index; this, in turn, is related to the resistance of functional vessel elements to break the water columns flowing in the sapwood at a particular moment (Sobrado 1997). These studies, in addition, can contribute to a better understanding of the structure and properties of the wood.
In this paper we attempt to integrate anatomical, histochemical and enzymatic data of the wood and bark of A.bilimekii, A.cochliacantha, C.multiflora and G.ulmifolia; we also contribute information on the specific gravity of xylem and on the distribution and uses of these species.
Material and methods
Features of the study sites .The tree samples came from three sites of tropical dry forest in the State of Morelos: a) Barranca Honda; 9.5 km South West from Yautepec,1450 mosl, NE hillside exposure; 35% slope; 99° 6 55 W longitude. b)Cerro de la Tortuga in Tetelpa; 1.5 km NorthWest from Zacatepec, 1000 mosl, SW hillside exposure, 20% slope; 99° 12 12 W longitude. c). Valle de Vázquez; 14 km SE from Jojutla de Juárez, 1050 mosl, NE hillside exposure, 10% slope; 99° 4 25 W longitude. The climatic and soil features proposed by Boyás (1991) in his ecological regionalization for the state of Morelos that characterize the sites of study are: subhumid warm climate, ridge, lime stone and rendzina soil.
Barranca Honda, Valle de Vázquez and Tetelpa are small rural communities of Mestizo people dedicated to agriculture and livestock, with a population of 700, 1100 and 600 respectively (Anonimous 1991). Between 10 to 15% of the population is illiterate. The economically active population was 21% (199), 22% (233) and 32% (166) respectively (Anonimous 1995).
Mature, healthy trees of each species were collected from the three sites in May, at the beginning of the rainy season. The precipitation during this month was 55 mm and the average temperature was 26 ºC, acording to data obtained from the Meteorological Station in Huautla, Morelos.
Anatomy studies. The trees were sampled to obtain wood from the centre and periphery of principal stem at 1.30 m (Breast Height Diameter-BHD)and from the first branches next to principal stem. Cubes of 1.5 x 1.5 cm were prepared and softened in a mixture of equal parts of water-glicerol-PEG. Cross, tangential and radial sections were obtained. Dissociated fibres and vessels were prepared following Johansen s protocol (1940). This cell material was mounted in glycerine-gelatine stained with safranine or Bismark brown.
The IAWA Committee s (Anonimous 1989)and Chattaway s wood cell size clasification (1932) were followed to describe wood microscopic features, and Kribs (1968) for describing rays. Each description includes specific gravity data obtained after dividing dry weight (kg) by weight of green volume (kg). A comparative table of vulnerability index values (average vessel diameter/ # of vessels in a cross section of 1 mm2 )is also given.
Trokenbrot s nomenclature (1990) was followed for bark description. Mean and standard deviation of wood and bark measurable cell elements were calculated to p<0.05.
All botanical samples were registered and stored in the Metropolitan Herbarium (UAMIZ) of Universidad Autónoma Metropolitana, México (Table 1).
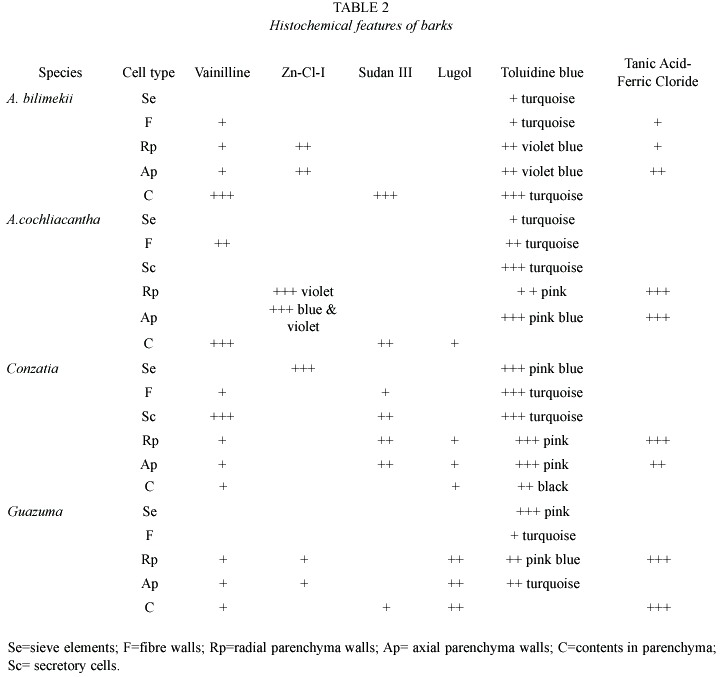
Histochemical studies. Toluidin-O blue was used to differentially stain cellulose (blue), hemicelluloses (purple,pink,and violet colors)and lignin (turquoise). Phenolic compounds were specifically stained by the vainillin technique. The Zinc-Iodine-Chlorine technique was used to differentially stain cellulose, lignin and some intracellular carbohydrates. Lipids were stained using Sudan III. Tannic acid-ferric chloride technique was used to stain calcium pectates. Starch grains were localized by Lugol staining. (Johansen 1940, Jensen 1962, Krishnamurty 1988, Quintanar et al. 1997). The intensity of the reactions was evaluated as previously described (Rivera et al.1999).
Enzyme cytochemistry. Catalase, phenoloxidase and succinate dehydrogenase were tested in sections of freshly cut material, according to Harris and Oparka (1994). Catalase was localized as black, brown or dark gray deposits. Phenoloxidase activity sites were localized in lumina and walls of axial and radial parenchyma as blue or black deposits. Succinate deshydrogenase activity sites could be found as brown or black deposits in cell lumina (Bancroft 1975, Datta and Kumar 1987).
Uses. Much of the information concerning the traditional uses of the species here studied was obtained by one of the authors (Solares) after extensive personal experience working in the communities. Additional information was recently collected by applying a questionnaire on the uses of these and other plants to at least ten selected persons from each site. Some information for Barranca Honda was taken from Boyás and Solares (1993b).
Results
Acacia bilimekii Humb.&Bonpl.
General features of the species
Common name:Tehuixtle.-This species can be found as shrubs or trees up to 6 m tall, with a stem diameter 15 to 40 cm. Isolated individuals, distributed about 200 trees/km2 in density. When the dry forest is disturbed its growth can be favoured, as it happened in Izúcar de Matamaros, state of Puebla (Rzedowskii 1978), and Tehuixtla, state of Morelos (Solares 1997a).
Gross features of bark:The bark is wrinkled with large, thick plates; non collapsed secundary phloem of 2.5 mm, collapsed secundary phloem of 0.8 mm and rhytidome of 5.25 mm.
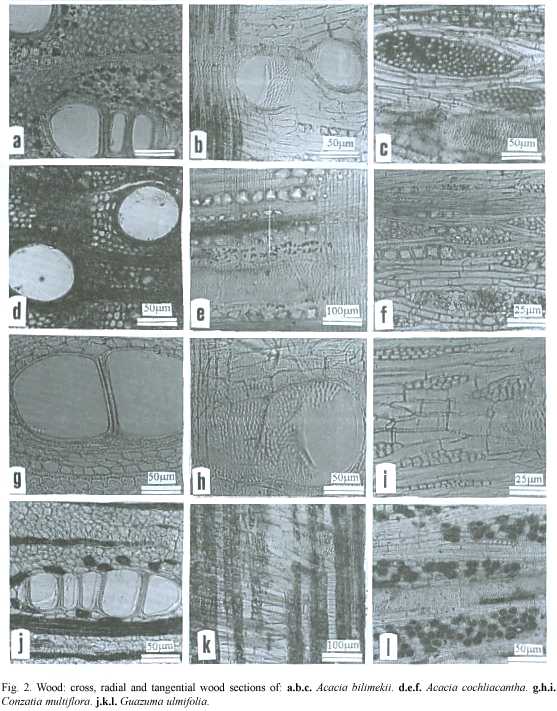
Histochemical and microscopic features of secondary phloem (Table 2,Figs.1a-c):

Sieve elements .These cells measure 208 (180-242) µm long by 38 to 45 µm wide; the cell wall width is 2 to 3 µm. Sieve plates are scalari-form and show low lignification of cell walls.
Axial parenchyma cells. This cells are arranged as short tangential bands that sligthly dilate tangentially at 700 µm from the vascular cambium. Sometimes the small bands, which are longitudinal strands interrupted by rays, get together in the dilated phloem rays.
Radial parenchyma cells . They are arranged in 3-5 series, reaching up to 10 series when they dilated. Rays are 91 µm (77-115) wide by 436 (306-536)µm heigth. At 1.9 mm from the vascular cambium ray dilatation is bigger, reaching more than 30 series. Rays are homogeneus formed by procumbent cells.
Axial and radial parenchyma cell walls showed low intensity when stained for polyphenols and medium intensity for pectins and cellulose (violet and blue reactions). Calcium pectates stain with medium intensity in axial parenchyma walls, but with low intensity in radial parenchyma walls. Ambar deposits in unstained axial and radial parenchyma lumina showed an intense reaction when polyphenols and lipids were stained.
Fibres. These cells are arranged in packages surrounded by crystal-containing axial parenchyma sheats. They are 1016 (879- 1201) µm long, 14 (10-20)µm wide, with a 6 (4-8) µm wall thickness. Primary fibre walls showed low intensity staining reactions for polyphenols, lignins and calcium pectates, whereas secondary walls showed no staining at all for the same compounds.
Periderm. This region is formed by suberized cells touching the inner dilated axial parenchyma. Two layers of periderms were seen. Isodiametric inner layer walls of outer periderm showed a medium intensity reaction for lipids and the orange color of rhytidom layer increased. These walls showed no reaction for lignin.
Enzimatic cytochemistry (Table 11): High activity of catalase was found in the lumina of axial and radial parenchyma cells. The cambial region showed phenoloxidase activity. No activity of succinate dehydrogenase.

Gross features of wood:This species does not present a clear heartwood region. Wood is yellow, 2.5Y 8/6 color, no odour nor taste, interlocked grain, medium texture and smooth figure.
Histochemical and microscopic features of secondary xylem (Table 3,Fig.2a-c): Vessel elements (pores) visible to the naked eye, the rest of the xylem cells can be seen with a magnifying glass.
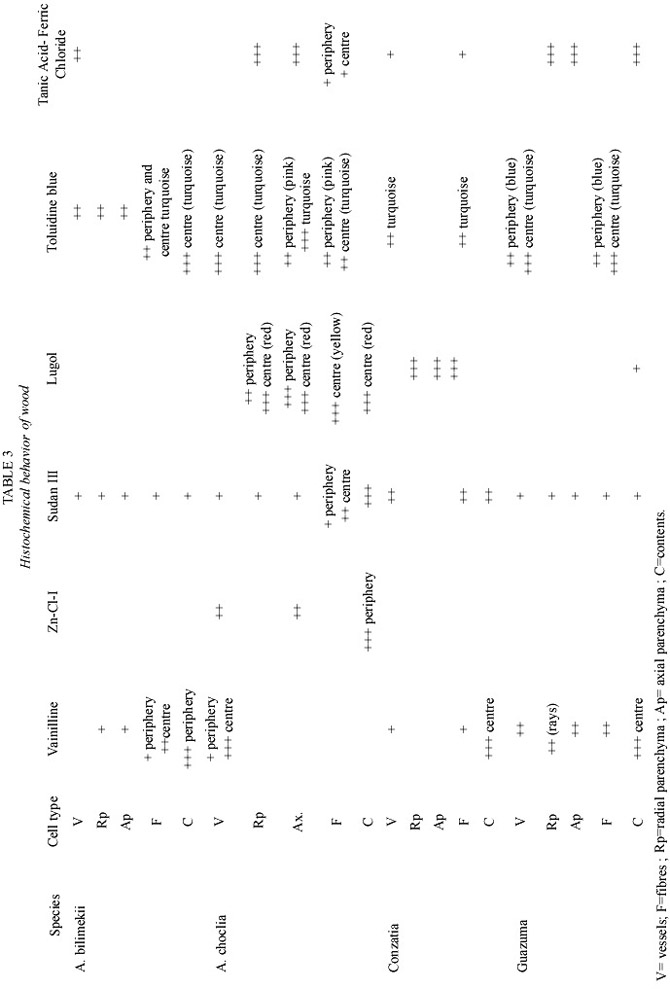
Vessel elements. Diffuse porous arrangement, mostly solitary and in radial multiple of 2 to 4 vessels, some in clusters of 3 vessels, moderately few. These cells are very short and medium vessel diameter; have alternate intervessel pits, simple perforation plates, vessel-ray pits similar to intervessel pits. Some vascular traqueids present.
Some ambar deposits in unstained vessel lumina showed a medium reaction when stained for calcium pectates and polyphenols. A low reaction for lipids was observed.
Axial parenchyma. Aliform and confluent containing many starch grains in the periphery that are substituted by black and turquoise polyphenol deposits toward the centre. Crystals present.
Radial parenchyma .Most rays are 4 (2- 5) series, homogeneus, moderately numerous, extremely low, medium width. Absence of starch in outer sapwood but black polyphenol deposits in the middle of xylem are present.
Parenchyma walls showed low reaction for polyphenols and lipids and medium reaction for lignin, which was very similar to reactions presented by vessel walls.
Fibres. Libriform, of medium length, fine diameter and thick walls. In outer sapwood low intensity staining reaction to polyphenols, lipids and medium reaction for lignins were observed.
Enzimatic cytochemystry (Table 11): Axial and radial parenchyma lumina showed medium activity of catalase. Medium and high activities of phenoloxidase and succinate dehydrogenase, respectively, were found in axial an radial parenchyma lumina towards vascular cambium.
Specific gravity: High (0.81).
Acacia cochliacantha Mcbride.
General features of the species
Common name: cubata. This species can be found as trees but mostly as shrubs,up to 5 m tall, having a stem diameter up to 20 cm. They are about 400 trees/km2 in density. It can grow in disturbed dry forests substituting other species, making almost pure forest formations known as cubateras.
Gross features of bark: The bark is smooth; non collapsed secundary phloem of 1.05 mm and collapsed one of 0.99 mm.
Histochemical and microscopic features of secondary phloem (Table 2,Fig.1d-f and 3d,k):
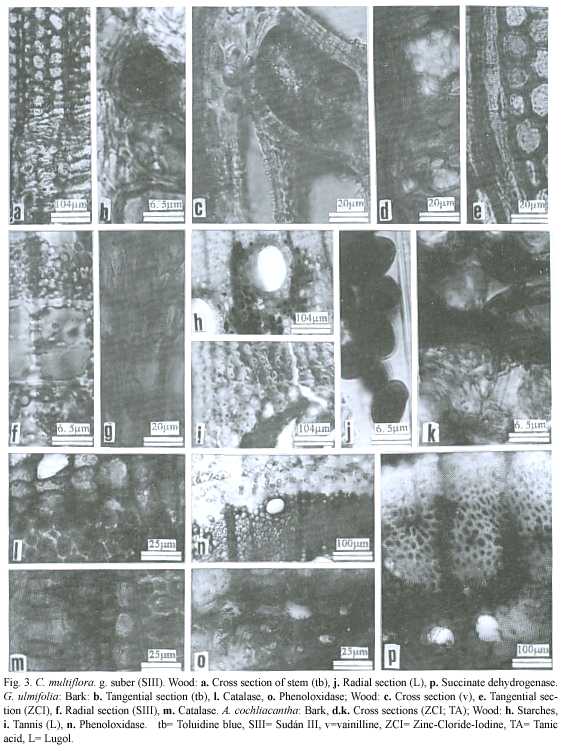
Sieve elements .These cells are 402 (300- 460)µm long, 30 to 50 µm wide, and 4 to 12 µm wall thickness. Sieve plates are scalari-form and show low lignification of cell walls.
Axial parenchyma .This cells are arranged in tangential bands which eventually are dilated form a concentric band as far as 1 mm from the vascular cambium. These bands, which are longitudinal strands interrupted by rays, get together in the dilated phloem rays. Some polyphenol deposits in lumina are found.
Radial parenchyma. They are arranged in 3-7 series and when they dilated reach more than 40 series. Rays are 54 (45-61) µm wide and 508 (382-842) µm height. At 800 µm far from the vascular cambium ray dilatation is bigger, reaching more than 40 series. Rays are homogeneus, formed by procumbent cells. Some deposits were lipids and other reacted intensely for poliphenols. Romboidal crystals were found. The radial and, even more, the axial parenchyma walls showed an intense reaction to cellulose and hemicellulose.
Fibres. These cells are arranged in packages surrounded by crystal-containing axial parenchyma sheats. They are 1281 (1162-1392) µm long, 14 (10-20) µm wide and 5 (4-8) µm in cell wall thickness. Primary fibre walls showed a medium reaction for lignins and polyphenol staining. It is remarkable the lack of reaction of secondary walls for the same compounds.
Sclereids. A concentric band of brachysclereids is present next to the axial parenchyma band. Its walls showed a high reaction for lignins.
Periderm. Highly suberized walls are arranged as the last concentric band of the stem.
Enzime cytochemistry (Table 11): High activity of catalase was found in axial and radial parenchyma lumina. Blue deposits of phenoloxidase were frequently present in axial parenchyma lumina of non collapsed phloem. Some activity of phenoloxidase was found in a meristematic region next to the collapsed phloem, and high activity of succinate dehydrogenase in the axial and radial parenchyma near to that meristematic region, this activity decreased toward the vascular cambium.
Gross features of wood:This species does not present a clear heartwood region. Wood is yellow, color (2.5Y 8/6),no characteristic odour nor taste, interlocked grain, medium texture and smooth figure.
Histochemical and microscopic features of secondary xylem (Table 3,Fig.2d-f,3h-i): Vessel elements (pores)and axial parenchyma visible to the naked eye, rays can be seen with a magnifying glass.
Vessel elements. Diffuse porous arrangement, mostly in radial multiple of 2 to 3 vessels, some solitary and in clusters of 3 vessels, moderately few. These cells are very short and medium vessel diameter, have alternate intervessel pits, simple perforation plates, vessel-ray pits similar to intervessel pits and vestured. Some vasicentric traqueids are present.
Vessel walls reacted low for polyphenols and medium for cellulose, particularly those of the outer sapwood. In the middle of the stem a high degree of lignification and deposition of polyphenols in lumina are observed. Black deposits of calcium pectates are present in vessels of outer sapwood and red tannin deposits in vessels of inner wood. The color of ambar deposits in unstained vessel lumina deepened when lipids were stained.
Axial parenchyma. Aliform and confluent, with romboidal crystals of two sizes. Starch is abundant in outer sapwood and red tannin deposits in inner wood. A medium reaction for cellulose and hemicellulose in axial parenchyma of sapwood and very high for calcium pectates were observed. The heartwood showed a high reaction for lignins.
Radial parenchyma. Rays are uniseriates or have 4 (2-5)series, homogeneus, moderately numerous, extremely low and medium width. Absence of starch in outer sapwood. Black polyphenol deposits in the middle of xylem. Parenchyma walls showed a high reaction for calcium pectates next to the cambial region, however, toward the middle, lignins reacted very high. Using lugol some deposits in sapowood reacted to carbohidrates, whereas in the middle tannins were observed.
Fibres. Are libriform, of moderately short length, fine diameter and thick walls. The reaction to lignin increased from the outer sapwood toward the center, whereas the reaction for cellulose and hemicellulose decreased. A low reaction for calcium pectates was shown by the outer sapwood fibres, by primary walls of center fibres and by the middle lamella. Lipids increased from outer sapwood towards the pith.
Enzimatic cytochemystry (Table 11,Fig. 3n): High activity of catalase and blue deposits of phenoloxidase were present in axial and radial parenchyma throughout all the stem. Black deposits of succinate dehydrogenase were present in the radial parenchyma throughout all the stem.
Specific gravity:High:0.75.
Conzattia multiflora (Rob)Stand.
General features of the species
Common name: parotilla, guayacán. This species is a tree up to 17 m tall, stem diameter from 20 to 70 cm. Is amply distributed in the tropical dry forests of Morelos State, its density is 4000 trees/km2 (Fig.2 and 3).
Gross features of bark:The bark is smooth with marked lenticels.Secondary phloem of 1.08 mm and peridem 0.95 mm wide.
Histochemical and microscopic features of secondary phloem (Table 2, Fig.1g-i, 3g,):
Sieve elements .Cells 402 (320-460) µm long, 32 to 55 µm wide,and 4 to 6 µm wall thickness. Sieve plates are scalariform, showing radial and tangetial distribution in sieve elements and a high staining reaction for cellulose and hemicellulose.
Axial parenchyma.This cells are surrounding the sieve elements and tend to form bands slightly dilated as far as 610 µm from the vascular cambium.These bands, which are longitudinal strands not interrupted by rays, have polyphenol deposits and many druses.
Radial parenchyma They are arranged uniseriate and biserate, when dilated they reach more than 20 series. Rays are 41 (30-46) µm wide and 235 (198-306) µm height. At 600 µm from vascular cambium, ray dilatation is bigger, reaching more than 20 series. Rays are homogeneus, formed by procumbent cells. A medium reaction was observed for polyphenols in ray lumina. Starch grains are present.
Axial and radial parenchyma walls reacted slightly to polyphenols and intensely to cellulose, hemicellulose and calcium pectates. A medium reaction for lipids was obseved.
Sclereids. A brachysclereids concentric band next to the ray dilated region is present. In outer layers this cells form little packages tangentially arranged. High reaction to lignin was observed.
Fibres. Very few cells, diffusely distributed, 793 (650-850) µm long, 19 (16-24) µm diameter, and 8 (6-10)µm cell wall thickness. Walls showed a high reaction for lignins and low for polyphenols.
Secretory cells .Cells in the non collapsed region have thin walls and big cell cavities full of tyloses and oils. Walls of these cells become thicker towards the collapsed phloem and lignin, oils and polyphenols increase.
Periderm. Formed by a concentric band of brachysclereids with very thick walls, a suberized parenchyma that includes brachysclereid packages and an outer epidermic layer.
Enzime cytochemystry (Table 11,Fig 3o): High activity of catalase and deshidrogenase succinate was shown in almost all the cell types except in brachysclereids. Axial parenchyma and vascular cambium showed medium activity of phenoloxidase, this activity increased toward the periphery, principally in the suberized and collenchyma cells.
Gross features of wood: This species does not show a clear heartwood region. Wood is yellow (2.5Y 8/6 color), no odour nor taste, interlocked grain, medium texture and smooth figure.
Histochemical and microscopic features of secondary xylem (Table 3,Fig.2g-i,3a,j): Vessel elements (pores) visible to the naked eye, rays and axial parenchyma can be seen with a magnifying glass.
Vessel elements. Diffuse porous arrangement, mostly solitary, but also in radial multiple of 2 to 6 vessels and some in clusters of 3 to 6 vessels, few. This cells are very short and moderately large diameter, have alternate intervessel pits, simple perforation plates and vessel-ray pits similar to intervessel pits. Some vascular traqueids containing gums are present.
Vessel walls slightly reacted to polyphenol staining and moderatly to lignins. Black deposits of calcium pectates and lipids are present.
Axial parenchyma. Scanty vasicentric paratracheal.
Radial parenchyma. Rays are uniseriate and biseriate, homogeneus, numerous, extremely low and very fine width.Toward the centre, walls showed medium reaction to lignin. Lipids are present.
Fibres. Are libriform, of medium length, medium diameter and very thin walls. Abundant starch grains, a moderate presence of lipids and calcium pectates were observed in the outer sapwood; toward the centre, the reaction to polyphenols was high and medium to lignins.
Enzime cytochemystry (Table 11,Fig.3o): Catalase activity was found in all cells near to vascular cambium, its activity decreased towards the pith but is still seen in radial parenchyma. Low activity of succinate dehydrogenase was present in parenchyma cells near to the vascular cambium. No activity of phenoloxidase was found.
Specific gravity: Medium (0.55).
Guazuma ulmifolia Lam.
General features of the species
Common name: cuahulote. This species can be found as trees, up to 9 m tall, 20 to 40 cm stem diameter. Due to an excessive rate of exploitation, today it reaches a low density of 300 trees/km2 and it has an isolated distribution.
Gross features of bark:The bark is wrinkled with thick large plates; non collapsed secundary phloem of 1.56 mm, collapsed secundary phloem of 0.15 mm and rhytidome up to 3 mm.
Histochemical and microscopic features of secondary phloem (Table 2,Fig.1j-l,3b):
Sieve elements .These cells are 265 (250- 286)µm long, 10 to 25 µm wide and 2 to 3 µm wall thickness. Sieve plates are scalariform and show high reaction to cellulose and hemicellulose. This cells have a tangential arrangement similar to the axial parenchyma.
Axial parenchyma cells. Arranged in regular short tangential bands.The small bands are uninterrupted longitudinal strands. At 2.8 mm from the vascular cambium,a marginal concentric band divides the non collapsed phloem from apparently annual regions of old non fuctional phloem which are 800 µm width in average.Crystals are present.
Radial parenchyma Arranged in 4 (3-5) series,reaching more than 10 series when dilated. Rays are 13 (12-15) µm wide and 62 (58-66) µm height. At 400 µm from the vascular cambium ray dilatation is bigger, reaching more than 10 series. Rays are homogeneus, formed by squared cells. Oval secretory canals along the rays are observed.
Axial and radial parenchyma walls showed a low reaction to polyphenols. Radial and axial parenchyma wall showed a medium reaction to cellulose and hemicellulose and high to calcium pectates, however, a medium reaction to lignin was observed in axial parenchyma walls surrounding fibres. Lipids, starch grains and polyphenols were detected in their lumina.
Fibres. These cells are arranged in packages surrounded by a crystal-containing axial parenchyma sheath. They are 704 (680-727) µm long, 16 (14-18) µm diameter and 7.5 (6-8) µm cell wall thickness. Walls showed a low reaction for lignins and none for polyphenols.
Periderm. Formed by suberized cells belonging to the dilated radial parenchyma band and the phelema.
Rhytidome is formed by four layers of suberized secondary phloem and its periderm.
Enzime cytochemistry (Table 11,3l, 3ñ): The activity of catalase was high in most cells, except in fibres and dilated rays. The activity of phenoloxidase was high in most cells, except fibres. The axial parenchyma and suberized cells showed a high activity of succinate dehydrogenase.
Gross features of wood: This species does not have a clear heartwood region. Wood is pale brown (10YR 8/3 color), no odour nor taste, straigth grain, medium texture and smooth figure.
Histochemical and microscopic features of secondary xylem (Table 3,Fig.2j-l,3c,e, f): Vessel elements (pores)and rays are visible to the naked eye; axial parenchyma can be seen with a magnifying glass.
Vessel elements. Diffuse porous arrangement, mostly in radial multiple of 2 to 7 vessels and some solitary, moderately numerous. These cells are very short and medium vessel diameter, have alternate intervessel pits, simple perforation plates, and vessel-ray pits similar to intervessel pits. Walls showed a moderate reaction to polyphenols. In the outer sapwood, vessels reacted moderately to cellulose (blue)and strongly for lignin in the centre. Polyphenols inccreased towards the centre. Vascular traqueids were present.
Axial parenchyma. Vasicentric and diffuse in aggregates.
Radial parenchyma. Rays are 4 (2-6)series, heterogeneus, formed by tile and procumbent cells, moderatly numerous, extremely low and medium width. Parenchyma walls showed a moderate staining for polyphenols and intense for calcium pectates. Some deposits in ray lumina were identified as starch grains and calcium pectates. A high amount of polyphenols was observed in procumbent ray cells.
Fibres. Libriform, of medium length, medium diameter and thin walls. Medium reaction for polyphenols in walls. The content of cellulose decreased toward the center whereas lignin increased.
The inner layer of the walls showed a high reaction to cellulose and hemicellulose, this reaction increased in the medium lamella next to thiner walls.
Enzime cytochemystry (Table 11,3m): Activity of catalase was moderate in the inner walls of fibres and vessels and very high in radial parenchyma walls. High activity of phenoloxidase was observed in parenchyma. Succinate dehydrogenase activity was low in vessels but high in radial parenchyma.
Specific gravity: Medium (0.58).
Traditional uses of the studied species
Table 12 records the uses given to the four species in the three sites of collection. Most rural communities of Morelos use their forest resources in similar ways, however, some differences among particular places are due to their own local customs. Forest products are obtained all year-round, but more intensively during the dry season (November-April), when most people are not working in agriculture. The two Acacia species and G.ulmifolia are heavily used in the three places, where several different uses were recorded during the fieldwork: fire- wood, dead and live fences, fodder, construction, "tutors " (supporting stakes for cultivated plants), handcrafts, farming tools, extraction of tanning products, and medicine. The fruit of cuahulote, G.ulmifolia , is used in combination with the fruits of Basacantha echinocarpa (granjel)and Crescentia alata (cuatecomate) as medicine for treating kidney diseases: pulp of the three fruits is boiled in water and the resulting infusion is drunk during at least ten days.

It is notewhorthy that there is no relationship beteween use and abundance. An example is shown by G.ulmifolia (cuahulote), the species most heavily used in these three subzones, with a density of 300 trees/km2 , whereas C. multifolia (parotilla), the most abundant (4000 trees/km2 ), is only moderatly used as fodder and for making beams. During the long dry periods goats feed on the bark of A.cochliacantha that, according to our results, is rich in carbohydrates; the regeneration capacity of the bark is limited and overfeeding has inflicted an irreversible damage on this species (Solares 1995,Orduño 1998).
A.cochliacantha and A.bilimekii are also used as fuel and fodder. G.ulmifolia is used for medical purposes and furniture making and C.multifolia only as fuel (Nolazco and Sánchez 1991).
Discussion
Secondary Phloem
Many drought-deciduous species of tropical dry forests have smooth and pale bark; this, together with common trends in their cell structure and organization, could be related to their survival in the extreme conditions prevailing in such environments. The structure, composition and function of the secondary phloem is involved in mechanical and biological protection and in the nutrition of the secondary stem.
Common structural trends in the bark of the four species studied are: axial parenchyma bands interrupted by rays that dilate at different distances from the vascular cambium and end in a tangential concentric band. This arrangement has been also found in Cercidium plurifoliolatum Micheli Mich. and Lysiloma divaricata (Jacq.) Macdr. (Paredes et al.2001), which are also drought-deciduous species of tropical dry forests. The arrangement of axial parenchyma, rays, fibres and periderm follow the pattern proposed by Furuno (1990), in which small axial parenchyma bands alternate with fibre bands or sieve elements, except in C.multiflora in which fibres are surrounded by a crystal-containing axial parenchyma sheath. This arrangement appears as a reticle-like pattern and is related to mechanical properties like: high adhesion (difficulty in getting loose), low radial shrinkage due to ray size and arrangement. Well lignified fibres and sclereids, as in C.multiflora, contribute hardness to the bark. The abundance of different types of crystals provides mechanical resistence to infection. These anatomical properties and the abundance of extractives could be a strategy to contend with very high temperatures during the dry season and a great diversity of pathogens. Aditionally, the presence of tyloses in laticifer canals in Conzatia could be a mechanism to constrain the dispersal of pathogens.
The extractives present in the bark of these tropical dry forest trees are mainly of phenolic origin and are more abundant than tannins. The presence of lipids and starch grains in parenchyma cells, together with catalase and phenoloxidase activities, suggest a high level of metabolic activity, probably associated to the synthesis of extractives and to the process of lignification (Thomas et al.1995).
Secondary xylem
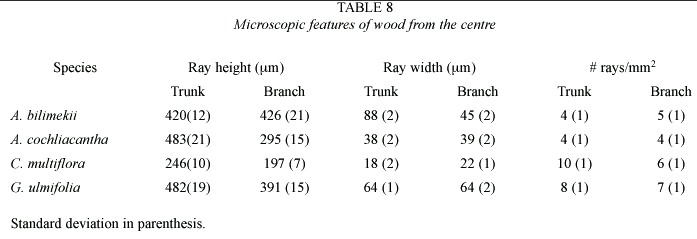
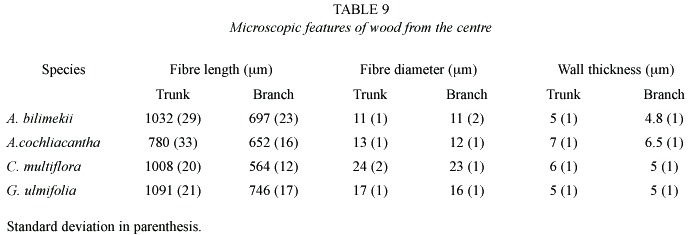
Common structural trends in the wood of the four species studied are: the number of vessels/mm2 in branches increases as compared to that in the trunk, whereas vessel diameter and length decrease. In the trunk, rays are higher, wider and more numerous; fibres are longer and their walls thicker. Wood diffuse porous, vessels in radial multiple, solitary and clustered, were common. Vessels have simple perforation plates, areolate-alternate intervessel pits and vessel-ray pitting similar to intervessel pits. Rays are wider, higher and more per mm in the outer than in the inner xylem. The number of vessels/mm2 increased towards the centre but vessel diameter and length increased towards the periphery. Fibre length diameter increased towards the outer layers althougth walls are thicker towards the centre.
Axial parenchyma in Conzatia was scanty vasicentric, but in the rest of the species was aliform, aliform confluent, and in bands. Multiseriate rays were present in Acacia species and G.ulmifolia but uniseriate and biseriate in Conzatia .
The four species here studied show high vulnerability indexes (Table 10), this should indicate a low resistance to cavitation, in other words they should not stand high water tension in vessels. Beureria cumanensis , a drought-deciduous species from Venezuela, has a vulnerability index of 0.68 (Sobrado 1997), more than one order of magnitude smaller than our four species. This apparent inconsistency obliges to study their physiological vulnerability in the future. The state of Morelos has abundant subsoil-water in layers not too deep, which could be reached by the roots of trees and could help to explain the high vulnerability indexes found in our four species. The case of Conzatia is remarkable since its index is more than two orders of magnitude higher than B.cumanensis, and is the most important species in the structure of the Morelos dry forest.
Histochemical results showed an increase of lignification of parenchyma and fibres towards the centre of the stem; the lipids and carbohydrates were progresively substituted by tannins and polyphenols. Whereas A. cochliacantha and Conzatia showed abundant lipid and starch deposits, calcium pectates were prevalent in A.bilimekii and Guazuma . In spite of the progressive lignification and polyphenol deposition there is no difference in color between sapwood and heartwood in the four species.
Our results showed that vascular region had the greatest enzimatic activity. Catalase was found active mainly in axial and radial parenchyma and in the Guazuma vessels, this could be related with the presence of lipids. Phenoloxidase activity was found in the parenchyma cells in three of the species. The occurrence of this enzime is associated to the lignification process, this could explain its presence in xylem, however Conzatia is an exception as it showed intense reactions to cellulose and calcium pectates instead. Succinate-dehydrogenase was very active in parenchyma cells but in A.cochliacantha this activity was localized throughout the xylem. The presence of these enzymatic activities indicate a high metabolic rate and in particular can be related to the metabolism of lipids that, in these tissues, serve as precursors for the synthesis of extractives. These results suggest that the formation and maturation of woody tissue is very active at the beginning of the rainy season.
The arrangement of axial and radial parenchyma could be related to an efficent water storage strategy and low tangential shrinkage values, that guarantee the survival during long periods of drough avoiding functional and mechanical dysfunction. To solve some of these questions the vulnerability curves and root systems of the four species should be studied, particularly in Conzatia, which has fibres of very wide diameter, containing high amounts of starch grains that could compensate a dysfunctional axial parenchyma. Additionally, the high enzymatic activities present in parenchyma cells show that they are very active in metabolic processes. The abundance of these cells in the species studied, in contrast with some temperate species such as Alnus spp, Carpinus caroliniana, Arbutus spp, etc.(Quintanar and De la Paz 1996) could be related to the multiple roles that they seem to play.
In spite of the scant uses reported in the literature for these species,our field data revealed that they are widely used in the three communities under study. The present common uses given to these species are based on tradition and practical considerations and, sometimes, do not correspond to the particular stem properties of the species. The anatomical and histochemical properties here studied suggest that each species could be rationally employed for some specific uses. The Acacia species should be of slow growth since their specific gravity values correspond to a highly lignified xylem, this explains its use as fences and farming tools. Conzatia, in contrast, should grow faster and have a lower decay resistance than the Acacia species, since it has the thinnest fibre walls and a lower content of extractives. The anatomical and histochemical properties of Guazuma are intermetiate between Conzatia and the Acacia species. The Acacia species should be employed for making tools, flooring, furniture, live or dead fences and other uses that require wood with high resistance to impact and friction and higher decay resistance. Guazuma could be employed in carving, turnering and for the manufacture of handcrafts or industrial products like pencils and veneers. Conzatia, on the other hand, seems to be not adequate for industrial or technological uses, but must be conserved as an important element in the dry forest ecosystem and has a potential use as an ornamental tree (Solares 1997a).
The stem anatomy and histochemistry reveal that there are cell patterns in bark and xylem, as well as deposition of storage substances, linked to strategies of survival to extreme dry conditions. Dry conditions should be related to a decrease of diameter and an increase of the number of vessels, but perhaps subsoil water accessibility and deciduousity are important factors that explain the vessel dimension found in this work, particularily in Conzatia. In addition, the abundance of storage cells can guarantee water supply in the dry season.
Histochemistry and enzime citochemistry suggest that the metabolism of storage lipids, providing precursors for lignification and the synthesis of extractives, is already active at the beginning of the rainy season.
Our field data reveal that the species here studied are widely used in the three communities under study. Alternative uses for these species are proposed based on their specific gravity as a density index.
Resumen
Se estudió la anatomía e histoquímica del tallo secundario de Acacia bilimekii, Acacia cochliacantha, Conzatia multiflora y Guazuma ulmifolia. Las muestras de tallo se colectaron en una selva baja caducifolia del estado de Morelos, México. La actividad enzimática se estudió en cortes frescos de caras tangenciales, radiales y transversales. La anatomía e histoquímica se hizo en cortes similares de muestras previamente ablandadas con una mezcla de agua-glicerol-PEG. Los resultados muestran que el patrón anatómico de la corteza y madera, así como las características histoquímicas no enzimáticas están relacionados con el acceso al agua y el clima; estos patrones garantizan que las estrategias mecánicas de resistencia al deterioro les permitan sobrevivir a condiciones extremas. Los resultados de la histoquímica y la citoquímica enzimática sugieren que la lignificación y la síntesis de extractivos a partir de los lípidos de reserva se encuentra activa desde el principio de la estación de lluvias. Se sugieren usos potenciales para las especies estudiads de acuerdo con las densidades relativas.
Palabras clave: Anatomía, corteza, madera, histoquímica, citoquímica, enzima, usos tradicionales, especies caducifolias, sequía, bosque seco, México.
References
Anonimous. 1975. Inventario Forestal del estado de Morelos. México. Dirección General del Inventario Forestal. SARH. Publicación No.32.41 p [ Links ]
Anonimous. 1989. IAWA list of microscopic features for hardwood identification. IAWA, Bull. n.s. 10(3): 219-332. [ Links ]
Anonimous. 1991. XI Censo General de Población y Vivienda; VII Censo Agropecuario Morelos. Instituto Nacional de Estadística, Geografía e Informática (INEGI). Aguascalientes, Aguascalientes, México.84 p. [ Links ]
Anonimous. 1994. Inventario Nacional Forestal. Subsecretaría Forestal y de la Fauna. SARH. Coyoacán. México D.F.100 p. [ Links ]
Anonimous. 1995. Perfil Sociodemográfico; Estados Unidos Mexicanos. Instituto Nacional de Estadística, Geografía e Informática (INEGI).Aguascalientes, Ags. México. 140 p. [ Links ]
Bancroft,J.D.1975.Histochemical techniques. Butterworths.London.347 p. [ Links ]
Boyás, D., J. C. 1991. Regionalización Ecológica del estado de Morelos, p.17-25. In Medardo T.U.(editor). Primeras Jornadas de Investigación en el estado de Morelos. UNAM. México. [ Links ]
Boyás, D., J. C., J. Javelly G., M. Cervantes S., F. Solares A., M. Linares A., L. Sandoval M., R. Soto & I. Naufal T. 1993a. Diagnóstico Forestal del estado de Morelos. INIFAP-SAGAR. Campo Experimental Zacatepec. Publicación Especial 7. Zacatepec, Morelos, México. 240 p. [ Links ]
Boyás, J. C. and Solares, A. F. 1993b. IV Guía de la excursión botánica a las Lagunas de Zempoala y ejido Barranca Honda en el estado de Morelos. In: Memoria del XXX Aniversario del Herbario Nacional Forestal y de la VII Reunión Nacional de Encargados de Herbario. Secretaría de Agricultura y Recursos Hidráulicos. Instituto Nacional de Investigaciones Forestales y Agropecuarias (INIFAP).Publicación Especial No.62. 2ª Ed. Coyoacán. D. F. P. 141-154. [ Links ]
Chattaway, M. 1932. Proposed standards for numerical values used in describing woods. Trop. Woods 29: 20-28. [ Links ]
Datta, S. K. & A. Kumar. 1987. Histochemical studies of the transition from sapwood to heartwood in Tectona grandis. IAWA Bull. n. s. 8:363-368. [ Links ]
Esau, K. 1965. Plant Anatomy. Wiley, New York. 767 p. [ Links ] Fahn, A. 1990. Plant Anatomy. Pergamon, England. 587 p. [ Links ]
Flores M., G., J. Jiménez, X. Madrigal S., F. Moncayo & F. Takaki 1971. Tipos de vegetación de la República Mexicana. Dirección de Agrología. SRH. México, D. F. 57 p. [ Links ]
Furuno, T. 1990. Bark structure of deciduous broad-leaved trees grown in the San in Region, Japan. IAWA Bull. n.s.11 (3):239-254. [ Links ]
Harris, N. & K. J Oparka. 1994. Plant cell biology: a practical approach. Oxford University. Great Britain. 329 p. [ Links ]
Jensen, W. A. 1962. Botanical histochemistry. W. H. Freeman, San Francisco. 408 p. [ Links ]
Johansen, D. A. 1940. Plant microtechnique. McGraw Hill Book Co., New York. 523 p. [ Links ]
Junikka, L. 1994. Survey of english macroscopic bark terminology. IAWA J. 15(1):3-45. [ Links ]
Krishnamurty, K. V. 1988. Methods in plant histochemistry. Bharathidasan University. S. Viswanathan Press. India. 89 p. [ Links ]
Kribs, D. 1968. Commercial foreing woods on the American market. Dover Publications, New York. 241 p. [ Links ]
Miranda, F. & E. Hernández X. 1963. Los tipos de vegetación de México y su clasificación. Bol. Soc. Bot. Méx. 28:29-179. [ Links ]
Nolazco, E .& A. Sánchez V. 1991. Guía para el reconocimiento de los principales árboles del alto balsas. División de Ciencias Forestales. Universidad Autónoma de Chapingo. Chapingo, México.207 p. [ Links ]
Nunes, E., T. Quilhó & E. Pereira. 1996. Anatomy and chemical composition of Pinus pinaster. IAWA J. 17(2):141-149. [ Links ]
Orduño, C. A. 1998. Anatomía de la corteza de cuatro especies de la selva baja caducifolia del estado de Morelos: origen, desarrollo y regeneración. Tesis de Maestría. Colegio de Postgraduados. Montecillos, México. 88 p. [ Links ]
Paredes, L. C., A. Quintanar, S. Acosta, G. Orozco, C. de la Paz Pérez-Olvera & A. Torre-Blanco. 2001. Histoquímica de la corteza de cinco angiospermas de una selva baja de Oaxaca. Madera y Bosques 7(2):49-61. [ Links ]
Quintanar, A. & C. De la Paz Pérez O.1996.Anatomía de la madera de ocho especies de clima templado. Bol. Soc. Botánica de México 58:5-14. [ Links ]
Quintanar, A, L. Rivera, A. Torre-Blanco & S. Rebollar. 1997. Comparative histochemistry and cell morphology of sapwood and heartwood of Gliricidia sepium (Fabaceae). Rev. Biol. Trop. 45:1005-1011. [ Links ]
Rivera, L., A. Quintanar & C. De la Paz Pérez.1999. Comparación histoquímica de albura y duramen de tres especies de Quercus .Madera y Bosques 5(1): 27-41. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa. México.432 p. [ Links ]
Sobrado, M. A. 1997. Embolism vulnerability in drought-deciduous and evergreen species of a tropical dry forest. Acta OEcol. 18(4):383-391. [ Links ]
Solares, A., F. 1995. Capacidad de regeneración de la corteza y evaluación fitoquímica antes y después del descortezamiento en cuachalalate. Tesis de Maestría. Colegio de Postgraduados. Montecillos, México. 100 p. [ Links ]
Solares A., F. 1997a. Determinación de la calidad de arbolado de 10 especies de la selva baja caducifolia en dos unidades ecológicas del estado de Morelos. INIFAP- SAGAR. Campo Experimental Zacatepec. Publicación Especial 7. Zacatepec, Morelos. México. 27 p. [ Links ]
Solares A., F. 1997b. Manejo, Conservación y Uso Integral del Cirián (Crescentia alata .H.B.K.) en el estado de Morelos. INIFAP-SAGAR. Campo Experimental Zacatepec. Publicación Especial 13. Zacatepec, Morelos, México. 25 p. [ Links ]
Solares A., F. 1999. Estudio químico-biológico para la producción sostenida de corteza para uso farmacéutico-industrial en el estado de Morelos, p. 20-28. In Informe Técnico. Alternativas de Conservación y Manejo de Especies Forestales del Trópico Seco. INIFAP-SAGAR. México. [ Links ]
Thomas, V., D. Premakumari, C. P. Reghu, A. O. N Panikkar & C. K. Sarawathy Amma. 1995. Anatomical and histochemical aspects of bark regeneration in Hevea brasiliensis. Annals of Botany.75:421-426. [ Links ]
Trockenbrodt, M. 1990. Survey and discussion of the terminology used in bark anatomy. IAWA Bull. ns. 11: 141-166. [ Links ]

