










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 n.4 San José Dec. 2004
Hábitos alimentarios de los peces Lutjanus peru y Lutjanus guttatus (Pisces:Lutjanidae)en Guerrero,México
Agustín A.Rojas-Herrera1,Maite Mascaró2 & Xavier Chiappa-Carrara3
1 Escuela Superior de Ecología Marina,Universidad Autónoma de Guerrero.Apdo.Postal 1606,39390 Acapulco, México;rojash56@hotmail.com
2 Laboratorio de Ecología y Biología Marina Experimental,Facultad de Ciencias,Universidad Nacional Autónoma de México.Calle 26 #1,Playa Norte,24140 Ciudad del Carmen,Campeche,México; mmm@hp.fciencias.unam.mx
3 Unidad de Investigación en Ecología Marina,FES –Z.Dirección actual:Unidad Académica Sisal,Universidad Nacional Autónoma de México,Sisal,Yucatán.CP 97355 México; chiappa@servidor.unam.mx
Recibido 21-IX-2002. Corregido 17-III-2003. Aceptado 07-IV-2003.
Abstract: Feeding habits of the fishes Lutjanus peru and Lutjanus guttatus (Pisces:Lutjanidae)of Guerrero, México.Diet composition of the Pacific snapper (Lutjanus peru )130-684 mm fork length (FL)and the spotted snapper (Lutjanus guttatus )120-550 mm FL,was analyzed.Monthly samples were obtained from commercial landings in three regions off the coast of Guerrero,Mexico.Percentage by number (%N),percentage by weight (%W),and percentage of occurrence (%O)were calculated for each prey and summarized as the index of relative importance.Both species are polyphagous predators feeding on a variety of prey:Sixty-eight prey items,mainly fish (%W =50.9),crustaceans (%W =35.6),and mollusks (%W =7.2),were found in the stomach contents of L.peru ,while 88 components were identified in the diet of L.guttatus ,the most important prey being fish (%W =50.8)and crustaceans (%W =43.4).Diet overlap between species is not significant (p<0.05),indicating that competition for food is unlikely.Based on published values of the relative importance index,the diets of these Lutjanidae were analyzed considering different regions of the Pacific coast of Mexico and Costa Rica,and similarities among sites and species were discussed.The cluster analysis showed that similarities among species inhabiting in the same area are more important than within organisms of the same species living in different zones.Results suggest that prey availability rather than food selectivity,conditions the feeding behavior of these fish species.Rev.Biol.Trop.52(4):959-971.Epub 2005 Jun 24.
Key words: Feeding habits,diet,Lutjanidae,Lutjanus peru ,Lutjanus guttatus ,coast of Guerrero, Mexico.
Los pargos (Familia Lutjanidae)representan,junto con las cabrillas (Familia Serranidae, Subfamilia Epinephelinae),un componente importante de las pesquerías,en las zonas tropicales del mundo (Polovina y Ralston 1987). Debido a que las pesquerías de estas zonas son principalmente artesanales,la importancia de estas especies adquiere más relevancia dado el alto número de personas a las que benefician de manera directa,ya sea como una fuente de ingresos o como una fuente de alimentación.
La pesca de lutjánidos en México ascendió en 1999 a 6-439 toneladas,de las cuales 2-994 toneladas se capturaron en las costas del Pacífico (46.5%).En el estado de Guerrero se capturaron 210 toneladas,correspondientes a las especies Lutjanus peru (Nichols y Murphy 1922)y Lutjanus guttatus (Steindachner 1869),de ahí que se le considere una región de pesca importante de este recurso en el litoral del Pacífico mexicano (SEMARNAP 2000).
Existen numerosas publicaciones sobre la biología trófica de la familia Lutjanidae. Entre las que se refieren a L.peru y L. guttatus se encuentran las de Díaz-Uribe (1994),Santamaría-Miranda (1996,1998), Rojas (1997a),Rojas-Herrera (1996),Santamaría-Miranda y Elorduy-Garay (1997),Saucedo-Lozano et al .(1999),Saucedo-Lozano y Chiappa-Carrara (2000),Chiappa-Carrara et al .(2004),Rojas et al.(2004).En estos informes se describe la composición de la dieta de estos organismos de forma aislada y no se aporta información sobre los tipos y las cantidades de presas comunes a ambas especies.
En este trabajo,se analizan los hábitos alimentarios de L.peru y de L.guttatus provenientes de la captura comercial en la costa de Guerrero,México y se estudia el grado de traslape en el uso de los componentes alimentarios de estas especies.Estos datos se comparan con los obtenidos por Saucedo-Lozano et al. (1999),Saucedo-Lozano y Chiappa-Carrara (2000)y Rojas (1997a,b)en otras áreas de distribución de estas especies y,finalmente,se analiza la hipótesis de su coexistencia a través de la exclusión competitiva.
Material y métodos
Entre febrero,octubre-diciembre de 1993 y enero,marzo-diciembre de 1994 se obtuvieron especimenes de Lutjanus peru y L.guttatus en tres regiones de la costa de Guerrero,México.En el Cuadro 1 se muestra la distribución temporal del número de estómagos con alimento analizados para cada especie (L.peru : n =416;L.guttatus :n =239).Las áreas de muestreo fueron Punta Maldonado (16º 18 N y 98º 30 W),el puerto de Acapulco (16 º51 N y 99 º55 W)y Puerto Vicente Guerrero (17º 18 N y 101º 3 W).Los peces provienen de las capturas comerciales nocturnas realizadas a profundidades de 25 a 100 m,utilizando líneas con anzuelos de tamaños "7 "y "8 ".De cada ejemplar se obtuvo la longitud horquilla (LH) con un ictiómetro convencional (± 1 mm)y se extrajeron los tractos digestivos mediante un corte ventral longitudinal,desde el borde superior del opérculo hasta el ano,fijándolos en formaldehído al 10%.En el laboratorio,el contenido de cada estómago se vació en un recipiente y se analizó con un microscopio estereoscópico. Las presas se identificaron hasta el taxon más bajo posible con la ayuda de claves específicas (Jordan y Evermann 1896-1900,Rathbun 1930,Garth 1958,Ramírez-Hernández y Páez-Barrera 1965,Keen 1971,Castro-Aguirre 1978,Yáñez-Arancibia 1978a,b,Brusca 1980,Van der Heiden y Findley 1988,Hendrickx y Salgado-Barragán 1991,Allen y Robertson 1994,Allen 1995, Hendrickx 1995a,1995b,Pérez-Farfante y Kensley 1997,Castro-Aguirre et al .1999). Una vez identificados,los componentes alimentarios presentes en cada estómago,fueron pesados en una balanza electrónica (g ± 0.01 mg).Para efectuar el análisis global de la dieta se agruparon todos los datos obtenidos (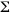 ni ) (Cuadro 1)por cada especie, mientras que para el análisis temporal de la dieta,los datos se agruparon considerando cada uno de los meses de muestreo (ni)(Cuadro 1).
ni ) (Cuadro 1)por cada especie, mientras que para el análisis temporal de la dieta,los datos se agruparon considerando cada uno de los meses de muestreo (ni)(Cuadro 1).

A partir de la enumeración (N)y del peso (P)de los distintos componentes alimentarios se calcularon los porcentajes numéricos (%N),gravimétricos (%P)y de frecuencia de ocurrencia (%FO,Hyslop 1980).La importancia cuantitativa de cada rubro alimenticio se obtuvo mediante el índice de importancia relativa (IIR)propuesto por Pinkas et al.(1971), modificado por Rosecchi y Nouaze (1987)de acuerdo con la expresión:
IIR = %FO (%N + %P)
Para encontrar similitudes en la dieta de estos lutjánidos en distintas áreas de distribución,y con base en los valores del IIR de las presas de L.peru y L.guttatus ,obtenidos en este trabajo,aquellos informados por Saucedo-Lozano et al.(1999)y Saucedo-Lozano y Chiappa-Carrara (2000)para las costas de Jalisco y de Colima,México,así como los de Rojas (1997a,b) para L.guttatus y L.colorado del Golfo de Nicoya,Costa Rica,se realizó un análisis de conglomerados.Se utilizó la métrica de correlación de Pearson y se obtuvieron las agregaciones de los promedios aritméticos de los valores absolutos de correlación,utilizando como medida de distancia el método de los grupos pareados sin ponderación (UPGMA, Rohlf 1963,Visauta-Vinacua 1998).Este algoritmo da peso al número de presas y muestra la mayor correlación cofenética,por lo que su representación gráfica se asemeja a la matriz original (Legendre y Legendre 1998).
Se calcularon los índices de diversidad (B)(Levins 1968)y de traslape de las dietas (Cx)(Horn 1966)y ( xy )(MacArthur y Levins 1967)de acuerdo con las expresiones:
xy )(MacArthur y Levins 1967)de acuerdo con las expresiones:

en las que Pi es la proporción con la cual cada categoría de presa contribuye numéricamente a la dieta de las especies X y Y .
B es un indicador de la amplitud del nicho trófico y comprende valores desde uno (indicando una dieta altamente específica)hasta el número total de categorías alimenticias (k ).Este índice es más robusto que su versión normalizada,cuando se desconoce la abundancia de las presas en el ambiente o cuando éstas no son igualmente abundantes (Feinsinger et al . 1981) .Los valores de este índice son comparables cuando la magnitud de k es mayor que la de B (Hespenheide 1975).
Los valores posibles de Cx se encuentran en el intervalo 0  Cx 1.De acuerdo con Wallace (1981)y Cortés (1997),este índice presenta un sesgo pequeño,en comparación con otras medidas de traslape dietético,cuando el tamaño de la muestra (ni )y el número de presas en el contenido estomacal (k )no son iguales.El índice xy es asimétrico y los valores posibles fluctúan desde 0 hasta >1.En ambos casos, cero representa una separación completa de las dietas mientras que valores = 1 indican un traslape completo.La estimación de los intervalos de confianza (IC, =0.05)de los índices,se efectuó mediante la aplicación del método bootstrap (Mueller y Altenberg 1985, Efron y Tibshirani 1991),realizando 2 000 re-muestreos de los datos (Hamilton 1991). A pesar de que no se han desarrollado procedimientos estadísticos formales para probar la significancia estadística de los índices Cx y xy (Caillet y Barry 1979)y aquellos que existen requieren que se cumplan ciertos supuestos difíciles,como la homoscedasticidad de las varianzas (Maurer 1982),se ha considerado que los valores son biológicamente significativos cuando exceden de 0.60 (Zaret y Rand 1971,Mathur 1977).
Cx 1.De acuerdo con Wallace (1981)y Cortés (1997),este índice presenta un sesgo pequeño,en comparación con otras medidas de traslape dietético,cuando el tamaño de la muestra (ni )y el número de presas en el contenido estomacal (k )no son iguales.El índice xy es asimétrico y los valores posibles fluctúan desde 0 hasta >1.En ambos casos, cero representa una separación completa de las dietas mientras que valores = 1 indican un traslape completo.La estimación de los intervalos de confianza (IC, =0.05)de los índices,se efectuó mediante la aplicación del método bootstrap (Mueller y Altenberg 1985, Efron y Tibshirani 1991),realizando 2 000 re-muestreos de los datos (Hamilton 1991). A pesar de que no se han desarrollado procedimientos estadísticos formales para probar la significancia estadística de los índices Cx y xy (Caillet y Barry 1979)y aquellos que existen requieren que se cumplan ciertos supuestos difíciles,como la homoscedasticidad de las varianzas (Maurer 1982),se ha considerado que los valores son biológicamente significativos cuando exceden de 0.60 (Zaret y Rand 1971,Mathur 1977).
Resultados
Las tallas de los ejemplares analizados se encuentran comprendidas en el intervalo 130 LH 684 mm para el caso del huachinango Lutjanus peru ,y 120 LH 550 mm para el del flamenco L.guttatus. En e1 Cuadro 2 se muestra la composición del espectro alimentario de estas especies en la costa de Guerrero.En los estómagos de L.peru se identificaron 68 ítems alimentarios dentro de los cuales predominan los peces (%P =50.9),los crustáceos (%P = 35.6)y los moluscos (%P = 7.2) mientras que el espectro alimentario de L.guttatus estuvo integrado por 88 componentes que incluyeron principalmente peces (%P =50.8)y crustáceos (%P =43.4).
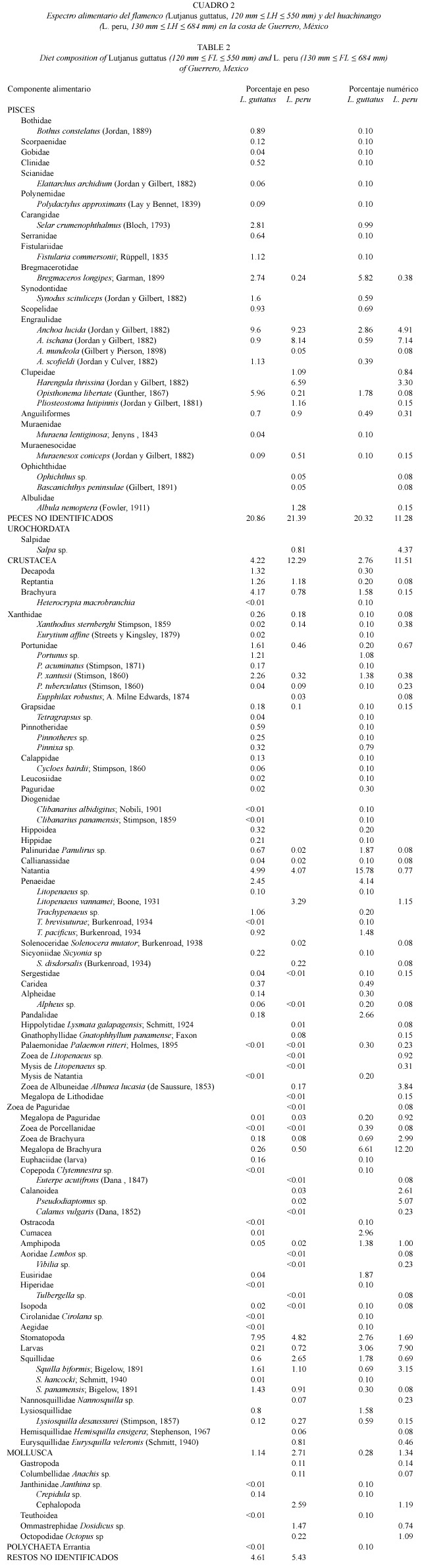
Los valores del índice de diversidad de Levins (1968)muestran que ambas especies presentan una dieta diversa y un número reducido de presas tienen un predominio numérico significativo.En el caso de L.guttatus ,sólo algunos componentes alimentarios (Bregmaceros longipes y Anchoa lucida entre los peces, Stomatopoda,Cumacea,la familia Pandalidae y larvas megalopa de Brachyura entre los crustáceos)sobrepasaron el 2%de la abundancia numérica total.En los contenidos estomacales de L.peru ,las presas con una importancia numérica  2%fueron A.lucida ,A.ischana y Harengula thrissina entre los peces,Salpa sp. (urocordados),las zoeas de Albunea lucasia , las zoeas y megalopas de Brachyura,los copépodos calanoideos (Pseudodiaptomus sp.)y las larvas de estomatópodos (Squilla biformis ) entre los crustáceos.Los valores de diversidad de la dieta no fueron significativamente distintos entre ambas especies (p >0.05),aunque en términos generales,B fue mayor en L.peru (B =13.05 ± 0.41 IC95%)que en L.guttatus (B = 11.95 ± 0.92 IC95%).
2%fueron A.lucida ,A.ischana y Harengula thrissina entre los peces,Salpa sp. (urocordados),las zoeas de Albunea lucasia , las zoeas y megalopas de Brachyura,los copépodos calanoideos (Pseudodiaptomus sp.)y las larvas de estomatópodos (Squilla biformis ) entre los crustáceos.Los valores de diversidad de la dieta no fueron significativamente distintos entre ambas especies (p >0.05),aunque en términos generales,B fue mayor en L.peru (B =13.05 ± 0.41 IC95%)que en L.guttatus (B = 11.95 ± 0.92 IC95%).
Con la finalidad de presentar un análisis más detallado de la dieta de estos lutjánidos, en la Fig.1 se presentan los valores del índice de diversidad calculado en forma mensual. Se observan variaciones temporales significativas (p<0.05)en el régimen alimentario de ambas especies debidas a valores elevados del %N que presentaron algunos componentes alimentarios.Por ejemplo,en el caso de L.peru durante el mes de noviembre de 1993 un solo componente de la dieta (larvas megalopa de Brachyura),presentó un valor numérico elevado (%N =77)y la diversidad,por lo tanto, alcanzó un punto mínimo (B =1.66 ± 1.55 IC95%).Sin embargo,pocas presas tuvieron valores de abundancia relativa altos a lo largo del período de muestreo.
El mismo comportamiento se observó en los datos de la abundancia relativa de las presas de L.guttatus ,considerando la temporalidad del muestreo.En el Cuadro 3 se presenta el listado de los componentes alimentarios cuyos valores de abundancia relativa mensual sobrepasaron el 15%del total.Únicamente presas como Opisthonema libertate ,Selar crumenophthalmus ,B.longipes y otros peces no identificados,así como Natantia,Penaeidae y las larvas megalopa de Brachyura, sobrepasaron el 15%de abundancia relativa.Los peces no identificados,agrupados como Pisces,están presentes con valores elevados de abundancia durante la mayor parte de los meses analizados.En los momentos en que no se presentan los peces en la dieta,son sustituidos por Natantia,Eurisidae y Pandalidea. Crustacea, Penaeidae, B. longipes y las megalopas de Brachyura, son abundantes de septiembre a noviembre de 1994.Vale la pena señalar que en octubre y noviembre de 1993,estos componentes también se presentaron en los contenidos estomacales de L. guttatus y L.peru ,si bien con valores de abundancia menores al 15%.
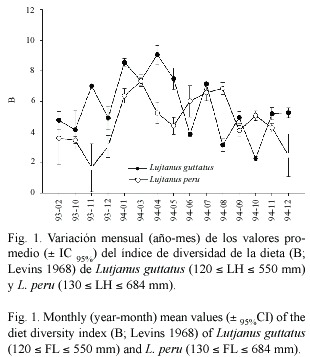
Los valores de los índices de traslape de las dietas obtenidos (±IC 95%)se muestran en el Cuadro 4.De acuerdo con los criterios de Zaret y Rand (1971)y Mathur (1977)la magnitud de los valores promedio no es significativa a pesar de que existen presas comunes a ambas especies como B.longipes ,A.lucida ,A.ischana y Cynoponticus coniceps entre los peces y Portunus xantusii ,P.tuberculatus ,Panulirus sp.,Alpheus sp.,larvas megalopa de Brachyura,larvas de estomatópodos,Squilla biformis y Lysiosquilla desaussurei entre los crustáceos.
0.60)únicamente en diciembre de 1993 y el traslape se debe fundamentalmente a la presencia de "peces no identificados ",en los contenidos estomacales de las dos especies.
Discusión
Los resultados obtenidos en este trabajo muestran que L.peru y L.guttatus se alimentan de una gran variedad de presas,entre las que figuran los peces de las familias Engraulidae y Clupeidae, estomatópodos,penéidos, portúnidos y larvas de crustáceos (zoea y megalopas).En términos generales,estos resultados coinciden con las observaciones de Santamaría-Miranda (1996)sobre la alimentación de L. peru en la costa de Guerrero,las de Saucedo-Lozano et al. (1999)en juveniles de L.peru y las de Saucedo-Lozano y Chiappa-Carrara (2000)en juveniles de L.guttatus de las costas de Jalisco y Colima.Díaz-Uribe (1994),en las bahías de La Paz y La Ventana,encontró que L.peru se alimenta principalmente de organismos planctónicos,casi exclusivamente de colonias de urocordados (doliólidos y salpas)y de crustáceos (principalmente eufáusidos en el caso de los organismos adultos y carídeos,misidáceos y estomatópodos en el de los juveniles).Por último,Rojas (1997a) reporta que la alimentación de L.guttatus en el Golfo de Nicoya en Costa Rica,está compuesta por tres familias de peces (Engraulidae, Centropomidae y Ophychthidae)y 5 familias de crustáceos,entre las que domina Penaeidae.Según este autor,L.guttatus se comporta como un depredador béntico.
Es necesario señalar que los organismos analizados en este trabajo poseen un espectro alimentario más amplio que el reportado por los autores citados.Este hecho puede estar relacionado con las variaciones geográficas de la abundancia y disponibilidad de las presas de las que se alimentan estas especies.De hecho, los valores de similitud que se obtuvieron en el análisis de conglomerados (Visauta-Vinacua 1998),aplicado a los valores porcentuales del IIR (Pinkas et al .1971),arrojaron dos grupos bien definidos de afinidad.El primero está integrado por L.peru y L.guttatus de la costa de
Guerrero y por los juveniles de L.peru y L.guttatus de la costa de Jalisco y Colima;mientras que el segundo,por L.guttatus y L.colorado del Golfo de Nicoya,Costa Rica (Fig.3).La similitud observada entre las dietas de L.peru y L.guttatus provenientes del litoral de Guerrero está determinada por el alto consumo de peces.En Jalisco y Colima,los juveniles presentan alto consumo de crustáceos (Amphipoda en el caso de L.peru y estadios juveniles de penéidos en el de L.guttatus )mientras que en Costa Rica,tanto L.guttatus como L.colorado, mostraron un consumo importante de Natantia (principalmente Penaeidae).

L.peru y L.guttatus son depredadores generalistas,capaces de ingerir un amplio espectro de especies,fundamentalmente peces y crustáceos.Al parecer,es la disponibilidad de las presas en cada región,más que la selección de presas,la que determina la alimentación de estas especies.Esto explica el hecho de que se haya encontrado más afinidad entre las dietas de aquellas especies que habitan en un mismo sitio,que entre organismos de la misma especie presentes en diferentes localidades. En el mismo sentido,trabajos previos han demostrado que las variaciones en la composición dietética de otro lutjánido,L.apodus ,dependen de las características del hábitat (Rooker 1995).Cabe señalar,sin embargo,que no queda descartado que estos lutjánidos puedan exhibir una preferencia alimentaria por algún tipo o tamaño de presa en particular,y que con ello revelen ciertos rasgos de selectividad.La preferencia alimentaria de depredadores oportunistas tiene una base conductual relacionada con los estímulos que el depredador percibe de su presa,como el olor,el movimiento,el color o la abundancia (Kaiser et al.1993).Estos estímulos constituyen piezas de información para la toma de decisiones individuales sobre qué presa atacar e ingerir (Bouskila y Blumstein 1992)y frecuentemente,están asociados al valor energético que ésta puede retribuirle al depredador (Pyke et al .1977,Hughes 1980).
El análisis del espectro alimentario de estas especies muestra que no existen variaciones cualitativas estacionales en el uso de los distintos componentes de la dieta.A lo largo del período de estudio,se encontraron los mismos grupos de presas en el contenido estomacal de L.guttatus y L.peru .Sin embargo,existen diferencias cuantitativas:el mayor consumo de presas planctónicas (larvas de decápodos,por ejemplo),ocurrió en los meses de verano (julio)prolongándose hasta noviembre-diciembre, que en la zona de estudio corresponde al periodo en el que se registra el ~80%de la precipitación total anual (Meli 1998).Este hecho puede estar relacionado con elevaciones en la abundancia de presas planctónicas,debidas al aumento de la producción primaria,como consecuencia del mayor flujo de nutrimentos de la tierra a la zona costera.El incremento en la representación proporcional de ciertos componentes dietéticos,conforme aumenta su abundancia relativa en el medio, constituye una evidencia más en el sentido de que la disponibilidad de las presas determina, de manera importante,los hábitos alimentarios de estos lutjánidos.
La evaluación del traslape dietético entre las especies estudiadas se realizó mediante la aplicación de dos índices.Si bien los valores de Cx y de xy calculados globalmente son similares (p>0.05),la utilización del índice asimétrico de traslape de MacArthur y Levins (1967)muestra que la dieta de L.guttatus se traslapa con más intensidad a la de L.peru (Fig.2B).Si se considera,utilizando como referencia los valores obtenidos en el mes de diciembre de 1993,que las estimaciones de B son menores en L.peru ,es posible plantear la existencia de una correspondencia unidireccional en la utilización de los recursos alimentarios:en el caso de L.guttatus ,la dieta es más diversa y los grupos alimentarios son utilizados en mayor cantidad.
Si bien Holt (1987)indica que los índices de traslape de dietas miden el grado de utilización conjunta de un recurso por un par de especies,y demostró que la relación entre el traslape y la competencia es compleja,la interpretación de dichos índices permite inferir algunos patrones comunitarios generales. Los valores de traslape de las dietas de L.peru y L.guttatus sugieren que,a pesar de la similitud existente en los componentes alimentarios de estas especies,no existe un alto grado de competencia interespecífica por el alimento, apoyando la idea de Holt (1987),de que la condición multidimensional del nicho determina la aptitud de una especie con relación a la otra. Los resultados informados por Sierra (1997) para las especies de lutjánidos que habitan en el Golfo de Batabanó,Cuba,son consistentes con este análisis.Morton et al .(1988)e Ibáñez-Aguirre (1993),plantean que el traslape de las dietas entre dos especies ocurre cuando los recursos son escasos o poco diversos.De esta forma,es posible suponer que los recursos alimentarios,y en particular aquellas especies de presas que ambos lutjánidos comparten en el litoral de Guerrero,son lo suficientemente diversos y abundantes como para permitir la coexistencia de estas especies que utilizan distintos componentes del hábitat,como una estrategia para disminuir la competencia entre ellas.
Esta aparente contradicción con la idea de Gauze (1934)de que dos especies que utilizan los mismos recursos no consiguen coexistir, puede ser explicada por dos argumentos.Por una parte,las diferencias temporales en la dieta de estos peces pueden estar en sincronía con los patrones de producción estacionales o con los ciclos reproductivos de cada especie, dando como resultado una repartición de los recursos alimentarios que permite la coexistencia de ambas especies (Pianka 1974).En este caso, Weatherley (1963)menciona que la competencia no es sino un problema efímero considerando la naturaleza plástica de la dieta de estos peces.
Por otra parte,dichas diferencias pueden tener una componente evolutiva que ha sido poco estudiada.Rooker (1985)y Saucedo-Lozano (2000)encontraron que existen diferencias ontogenéticas en la morfología mandibular de Lutjanus apodus (70 LH 420 mm),L.peru (41 longitud total (LT) 303 mm)y L.guttatus (66 LT 341 mm). Las diferencias morfométricas,tanto ontogenéticas como interespecíficas,en los huesos dentarios y premaxilares de estos lutjánidos, pueden estar relacionadas con las variaciones dietéticas que presentan estas especies.
Agradecimientos
El presente trabajo fue posible gracias al apoyo del CONACYT a través de la Dirección de Asuntos Internacionales dentro del programa de cooperación México-Cuba 1994 y al financiamiento obtenido mediante el proyecto PAPIIT IN211999.Las sugerencias de cuatro revisores anónimos permitieron que el manuscrito adquiriera su forma definitiva.
Resumen
Se analiza la composición de la dieta del huachinango (Lutjanus peru )y del flamenco (Lutjanus guttatus ) en la costa de Guerrero,México.Los ejemplares se obtuvieron mensualmente de las capturas comerciales en tres regiones de pesca y presentaron intervalos de talla comprendidos entre 130 y 684 mm de longitud horquilla (LH)para el caso de L.peru ,y de 120 a 550 mm para L. guttatus .A partir del número y del peso de los distintos componentes alimentarios,identificados hasta el taxon más bajo posible,se calcularon los porcentajes numéricos (%N),gravimétricos (%P)y de frecuencia de ocurrencia (%FO)que fueron sintetizados como valores de importancia relativa.En los estómagos de L.peru se identificaron 68 ítems alimentarios dentro de los cuales predominan los peces (%P =50.9),los crustáceos (%P =35.6)y los moluscos (%P =7.2)mientras que el espectro alimentario de L.guttatus estuvo integrado por 88 componentes que incluyeron principalmente peces (%P =50.8)y crustáceos (%P =43.4).Ambas especies son depredadoras generalistas,con un amplio espectro de presas.Los valores de los índices de traslape de las dietas de estas especies sugieren que no existe un alto grado de competencia intraespecífica por el alimento.Con el fin de encontrar similitudes en la dieta de estos lutjánidos en varias localidades de México y Costa Rica,se realizó un análisis de conglomerados con los valores de importancia relativa de las presas,publicados en varios trabajos.Se encontró más afinidad en la dieta de aquellas especies que habitan en una misma localidad, que entre organismos de la misma especie distribuidos en diferentes zonas.Se infiere que la disponibilidad de las presas,más que la selectividad por el alimento,determina la dieta de estas especies.
Palabras clave:Hábitos alimenticios,dieta,Lutjanidae, Lutjanus peru ,Lutjanus guttatus, Guerrero, México.
Referencias
Allen,G.R.1995.Lutjanidae,p.1231-1244.In W.Fisher, F.Krupp,W.Schneider,C.Sommer,K.E.Carpenter & V.H.Niem (eds.).Guía FAO para la identificación de especies para los fines de la pesca,Pacífico Centro-Oriental,Vol.3. [ Links ]
Allen,G.R.&D.R.Robertson.1994.Fishes of the Tropical Eastern Pacific.University of Hawaii,Honolulú. 322 p. [ Links ]
Bouskila,A.&D.T.Blumstein.1992.Rules of thumb for predation hazard assessment:predictions from a dynamic model.Am.Nat.139:161-176. [ Links ]
Brusca,R.C.1980.Common intertidal invertebrates of the Gulf of California.University of Arizona,Tucsón. 153 p. [ Links ]
Caillet,G.M.&J.P.Barry.1979.Comparison of food array overlap measures useful in fish feeding habit analysis,p.67-79.In S.J.Lipovsky &C.A.Simenstad (eds.)Fish Food Habit Studies:Proceedings of the 2nd Pacific Northwest Technical Workshop.Washington Sea Grant Program,University of Washington, Seattle. [ Links ]
Castro-Aguirre,J.L.1978.Catálogo de peces marinos que penetran a las aguas continentales de México con aspectos zoogeográficos y ecológicos.Departamento de Pesca,Dirección General del Instituto Nacional de la Pesca,México.228 p. [ Links ]
Castro-Aguirre,J.L.,H.S.Espinoza-Pérez &J.J.Schmitter-Soto.1999.Ictiofauna estuarino-lagunar y vicaria de México. Limusa, México.711 p. [ Links ]
Chiappa-Carrara,X.,A.A.Rojas-Herrera &M.Mascaró. 2004.Coexistencia de Lutjanus peru y Lutjanus guttatus (Pisces:Lutjanidae)en la costa de Guerrero, México:relación con la variación temporal en el reclutamiento. Rev. Biol. Trop. 52:177-185. [ Links ]
Cortés,E.1997.A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes.Can.J.Fish. Aquat.Sci.54:726-738. [ Links ]
Díaz-Uribe,J.C.1994.Análisis trofodinámico del huachinango Lutjanus peru (Nichols y Murphy,1922)en las bahías de La Paz y La Ventana,B.C.S.México.Tesis de maestría,Centro de Investigaciones Científicas y de Educación Superior de Ensenada,México.57 p. [ Links ]
Efron,B.,&R.Tibshirami.1991.Statistical analysis in computer age.Science 253:390-395. [ Links ]
Feinsinger,P.,E.E.Spears &R.W.Poole.1981.A simple measure of niche breadth.Ecology 62:27-32. [ Links ]
Garth,J.S.1958.Brachyura of the Pacific coast of America: Oxyrhyncha.Allan Hancock Pac. Exped. 21:1-854. [ Links ]
Gause,G.F.1934.The struggle for existence.Hafner,New York.163 p. [ Links ]
Hamilton,L.C.1991.Bootstrap programming.Stata Tech. Bull.4:18-27. [ Links ]
Hendrickx,M.E.1995a.Camarones,p.417-528.In W. Fisher,F.Krupp,W.Schneider,C.Sommer,K.E. Carpenter &V.H.Niem (eds.).Guía FAO para la identificación de especies para los fines de la pesca, Pacífico Centro-Oriental,Vol.1. [ Links ]
Hendrickx,M.E.1995b.Cangrejos,p.565-630.In W. Fisher,F.Krupp,W.Schneider,C.Sommer,K.E. Carpenter &V.H.Niem (eds.).Guía FAO para la identificación de especies para los fines de la pesca, Pacífico Centro-Oriental,Vol.1. [ Links ]
Hendrickx,M.E.&J.Salgado-Barragán 1991.Estomatópodos (Crustacea:Hoplocarida)del Pacífico mexicano. An. Inst. Cienc. Mar Limnol.,Univ.Nal.Autón. México,Pub.Esp.20.200 p. [ Links ]
Hespenheide,H.A.1975.Prey characteristics and predator niche width,p.158-180.In M.L.Cody &J.M. Diamond (eds.).Ecology and evolution of communities.Harvard University. [ Links ]
Holt,R.D.1987.On the relation between niche overlap and competition:the effect of incommensurable niche dimensions. Oikos 48:110-115. [ Links ]
Horn,H.S.1966.Measurement of overlap in comparative ecological studies.Am.Nat.100:419-424. [ Links ]
Hughes,R.N.1980.Optimal foraging theory in the marine context.Ocean.Mar.Biol.Ann.Rev.18:423-481. [ Links ]
Hyslop,E.V.1980.Stomachs contents analysis -a review of methods and their application. J. Fish. Biol.50: 324-340. [ Links ]
Ibáñez-Aguirre,A.L.1993.Coexistence of Mugil cephalus and M.curema in a coastal lagoon in the Gulf of Mexico. J. Fish. Biol.42:959-961. [ Links ]
Jordan,D.S.&B.W.Evermann.1896-1900.The fishes of North and Middle America.Bull.US Nat.Mus. 47:1-3313. [ Links ]
Kaiser,M.J.,R.N.Hughes &R.N.Gibson.1993.Factors affecting diet selection in the shore crab,Carcinus maenas (L.).Anim.Behav.45:83-92 [ Links ]
Keen,M.A.1971.Sea shells of tropical West America. Marine mollusks from Baja California to Perú. Stanford University. 104 p. [ Links ]
Legendre,P.&L.Legendre.1998.Numerical ecology. Elsevier.853 p. [ Links ]
Levins,R.1968.Evolution in changing environments:some theoretical explorations.Princeton University.120 p. [ Links ]
MacArthur,R.&R.Levins.1967.The limiting similarity, convergence,and divergence of coexisting species. Amer. Nat. 101:377-385. [ Links ]
Mathur,D.1977.Food habits and competitive relationships of the bandfin shiner in Halawakee Creek, Alabama. Am. Mid. Nat.9:89-100. [ Links ]
Maurer,B.A.1982.Statistical inference for Mac Arthur- Levins niche overlap.Ecology 63:1712-1719. [ Links ]
Meli,R.1998.Damage in Acapulco by hurricane Pauline. Nat.Hazards Obs.22:1-2. [ Links ]
Morton,R.M.,J.P.Beumer &B.R.Pollock.1988.Fishes of a subtropical Australian saltmarsh and their predation upon mosquitoes.Environ.Biol.Fish.21:185-194. [ Links ]
Mueller,L.D.&L.Altenberg.1985.Statistical inference on measures of niche overlap.Ecology 66:1204-1210. [ Links ]
Pérez-Farfante,I.&B.Kensley.1997.Penaeoid and sergestoid shrimps and prawns of the world.Keys and diagnoses for the families and genera.Mem.Mus. Nat.Hist.Nat.Paris 175:1-233. [ Links ]
Pianka,E.R.1974.Niche overlap and diffuse competition. Proc.Nat.Acad.Sci.USA 71:2141-2145. [ Links ]
Pinkas,L.,M.S.Oliphant &I.L.K.Iverson.1971.Food habits of albacore,bluefin tuna and bonito in California waters. Calif. Dep. Fish Game Fish.Bull.152:1-105. [ Links ]
Polovina,J.J.&S.Ralston.1987.Tropical snappers and groupers.Biology and fisheries management.Ocean Resour. Mar. Policy Ser.,Westview,Boulder.659 p. [ Links ]
Pyke,G.H.,H.R.Pulliam &E.L.Charnov.1977.Optimal foraging:a selective review of theory and tests. Quart. Rev. Biol. 52:137-154. [ Links ]
Rathbun,M.J.1930.The cancroid crabs of America of the families Eurylidae, Portunidae, Atelecyclidae, Cangridae, and Xanthidae.Bull.US Nat.Mus.152:1-593. [ Links ]
Ramírez-Hernández,E.&J.Páez-Barrera.1965.Investigaciones ictiológicas en las costas de Guerrero 1. Lista de peces marinos de Guerrero colectados en el período 1961-1965. An. Inst. Nal. Invest. Biol. Pesq. 1:329-360. [ Links ]
Rohlf,F.J.1963.Classification of Aedes by numerical taxonomy methods (Diptera:Culicidae). Ann.Entomol. Soc. Am. 56:798-804. [ Links ]
Rojas,M.J.R.1997a.Hábitos alimentarios del pargo mancha Lutjanus guttatus (Pisces:Lutjanidae)en el Golfo de Nicoya,Costa Rica.Rev.Biol.Trop. 44/45:471-476. [ Links ]
Rojas,M.J.R.1997b.Dieta del "pargo colorado "Lutjanus colorado (Pisces:Lutjanidae)en el Golfo de Nicoya, Costa Rica.Rev.Biol.Trop.45:1173-1183. [ Links ]
Rojas M.J.R.,E.Maravilla &F.Chicas B.2004.Hábitos alimentarios del pargo mancha Lutjanus guttatus (Pisces:Lutjanidae)en Los Cóbanos y Puerto La Libertad,El Salvador.Rev.Biol.Trop.52:163-170. [ Links ]
Rojas-Herrera,A.A.1996.Análisis de la alimentación del flamenco Lutjanus guttatus (Steindachner,1869) (Pisces:Lutjanidae)de la costa de Guerrero,México.Mem.Primer Encuentro Regional sobre Investigación y Desarrollo Costero:Guerrero,Oaxaca y Chiapas.p.10. [ Links ]
Rooker,J.R.1995.Feeding ecology of the schoolmaster snapper,Lutjanus apodus (Walbaum),from Southwestern Puerto Rico.Bul.Mar.Sci.56:881-894. [ Links ]
Rosecchi,E.&Y.Nouaze.1987.Comparaison de cinq indices alimentaires utilisés dans l analyse des contenus stomacaux. Rev.Trav.Inst.Pêches Marit.49: 111-123. [ Links ]
Santamaría-Miranda,A.1996.Hábitos alimenticios del huachinango Lutjanus peru (Nichols y Murphy,1922)de la Costa Chica de Guerrero.Mem.X Congreso Nacional de Oceanografía, Manzanillo, México.p.59. [ Links ]
Santamaría-Miranda,A.1998.Hábitos alimenticios y ciclo reproductivo del huachinango Lutjanus peru (Nichols y Murphy,1922)(Pisces:Lutjanidae)en Guerrero,México.Tesis de maestría,CICIMAR,La Paz,México.64 pp. [ Links ]
Santamaría-Miranda,A.&J.F.Elorduy-Garay.1997. Hábitos alimenticios del huachinango Lutjanus peru (Nichols y Murphy,1922)de la costa de Guerrero. México.Mem.V Congreso Nacional de Ictiología, Mazatlán,México.p.54. [ Links ]
Saucedo-Lozano,M.,C.González-Sansón &X.Chiappa- Carrara.1999.Alimentación natural de juveniles de Lutjanus peru (Nichols y Murphy,1922)(Lutjanidae:Perciformes)en la costa de Jalisco y Colima, México.Cienc.Mar.25:381-400. [ Links ]
Saucedo-Lozano,M.&X.Chiappa-Carrara.2000.Alimentación natural de juveniles de Lutjanus guttatus (Pisces:Lutjanidae)en la costa de Jalisco y Colima, México.Bol.Centro Invest.Biol.34:159-180. [ Links ]
SEMARNAP.2000.Anuario Estadístico de Pesca 1999.Secretaría del Medio Ambiente,Recursos Naturales y Pesca, México.(También disponible en línea http://regpesc.semarnap.gob.mx:8891/estad/anua99/c98a05.xls). [ Links ]
Sierra,L.M.1997.Relaciones tróficas de los juveniles de cinco especies de pargo (Pisces:Lutjanidae)en Cuba. Rev. Biol. Trop. 44/45:499-506. [ Links ]
Wallace,R.K.Jr.1981.An assessment of diet-overlap indexes.Trans Am.Fish.Soc.110:72-76. [ Links ]
Weatherley,A.H.1963.Notions of niche and competition among animals,with special reference to freshwater fish.Nature 197:14-17. [ Links ]
Van der Heiden,A.M.&L.T.Findley.1988.Lista de peces marinos del sur de Sinaloa, México. An. Inst. Cienc. Mar Limnol.UNAM 15:209-224. [ Links ]
Visauta-Vinacua,B.1998.Análisis estadístico con SPSS para Windows (Volumen II Estadística multivaríante).McGraw-Hill,España.358 p. [ Links ]
Yáñez-Arancibia,A.1978a.Taxonomía,ecología y estructura de las comunidades de peces en las lagunas costeras con bocas efímeras del Pacífico de México.An.Inst.Cienc.Mar Limnol.UNAM, Pub.Esp.2.306 p. [ Links ]
Yánez-Arancibia,A.1978b.Patrones ecológicos y variación cíclica de la estructura trófica de las comunidades nectónicas en lagunas costeras del Pacífico de México.Rev.Biol.Trop.Vol.26 (Supl.1):189-218. [ Links ]
Zaret,T.M.&A.S.Rand.1971.Competition in tropical stream fishes:support for the competitive exclusion principle.Ecology 52:336-342. [ Links ]

