










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 suppl.2 San José Dec. 2004
Costa Rica durante El Niño (1997 - 1998)
Marco A. Quesada-Alpízar1 & Alvaro Morales-Ramírez2
1 Programa Regional de Posgrado en Biología, Sistema de Estudios de Posgrado, Escuela de Biología, Universidad de Costa Rica. 2060, San José, Costa Rica.
2 Centro de Investigación en Ciencias del Mar y Limnología (CIMAR) y Escuela de Biología, Universidad de Costa Rica, 2060 San José, Costa Rica; amorales@cariari.ucr.ac.cr
Recibido 13-XI-2003. Corregido 17-III-2004. Aceptado 21-V-2004.
Abstract
In order to understand the vertical distribution of planktonic organisms in the oceans, it is important to consider a number of factors. Including oceanic circulation patterns, the vertical structure of the water column and the vertical position of the thermocline. This study examines the hydrography of Golfo Dulce, a tropical fjord -like embayment in the Eastern Tropical Pacific, and the influence of the 1997-1998 ENSO on the system. Five sampling stations were selected and visited once a month for two years in order to obtain vertical profiles of temperature (ºC), salinity () and dissolved oxygen (mg O2/l). Results show a strong vertical stratification of the water column associated to the morphology of Golfo Dulces basin and to the effects of ENSO. Moreover, the existence of significant differences between the hydrographic characteristics of the inner and outer parts of the basin was detected. Finally, biannual events in the influx of cold, high salinity, and oxygen rich oceanic water masses into the basin were detected. These events might be related to the same forces driving the coastal upwelling system in the Gulf of Panama.
Key words: Golfo Dulce, anoxic basin, water masses, upwelling, El Niño, Eastern Tropical Pacific, Costa Rica.
La termoclina es una capa de agua donde es posible observar un cambio rápido de la temperatura en la dimensión vertical (Philander 1996, Thurman 1996). Se da con frecuencia en los océanos, especialmente en zonas tropicales y subtropicales, y juega un papel fundamental en la distribución de los organismos marinos. Al respecto, Longhurst (1985) enfatiza sobre la importancia que tiene una estructura física estable en la termoclina para mantener la gran diversidad de especies de zooplancton que se observa en esta parte de la columna de agua, sobre todo en zonas tropicales, donde se observan termoclinas estables y permanentes.
La región del Pacífico Oriental Tropical (POT) se distingue por presentar una alta producción biológica asociada a la presencia de zonas de afloramiento ecuatorial. Estas zonas muestran una significativa variabilidad intra e interanual. La primera es causada por cambios en la intensidad de los vientos, lo cual resulta en máximos de clorofila y biomasa durante el invierno en el hemisferio Sur. La variación interanual, por su parte, se relaciona con el fenómeno El Niño Oscilación Sureña (ENOS) (White et al. 1995), el cual consiste de un calentamiento anómalo de las corrientes marinas limítrofes orientales del Pacífico inducidas, a su vez, por anomalías en el campo de vientos del Pacífico Oriental. Como resultado del acople océano-atmósfera durante el ENOS, se observan efectos físicos a gran escala cerca de las
costas como, por ejemplo, el calentamiento de la capa de mezcla y el hundimiento de la termoclina y la nutriclina, entre otros (Dietrich 1963, Philander 1996, Thurman 1996, NOAA 2000). Como es de esperar, el ENOS tiene efectos significativos tanto sobre los procesos físicos como biológicos del POT.
El Golfo Dulce, en el Pacífico de Costa Rica, presenta características morfológicas que tienden a restringir la circulación del agua en su cuenca interna y a favorecer la formación de aguas profundas anóxicas. Por esta razón, su circulación se asemeja la de fiordos de latitudes altas (Richards et al. 1971). El término fiordo es aplicado a un estuario o a una cuenca profunda rodeada por terrenos de pendiente fuerte y en muchos casos, un umbral en su desembocadura (Rattray 1967). Lo anterior es de gran relevancia debido a que solamente existen cuatro cuencas tropicales con tales características: el Golfo Dulce, el Golfo de Cariaco (Venezuela), la bahía Darwin (Ecuador) y la bahía Kaoe (Indonesia) (Richards 1965). El presente trabajo describe los gradientes físicos presentes en la columna de agua del Golfo Dulce así como las posibles variaciones espaciales y temporales que estos puedan presentar a lo largo de un periodo de dos años, el cual coincidió con el fenómeno ENOS 1997-1998.
Material y métodos
Sitio de Estudio: El Golfo Dulce se ubica en el Pacífico sur de Costa Rica ( 8º 27 - 8º 45 N ; 83º 07 - 83º 30 W) y tiene cerca de 50 km de longitud y entre 10 y 15 km de ancho, con un área superficial de aproximadamente 680 km2 . Se caracteriza por poseer una cuenca interna de pendiente fuerte con una profundidad máxima de 215 m y una cuenca externa poco profunda de aproximadamente 70 m (Richards et al. 1971, Hartmann 1994, Thamdrup et al. 1996, Umaña 1998). La cuenca recibe de 4000 mm a 5000 mm de lluvia por año y la estación lluviosa se extiende desde abril hasta diciembre. La precipitación por lo general excede a la evaporación durante al menos ocho meses al año. Las máximas temperaturas atmosféricas se registran durante la estación seca, época durante la cuál también se da un incremento en el número de horas diarias de brillo solar (Hartmann 1994, Wolff et al. 1996).
Hidrografía: Las características de fiordo del Golfo Dulce se reflejan en la composición de sus masas de agua. Dos capas son evidentes: una capa superficial cálida (19-30 °C) y diluida (salinidad < 34), que alcanza los 50–60 m de profundidad, justo sobre la profundidad del umbral ubicado en la parte externa del Golfo. La otra capa la componen las aguas profundas y físicamente homogéneas, con un mínimo de temperatura de 15.4 °C y salinidades que alcanzan los 34.8. La mezcla y circulación de estas aguas es influenciada por las fuerzas de mareas, el viento, el ingreso de agua al sistema, el afloramiento de agua subsuperficial y la topografía de la cuenca (Richards et al. 1971, Córdoba & Vargas 1996). Recientemente, Svendsen et al. (2004) reportaron para el Golfo una estructura de corrientes de tres capas en la parte externa y una circulación de tipo estuarino en la parte interna.
La renovación de las aguas profundas en el Golfo Dulce es lenta y aparentemente restringida a dos periodos de entrada de agua subsuperficial (Hartmann 1994). Se ha identificado masas densas de agua que pasan sobre el umbral externo y entran a la cuenca, donde se hunden rápidamente y desplazan las aguas profundas hacia arriba (Richards et al. 1971).
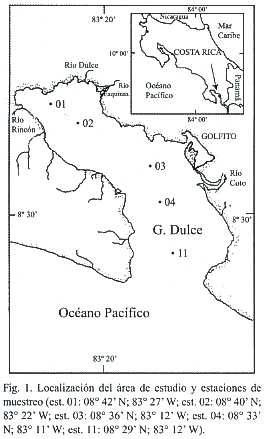
Muestreo: Se seleccionó un total de cinco estaciones de muestreo distribuidas en las partes interna (estaciones 01 y 02), media (estación 03) y externa (estaciones 04 y 11) del Golfo Dulce (Fig. 1). Mientras la profundidad de las estaciones 01, 02 y 03 supera los 150 m, la de las estaciones externas (04 y 11) no es mayor de 70 m. El programa de muestreo comprendió un total de 17 visitas al área, realizadas una vez al mes a lo largo de un periodo de dos años consecutivos, entre abril de 1997 y noviembre de 1998. Dicho programa incluyó muestreos tanto durante la época seca como la lluviosa. En cada fecha y estación se colectó una serie de muestras de agua usando una botella Niskin de 2 l a las siguientes profundidades: 0, 10, 20, 30, 40, 50, 60, 70 y 100 m. Además, a partir de 1998 se tomó una muestra adicional de agua a 150 m de profundidad en las estaciones 01, 02 y 03. Para cada una de las muestras, se determinó la salinidad (), con un refractómetro óptico, así como la temperatura (°C) y oxígeno disuelto (mg O2 /l), con un medidor de oxígeno YSI 52. A partir de estos datos, se distinguió una capa superficial (P1), ubicada por encima de la termoclina, una capa intermedia (P2), ubicada a la profundidad de la termoclina y una capa profunda (P3) por debajo de la termoclina. Además, en cada estación se midió la transparencia del agua usando el disco de Sechii. Se aplicaron pruebas de estadística descriptiva (promedio, desviación estándar, ámbito) y pruebas de ANDEVA (prueba post-hoc: Tukey) con un nivel de confianza de 95% con el fin de comparar las variables de temperatura, salinidad y oxígeno disuelto para cada estación y cada capa de agua muestreada (P1, P2, P3).
Resultados
Temperatura: Se observó un fuerte gradiente vertical de temperatura en todas las estaciones, dándose diferencias significativas en la temperatura promedio de cada capa muestreada (P1, P2 y P3) (F=423.23, p<0.01) a lo largo del periodo de estudio. En general, las capas superficiales (P1) de todas las estaciones se caracterizaron por tener promedios anuales de temperatura que varían entre 29.2 y 29.7 °C. Para el caso de las estaciones de mayor profundidad (01, 02 y 03), las capas intermedia (P2) y profunda (P3) tuvieron temperaturas promedio de 24.6-25.7 °C y de 20.4-21°C, respectivamente. Mientras tanto, en las estaciones de menor profundidad (04 y 11), los promedios de temperatura en las capas P2 y P3 variaron entre 26.9-27.3 °C y 23.4-23.7 °C, respectivamente. El caso de las estaciones 04 y 11 se considera por separado debido a que la menor profundidad de su columna de agua tiene influencia sobre las temperaturas mínimas esperadas para las capas intermedia y profunda. Además, al comparar los valores de temperatura por capa entre las estaciones, se determinó que existen diferencias significativas (F=3.71, p<0.05) entre éstas. Sin embargo, dichas diferencias se dan entre las estaciones internas (01, 02, 03) y las externas (04, 11) (Tukey, p<0.01) pero no dentro de cada grupo (Tukey, p>0.05).
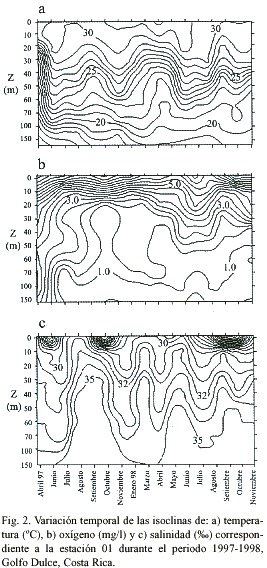
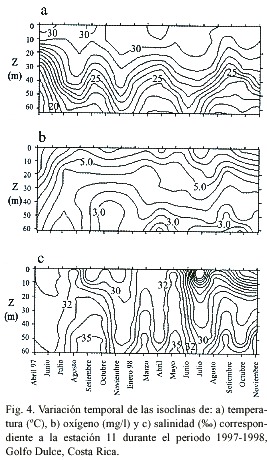
La temperatura de la columna de agua registró los valores más bajos en abril de 1997 en todas las estaciones. Dichos valores no se volvieron a presentar durante el periodo de estudio. Entre abril y junio de 1997 se observó un incremento en la temperatura de la columna de agua en las partes interna (estación 01) (Fig. 2a), media (estación 03) (Fig. 3a) y externa del Golfo Dulce (estación 11) (Fig. 4a). Dichas temperaturas persistieron durante todo el periodo de estudio. Además, se observaron eventos periódicos de enfriamiento de la columna de agua durante los meses de marzo-abril y agosto-septiembre de cada año. Estos eventos se reflejan en la elevación de las isotermas durante el periodo citado, para su posterior descenso en los meses siguientes 4a). Dicho patrón se repitió en cada una de las estaciones muestreadas y parece ser producto de periodos de entrada de agua oceánica (subsuperficial) a la cuenca del Golfo Dulce.
Oxígeno Disuelto: al igual que para el caso de la temperatura, se observó un fuerte gradiente vertical en los valores de oxígeno disuelto, reflejado en las diferencias observadas (F=442.6, p<0.01) entre las distintas capas muestreadas. Al observar los valores correspondientes a la concentración de oxígeno disuelto por capa, la P1 en todas las estaciones se caracterizó por tener promedios anuales de entre 4.4 y 5 mg O2 /l. En el caso de las estaciones 01, 02 y 03, las capas P2 y P3 mostraron promedios anuales en la concentración de O2 de 1.9-2.6 mg/l y 1.1-1.3 mg/l respectivamente. En las estaciones 04 y 11 los promedios anuales de las capas P2 y P3 variaron entre 3.3-3.5 mg O2 /l y 2.1-2.3 mg O2 /l, respectivamente.
Al igual que para el caso de la temperatura, los valores de oxígeno disuelto del grupo estaciones 01, 02 y 03 se diferencian de los de las estaciones 04 y 11 (Tukey, p<0.01) pero no existen diferencias entre las estaciones de cada grupo (Tukey, p>0.05). Sin embargo, se observaron patrones similares en el comportamiento temporal de las isoclinas de oxígeno disuelto en la parte interna (estación 01)(Fig. 2b) y parte externa del Golfo Dulce (estación 11)(Fig. 4b). Al observar la isoclina correspondiente a 1 mgO2 /l, por ejemplo, se aprecia como se da un incremento en los valores de oxígeno disuelto durante los mismos periodos en los que se reporta el enfriamiento de la columna de agua, lo cual sugiere la entrada de
agua relativamente rica en oxígeno. Lo anterior resulta más claro para el caso de la parte interna del golfo (Fig. 2b), la cual es la zona de mayor profundidad. En estas estaciones, resalta el hecho de que se dan concentraciones relativamente altas de oxígeno (hasta 1 mgO2 /l) a profundidades mayores a los 100 m, especialmente en los periodos en los que se observa el enfriamiento en la columna de agua.
En el caso de las estaciones externas (Fig. 4b), el comportamiento de las isolíneas de oxígeno disuelto no resultó tan claro. En ambos casos, durante el mes de abril (1997) la columna de agua presentó niveles de oxígeno relativamente altos hasta el fondo.
Salinidad: Los valores de salinidad promedio por año para cada capa no presentan fuertes fluctuaciones y sus valores se mantuvieron dentro de los siguientes rangos para todas las estaciones: 28.9-30.5 en la P1; 32.7-33.7 en la P2 y 34.1-34.9 para la P3. No se observaron diferencias significativas en la salinidad de las distintas estaciones (F=1.07, p>0.05) pero si en la salinidad de las distintas capas muestreadas (F=105.53, p<0.01). De este modo, los valores de salinidad en todas las estaciones y capas muestreadas se comportaron de manera similar, con pequeños incrementos en agosto-setiembre (1997) y marzo y mayo de 1998. Además, en todas las estaciones se registraron valores de salinidad relativamente bajos en abril de 1998, lo que coincide con el inicio de la estación lluviosa en el Pacífico sur en este año.
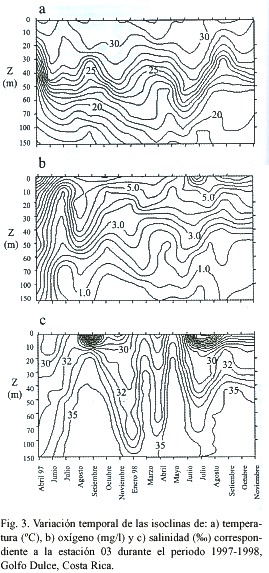
Las isohalinas para las estaciones internas (estación 01) (Fig. 2c), media (Fig. 3c) y externas (estación 11) (Fig. 4c) reflejan aumentos en la salinidad en aguas profundas durante los meses de septiembre (1997-98) y marzo y mayo de 1998, como es de esperar en caso de que se presenten entradas de masas de agua oceánica. Así mismo, las bajas salinidades que se observan en las aguas superficiales durante algunos meses coinciden con periodos de alta precipitación que se dieron días y/u horas antes de los muestreos.
Discusión
Como se esperaba, se comprobó la existencia de fuertes gradientes verticales en las condiciones de temperatura, oxígeno disuelto y salinidad en el Golfo Dulce. Esta es una característica que ha sido reportada en investigaciones anteriores (Richards et al. 1971, Córdoba & Vargas 1996, Thamdrup et al. 1996, Wolff et al. 1996) y se debe en gran medida a la morfología la cuenca del Golfo Dulce. Esta presenta un umbral poco profundo en la parte externa que restringe la circulación del agua, especialmente en la parte más profunda de la cuenca interna donde, al igual que en fiordos de latitudes altas, se llegan a desarrollar condiciones hipóxicas o anóxicas en ciertas épocas del año (Richards 1965, Rattray 1967, Riley 1967, Thurman 1996). Otro factor que sin duda contribuyó con la fuerte estratificación térmica de la columna de agua es el fenómeno ENOS, uno de cuyos efectos oceánicos más importantes es el incremento de la temperatura en la capa de mezcla del POT (Dietrich 1963, Margalef 1985, Philander 1996, Thurman 1996, NOAA 2000). Dada las altas precipitaciones que se dan en la zona de estudio, también fue posible observar un fuerte gradiente vertical de salinidad, con una capa superficial relativamente diluida y capas profundas con mayores salinidades. Además, se ha observado una estratificación biológica asociada a la estratificación física de la columna de agua, de manera que la mayor parte de los organismos planctónicos en el Golfo Dulce se encuentran distribuidos en la capa superficial cálida y rica en oxígeno (Quesada 2001).
Resulta particularmente interesante el hecho de que las estaciones internas (01, 02 y 03) del Golfo Dulce se diferencien, en cuanto a sus características hidrográficas, de las estaciones externas (04 y 11). Lo anterior coincide con la distribución espacial de la biomasa y la composición del zooplancton en el Golfo Dulce (Quesada 2001) y podría estar asociado a la mayor influencia oceánica a la que está expuesta la parte externa de la cuenca. También se debe considerar el hecho de que las estaciones 04 y 11 presentan profundidades de fondo menores, lo que favorece la presencia de mayores concentraciones de oxígeno disuelto en las aguas profundas. Finalmente, se debe destacar la presencia de la desembocadura del Río Coto-Colorado, el más grande de la cuenca, en las cercanías de la estación 04. Cabe subrayar que, a pesar de que la influencia de dicho río sobre las características oceanográficas de la zona es innegable, las características físicas de las estaciones 04 y 11 no se diferenciaron significativamente (Tukey, p>0.05).
De este modo, la parte externa del Golfo Dulce parece representar un sector de transición entre las condiciones oceánicas de mar abierto y las condiciones costeras en la parte interna de la cuenca. Lo anterior se apoya, una vez más, en los valores de biomasa, abundancia y composición del zooplancton (Quesada 2001) e incluso en la composición de la infauna en la zona externa del umbral (León-Morales & Vargas 1998).
Las distintas características físicas que presentan las masas de agua oceánica con respecto a las costeras, facilitan su identificación al analizar gráficos de isoclinas en una escala temporal (Dietrich 1963, Brown et al. 1989a, b, Thurman 1996). Anteriormente se ha reportado que masas de agua fría, relativamente ricas en oxígeno y de salinidades altas entran al Golfo Dulce en distintos momentos del año (Richards et al. 1971, Hartmann 1994, Thamdrup et al. 1996). Así mismo, se sabe que estos eventos son relativamente lentos y provocan la advección de nutrimentos a la zona fótica (Richards et al. 1971, Brown et al. 1989b, Thamdrup et al. 1996, Wolff et al. 1996, León-Morales & Vargas 1998). Sin embargo, a falta de estudios a largo plazo en la zona, la periodicidad de estas entradas de agua permaneció hasta ahora desconocida. Los datos obtenidos en el presente estudio no solo comprueban el fenómeno de entrada de masas de agua oceánica a la cuenca del Golfo Dulce, sino que permiten observar una marcada periodicidad en la entrada de dichas masas, habiéndose observado eventos en los meses de abril y setiembre de cada año durante el periodo de estudio. Lo anterior resulta evidente al observar las isotermas y las isohalinas de las estaciones 01, 03 y 11 (Figs. 2, 3 y 4), donde se aprecian fluctuaciones importantes en los valores de temperatura y salinidad en los periodos mencionados.
Sin embargo, los fenómenos de entrada de agua no se reflejan con claridad en las isoclinas de oxígeno disuelto. A pesar de que es posible observar incrementos en los valores de oxígeno en las aguas profundas y superficiales que coinciden con los periodos mencionados, dicho patrón no siempre resulta claro ni consistente con las fechas de entrada de agua. Esto se debe a que dicha variable es afectada por una serie de factores tales como el grado de mezcla vertical en la columna de agua, la productividad primaria, la tasa respiratoria de organismos heterótrofos y la acción de organismos descomponedores (Richards et al. 1971, Tilman et al.1982, Ikeda 1985, Reynolds 1989, Thurman 1996). Por lo tanto, cuando la detección de estas masas de agua oceánica no se da en un plazo relativamente corto desde el momento de su entrada a la cuenca, factores como los mencionados vendrían a alterar los valores originales de oxígeno disuelto y a encubrir parcial o totalmente el evento en los gráficos de isoclinas.
Con respecto a la fuerza que mueve los eventos de entrada de masas de agua oceánica al Golfo Dulce, Richards et al. (1971) y Hartmann (1994) sugieren que fenómenos de afloramiento costero estacional podrían ser los responsables de dichos eventos. Sin embargo, hasta la fecha no se ha aportado evidencia que permita sustentar tal afirmación.
El POT se caracteriza por tener aguas muy productivas gracias a la presencia de sistemas de afloramiento como el de Perú (uno de los sistemas más productivos del planeta), el domo de Costa Rica, el afloramiento de Papagayo y el del Golfo de Panamá. Asociado a esto, se ha observado que en el POT se dan típicamente dos máximos en la producción primaria y que el zooplancton varía de forma sincrónica con la clorofila (Longhurst & Pauly 1987). Dessier & Donguy (1985) encontraron que en el Pacífico ecuatorial, entre los 4ºS y los 4ºN, los máximos anuales de clorofila se dan en los meses de agosto (cuando el afloramiento ecuatorial es intenso) y entre marzo y mayo (afloramiento de menor intensidad). Estos incrementos temporales en la clorofila fueron observados por los autores precisamente en zonas de muestreo ubicadas frente a las costas de Panamá. Además, se observaron incrementos en la abundancia de copépodos que coinciden con las fechas anteriormente señaladas.
Los resultados de Dessier & Donguy (1985) coinciden con los datos obtenidos en la presente investigación respecto a la entrada de agua oceánica a la cuenca del Golfo. De hecho, la periodicidad observada en las entradas de agua al Golfo Dulce se ajusta a las fechas de afloramiento ecuatorial observadas por Dessier & Donguy (1985). Por esta razón, resulta razonable suponer que la fuerza que impulsa el sistema de afloramiento frente a las costas de Panamá podría estar asociada o ser la misma que impulsa la entrada de agua oceánica a la cuenca del Golfo Dulce. Más aún, resulta interesante observar que, de acuerdo a los datos obtenidos por Quesada (2001), los principales picos en la biomasa del zooplancton que se observan en cada una de las estaciones coinciden o se dan poco después (un mes) de los periodos en los que se observa la entrada de agua oceánica a la cuenca del Golfo Dulce.
Finalmente, queda abierta la posibilidad de que se den pulsaciones menores de agua oceánica a la cuenca del golfo en fechas distintas a las observadas en este estudio. Lo anterior se desprende de observaciones realizadas por Richards et al. (1971) y Thamdrup et al. (1996) en estudios de corto plazo en la zona. Entre las razones por las que el presente estudio no haya detectado tales pulsaciones se puede citar el efecto que el fenómeno ENOS tiene sobre los sistemas de afloramiento ecuatoriales (Dietrich 1963, Bougis 1976, Margalef 1985, White et al. 1995, Philander 1996, Thurman 1996, Lavaniegos & Gonzalez-Navarro 1999, NOAA 2000). El fenómeno ENOS pudo haber inhibido dichas pulsaciones durante el periodo de estudio, al reducir la intensidad de los fenómenos de afloramiento costero en el POT. Sin embargo, el patrón de entradas de agua observado en esta investigación, por ajustarse a la periodicidad del afloramiento en Panamá y por su consistencia a lo largo de dos años de estudio, probablemente represente los periodos más importantes y estables en el fenómeno de entrada de masas de agua oceánica al Golfo Dulce.
Existen varios aspectos que ponen en evidencia el efecto de ENOS sobre el Golfo Dulce, por ejemplo la marcada reducción en la precipitación durante 1997 (con respecto a años anteriores) (Coen 1983, von Wangelin & Wolff 1996, Quesada 2001) y probablemente una reducción en el caudal de los ríos. Otro punto importante es el hecho de que las temperaturas más bajas que registró la columna de agua durante el periodo de estudio se dieron en el mes de abril de 1997, justo antes de que se reportaran los primeros incrementos asociados a ENOS en la temperatura del agua (3-5ºC) en las estaciones oceanográficas de las islas Galápagos, en el Pacífico Oriental (NOAA 2000). De hecho, en el mes de junio del mismo año, el Golfo Dulce mostró un incremento en la temperatura del agua con respecto a abril, como se evidencia en la profundización de las isotermas (Fig. 2a, 3a y 4a).
Agradecimientos
Deseamos agradecer a Eleazar Ruiz por su ayuda en el campo. Este trabajo es una contribución del proyecto 808-97-243, "Ecología del zooplancton en el Golfo Dulce, Costa Rica, con especial énfasis en copépodos" coordinado por A.M.R., financiado por la Vicerrectoría de Investigación de la Universidad de Costa Rica y formó parte de los requisitos para el grado de M.Sc. en la Universidad de Costa Rica por parte del primer autor.
Resumen
Los patrones de circulación oceánica, la estructura vertical de la columna de agua y la posición vertical de la termoclina son factores de gran importancia para comprender la distribución vertical de los organismos planctónicos en los océanos. Este trabajo estudia la hidrografía del Golfo Dulce, un fiordo tropical en el Pacífico Oriental Tropical (POT), y la posible influencia sobre dicho sistema del ENOS 1997-1998. Se seleccionó un total de cinco estaciones de muestreo, para las cuales se obtuvo, mensualmente, perfiles verticales de temperatura (ºC), salinidad () y oxígeno disuelto (mg O2 /l) durante un periodo de dos años. Los resultados muestran una fuerte estratificación vertical de la columna de agua asociada a la morfología de la cuenca del Golfo Dulce y a la incidencia del fenómeno ENOS. Además, se determinó que existen diferencias significativas en las características hidrográficas de las partes interna y externa de la cuenca del Golfo Dulce. Finalmente, se detectaron eventos bianuales de entrada de agua oceánica fría, con salinidad y concentraciones de oxígeno disuelto relativamente altas, a la cuenca. Estos eventos podrían estar asociados al sistema de afloramiento costero del Golfo de Panamá.
Referencias
Bougis, P. 1976. Marine Plankton Ecology. Elsevier, New York, N.Y. 355 p. [ Links ]
Brown, J., A. Colling, D. Park, J. Phillips, D. Rothery & J. Wright. 1989a. Seawater: its composition, properties and behaviour. Pergamon, Oxford. 238 p. [ Links ]
Brown, J., A. Colling, D. Park, J. Phillips, D. Rothery & J. Wright. 1989b. Ocean Circulation. Pergamon, Oxford. 165 p. [ Links ]
Coen, E. 1983. Climate, p. 35-46. In D. Janzen (ed.). Costa Rican Natural History. University of Chicago. Chicago, Michigan. [ Links ]
Córdoba, R. & J.A. Vargas. 1996. Temperature, salinity, oxygen and nutrient profiles at a 200 m station in Golfo Dulce, Pacific coast of Costa Rica. Rev. Biol. Trop. 44(Suppl. 3): 233-236. [ Links ]
Dessier, A & J.R. Donguy. 1985. Planktonic copepods and environmental properties of the eastern equatorial Pacific: seasonal and spatial variations. Deep Sea Res. 32(9): 1117-1133. [ Links ]
Dietrich, G. 1963. General Oceanography: an introduction. Interscience, New York, 588 p. [ Links ]
Hartmann, H.J. 1994. The Golfo Dulce Marine Environment: Review and Critical Assesment. Department of Biology of La Rochelle, University of La Rochelle, Charente-Maritime, France. 34 p. [ Links ]
Ikeda, T. 1985. Metabolic rates of epipelagic marine zooplankton as a function of body mass and temperature. Mar. Biol. 85: 1-11. [ Links ]
Lavaniegos, B. & E. Gonzalez-Navarro. 1999. Cambios en la comunidad de copépodos durante el ENSO de 1992-93 en el canal de San Lorenzo, Golfo de California. Ciencias Mar. 25(2): 239-265. [ Links ]
León-Morales, R. & J.A. Vargas. 1998. Macroinfauna of a tropical fjord-like embayment: Golfo Dulce, Costa Rica. Rev. Biol. Trop. 46(6): 81-90. [ Links ]
Longhurst, A. 1985. Relationships between diversity and the vertical structure of the upper ocean. Deep Sea Res. 32(12): 1535-1570. [ Links ]
Longhurst, A. & D. Pauly. 1987. Biological Communities of tropical Oceans, p. 63-105. In A.R. Longhurst & D. Pauly (eds.). Ecology of Tropical Oceans. Academic. San Diego. [ Links ]
Margalef, R. 1985. Primary production in upwelling areas: energy, global ecology and resources. Int. Symp. Upw. W. Afr. 5(1): 225-332. [ Links ]
NOAA. 2000. National Oceanographic and Atmospheric Agency. On line at: http://www.noaa.gov/ [ Links ]
Philander, G. 1996. El Niño and La Niña, p. 72-87. In R.G. Pirie (ed). Oceanography: contemporary readings in ocean science. Oxford Univ. New York, N.Y. [ Links ]
Quesada, M.A. 2001. Caracterización, composición, abundancia y biomasa del zooplancton en el Golfo Dulce durante el periodo 1997-1998. Tesis de Maestría, Universidad de Costa Rica, San José. 120 p. [ Links ]
Rattray, M. 1967. Some aspects of the dinamics of circulation in fjords. In G.H. Lauff (ed.). Estuaries. American Association for the Advancement of Science. Washington, D.C. [ Links ]
Reynolds, C.S. 1989. Physical determinants of phytoplankton succession, p. 9- 56. In U. Sommer (ed.). Plankton Ecology: Succesion in plankton communities. Springer. New York. [ Links ]
Richards, F.A. 1965. Anoxic basins and fjords. pp.: 611- 645 In J.P. Riley & G. Skirrow (eds.). Chemical Oceanography, Vol. 1. Academic. London. [ Links ]
Richards, F.A., J.J. Anderson, & J.D. Cline. 1971. Chemical and physical observations in Golfo Dulce, an anoxic basin on the Pacific coast of Costa Rica. Limnol. Oceanogr. 16(1): 43-50. [ Links ]
Riley, G.A. 1967. The Plankton of Estuaries, p. 316-326. In G.H. Lauff (ed.). Estuaries. American Association for the Advancement of Science. Washington. [ Links ]
Svendsen, H., R. Rosseland, S. Myking, J.A. Vargas, O.G. Lizano & E.J. Alfaro. 2004. A physical-Oceanographycal study of Golfo Dulce. Rev. Biol. Trop. 52 (Supl. 3B): 1-10. [ Links ]
Thamdrup, B., D.E. Canfield, T.G. Ferdelman, R.N. Glud, & J.K. Gundersen. 1996. A biogeochemical survey of the anoxic basin Golfo Dulce, Costa Rica. Rev. Biol. Trop. 44 (Suppl. 3): 19-33. [ Links ]
Thurman, H.V. 1996. Essentials of Oceanography. Prentice Hall, New Jersey, New Jersey. 400 p. [ Links ]
Tilman, D., S. Kilham & P. Kilham. 1982. Phytoplankton community ecology: the role of limiting nutrients. Ann. Rev. Ecol. Syst. 13: 349-372. [ Links ]
Umaña, G. 1998. Characterization of some Golfo Dulce drainage basin rivers (Costa Rica). Rev. Biol. Trop. 46(6): 125-135. [ Links ]
von Wangelin, M. & M. Wolff. 1996. Comparative biomass spectra and species composition of the zooplankton communities in Golfo Dulce and Golfo de Nicoya, Pacific coast of Costa Rica. Rev. Biol. Trop. 44 (Suppl. 3): 135-155. [ Links ]
White, J., X. Zang, L. Welling, M. Roman & H. Dam. 1995. Latitudinal gradients in zooplankton biomass in the tropical Pacific at 140ºW during the JGOFS EqPac study: effects of El Niño. Deep Sea Res. II 42(2-3): 715-733. [ Links ]
Wolff, M., H.J. Hartmann, & V. Koch. 1996. A pilot trophic model for Golfo Dulce, a fjord-like tropical embayment, Costa Rica. Rev. Biol. Trop. 44 (Suppl. 3): 215-231. [ Links ]

