











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.52 no.1 San José mar. 2004
Actividad de las fosfatasas ácidas y alcalinas
(extracelulares e intracelulares) en hongos de la rizosfera de
Arachis hypogaea (Papiloneaceae)
Y. Gómez-Guiñán
Universidad de Oriente, Núcleo de Anzoátegui. Departamento de Ciencias, Laboratorio de Investigaciones Biológicas. Apdo. Postal 1131, Lechería, Estado Anzoátegui, Venezuela. Fax: 0208-281-92-39; irmagomez52@hotmail.com
Recibido 11-V-2001. Corregido 15-VIII-2002. Aceptado 07-II-2003.
Abstract
The potential role of the fungi, isolated from the peanut rhizosphere, in the production of extracellular and intracellular acid and alkaline phosphatase, was evaluated in vitro. Acid and alkaline extracellular phosphatases showed the highest activities, and the Penicillium species were the most efficient in their production. The correlation analysis showed that extracellular acid and intracellular acid phosphatase produced by Aspergillus niger, A. terreus, Penicillium sp. y P. brevicompactum were negatively correlated; while the extracellular and intracellular phosphatase activities, were positively correlated. The extracellular acid phosphatase activities produced in vitro by majority of fungi assayed, were not correlated with the acid phosphatase activity present in the peanut soil rhizosphere. Nevertheless, the extracellular alkaline phosphatase activities produced in vitro, were negatively correlated with the extracellular alkaline phosphatase activities present in the rhizosphere. The ability of phosphatase production by fungi isolated from peanut rhizosphere suggests they have great potential to contribute to the P mineralization in this zone.
Key words: Arachis hypogaea, acid phosphatase, alkaline phosphatase, extracelular phosphatase, intracelular phosphatase, fungi, rhizosphere.
La importancia de los microorganismos en ambientes naturales, deriva de su diversidad y sobre todo de su gran espectro de actividades, las cuales en la mayoría de los casos repercuten en el desarrollo de las plantas. En este sentido, la presencia de microorganismos con gran potencial en la mineralización del fósforo, especialmente en suelos de baja fertilidad, juegan un papel importante en la disponibilidad de este elemento para las plantas.
En la rizosfera, la presencia de un gran depósito de fósforo orgánico, el cual no puede ser utilizado por las plantas (Curl y Truelove 1986), enfatiza el papel de la flora microbiana en la producción de diferentes isoenzimas de las fosfatasas, involucradas en la mineralización del P, las cuales parecen estar controladas poligenéticamente (Kropp 1990).
La mineralización del fósforo orgánico está mediada por las enzimas fosfatasas. Estas pueden ser sintetizadas, tanto por las raíces de las plantas, las cuales producen fosfatasas ácidas (Ridge y Rovira 1971, Shaykh y Roberts 1974, Dinkelaker y Marschener 1992), como por la flora micótica y las bacterias capaces de producir fosfatasas ácidas y alcalinas (Dighton 1983, Tarafdar y Classen 1988).
Aunque gran parte de la actividad mineralizadora del P orgánico ha sido atribuída a las micorrizas (Ho y Zak 1979, Ho 1988, Antibius et al. 1992, Joner y Johansen 2000), se ha informado, que los microorganismos involucrados en procesos de descomposición, también son capaces de producir fosfatasas (Dighton 1983). Por lo que el papel potencial de las enzimas producidas por estos organismos en la mineralización de compuestos orgánicos de fósforo es de gran importancia ecológica.
Beever y Burns (1980), sugirieron la existencia de tres tipos de fosfatasas (ácida, alcalina y las fosfatasas con alta especificidad por el sustrato), en una gran variedad de hongos. Dada la importancia de esta característica de la flora micótica, su presencia en la zona de la rizosfera resulta de gran valor para el crecimiento de las plantas. A pesar de la importancia de este aspecto, desde el punto de vista agroecológico, las investigaciones sobre la capacidad de producción de fosfatasas por parte de los hongos rizosféricos no micorrizantes ha recibido poca atención.
El objetivo de la presente investigación, fue el de evaluar el papel potencial de hongos saprófitos, aislados de la rizosfera del maní, en la mineralización del fósforo orgánico, mediante la cuantificación in vitro de la producción de fosfatasas ácidas y alcalinas (extracelulares e intracelulares) y relacionar sus actividades entre sí y con las actividades producidas en el suelo de la rizosfera.
Materiales y métodos
La determinación de la actividad de las fosfatasas ácidas y alcalinas (extracelulares e intracelulares), se realizó por triplicado en hongos aislados de la rizosfera de Arachis hypogaea L., cultivado en suelo ácido (pH 5.4) de la Mesa de Guanipa, Estado Anzoátegui, Venezuela (8º47N y 64º13W). Su aislamiento se llevó a cabo mediante la técnica de dilución en placa, empleando el medio de cultivo agar Czapek-Dox (Gómez-Guiñán y Nageswara 1996).
Determinación de las colonias activas en la producción de fosfatasas: Las colonias activas en la producción de fosfatasas, se determinaron siguiendo el método descrito por Nahas et al. (1994). Para ello, los hongos previamente aislados de la rizosfera se inocularon en el medio de crecimiento agar Czapek-Dox y se incubaron por 3 días a 27ºC. Posteriormente, las placas fueron inundadas con una solución de p-nitrofenilfosfato (0.025 M), en buffer Universal (0.17 M) ajustado a pH 5.5 y pH 9.0 para la determinación cualitativa de las fosfatasas ácidas y alcalinas, respectivamente. Los medios inoculados se incubaron por 90 minutos a 37ºC y luego se sometieron a vapores de NaOH. La visualización de halos de color amarillo alrededor de las colonias, indicó la presencia de fosfatasas en el medio de cultivo.
Los hongos que resultaron positivos para la producción de fosfatasas ácidas y alcalinas, fueron seleccionados para la posterior determinación de sus capacidades en la producción de fosfatasas extracelulares e intracelulares.
Condiciones de cultivo: El cultivo de los hongos seleccionados se realizó por triplicado, mediante la inoculación de tacos de micelio de 5 mm de diámetro (1 por cada aislado) provenientes de cultivos jóvenes, en frascos con 50 ml de medio líquido, el cual contenía: 0.05 g de CaCl2 , 0.025 g de NaCl, 0.01 g de KH2 PO4 , 0.5 g de MgSO4 (7H2O), 0.003 g de ZnSO4 (7H2O), 0.001 g de MnSO4 (H2O), 1 g de extracto de levadura, 10 g de glucosa, 1 ml de FeCl3 y 1 000 ml de agua destilada. El pH del medio fue ajustado a pH 5.5. El crecimiento bacteriano fue inhibido mediante el uso de 1 ml de cloranfenicol (50 mg–1 ml). El cultivo fue incubado por 14 días, a 27ºC, en la oscuridad y en constante agitación (250 rpm).
Ensayo enzimático: Finalizado el período de incubación, el micelio de cada hongo fue cosechado mediante la filtración del medio de cultivo a través de un papel filtro Whatman Nº 2. El volumen del filtrado fue completado a 50 ml con agua destilada estéril. Este fue empleado para la determinación de las actividades de las fosfatasas ácidas y alcalinas extracelulares. El micelio cosechado, fue lavado tres veces con Buffer Universal y colocado sobre papel filtro para eliminar el exceso del mismo. Posteriormente, éste fue transferido a un recipiente con 20 ml del buffer universal (0.17 M) ajustado a pH 5.5 y 9.0 para la determinación de las fosfatasas ácidas y alcalinas intracelulares, respectivamente. El micelio fue homogenizado a 800 rpm por 30 segundos. A 3 ml de la suspensión de micelio se le añadió 1 ml de p-nitrofenilfosfato (0.025 M). Las mezclas de reacción se incubaron por 3 horas a 37ºC, y se centrifugaron a 2 000 g durante 10 minutos. A 0.5ml del sobrenadante le fue añadido 4.5 ml de NaOH (0.5 M). La cantidad de p-nitrofenol liberado fue determinada midiendo la absorbancia de la solución a 410 nm. Como controles se usaron medios en los cuales no se hizo crecer el micelio fúngico.
La determinación de la actividad de las fosfatasas ácidas y alcalinas extracelulares se realizó en forma similar a la descrita anteriormente, usando 3 ml del filtrado del medio de cultivo. En este caso, el control consistió en un medio hervido por 15 minutos y 1 ml de p-nitrofenilfosfato (Antibius et al. 1992).
El micelio cosechado fue secado a 80ºC hasta obtener un peso constante. Los resultados de las actividades enzimáticas se expresaron en mg de p-nitrofenol mg-1 de micelio seco h-1 a 37ºC.
Determinación de la actividad de las fosfatasas en la rizosfera: Una vez obtenidas las muestras de la rizosfera (Gómez-Guiñán y Nagueswara 1996), la actividad de las fosfatasas en esta zona fueron determinadas mediante incubación de las muestras (1 g) con Buffer Universal (0.17 M), ajustado a pH 5.5 y 9.0, para la determinación de las fosfatasas ácidas y alcalinas respectivamente; como sustrato se empleó 1 ml de una solución de p-nitrofenilfosfato (0.025 M) (Tabatabai y Bremner 1969). Para evitar la dispersión de las arcillas y extraer el p-nitrofenol del suelo, se emplearon 1 ml de CaCl2 (0.5M) y 4 ml de NaOH (0.5M), respectivamente (Burns 1978). Las mezclas se centrifugaron a 3 000 g por 10 min y se filtraron a través de un papel Whatman Nº 2. La formación de p-nitrofenol fue determinada por espectrofotometría a 410 mn.
Análisis estadísticos: Los resultados fueron sometidos a un análisis de varianza (ANOVA), para establecer diferencias significativas entre las actividades de las fosfatasas extracelulares e intracelulares, ácidas y alcalinas. Un análisis de regresión lineal permitió determinar la relación existente entre las fosfatasas extra e intracelulares producidas in vitro y entre éstas y las obtenidas en la rizosfera del maní.
Resultados
Los hongos aislados de la rizosfera del maní, que in vitro presentaron capacidad mineralizadora del P, pertenecieron a los géneros Aspergillus (Aspergillus sp., A. niger, A. terreu) y Penicillium (Penicillium sp., P. b r evicompactum y P. chrysogenum). Éstos variaron en sus capacidades para producir fosfatasas ácidas y alcalinas extracelulares e intracelulares. Las especies de Penicillium fueron más activas en la producción de fosfatasas ácidas y alcalinas extracelulares y fosfatasa ácida intracelular. (Cuadro 1).
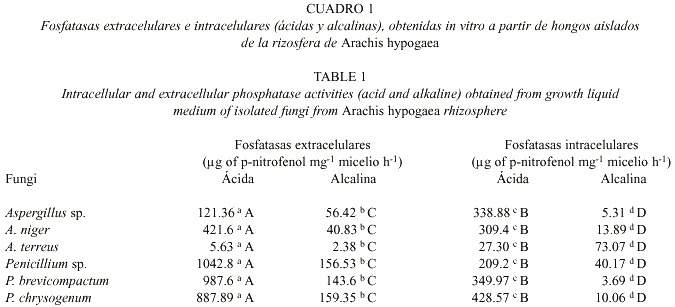
Al comparar la actividad de las fosfatasas ácidas extracelulares e intracelulares y las fosfatasas alcalinas extracelulares e intracelulares (Cuadro 1), se observa que, las fosfatasas ácidas y alcalinas producidas extracelular e intracelularmente, difirieron significativamente entre sí para los diferentes hongos ensayados, resultando las fosfatasas extracelulares las de mayor actividad.
A. terreus constituyó la excepción para las fosfatasas ácidas extra e intracelulares; sin embargo, esta especie presentó una mayor capacidad para producir fosfatasa alcalina intracelular (73.07 mg de p-nitrofenol mg-1 de micelio seco h-1 )
El Cuadro 2 expresa la existencia de una correlación negativa entre la actividad de las fosfatasas ácidas extracelulares e intracelulares para A. niger (Pearson p<.001), A. Terreus (Pearson p<.001), Penicillium sp. (Pearson p<.001) y P. brevicompactum (Pearson p<.01); mientras que las correlaciones entre las fosfatasas alcalinas (extracelulares e intracelulares) resultaron positivas para todos los hongos ensayados.


Al correlacionar las actividades de las fosfatasas extracelulares (ácidas y alcalinas) producidas in vitro, con aquellas presentes en el suelo de la rizosfera (Cuadro 4), sólo las actividades de las fosfatasas ácidas extracelulares producidas por A. niger y A. terreus, se correlacionaron con la actividad de las fosfatasas ácidas presentes en la rizosfera (Pearson p<.001 y p<.001, respectivamente); mientras que las actividades de las fosfatasas alcalinas extracelulares producidas in vitro y las presentes en el suelo de la rizosfera se correlacionaron negativamente entre sí (Cuadro 4).

Discusión
La mayor actividad de las fosfatasas ácidas producidas extra e intracelularmente por los hongos ensayados, corrobora los informes de Casida (1959) y Nahas et al. (1994), quienes afirman que las fosfatasas ácidas, en comparación con las fosfatasas alcalinas, son más comunes entre los hongos del suelo; lo cual podría sugerir, que en éstos, el grado de actividad de la fosfatasa ácida intracelular pudiera considerase cómo un índice de la actividad metabólica para estos organismos. Estos resultados contrastan con las comunicaciones de Ho y Zak (1979), Ezawa et al. (1995) y Joner y Johansen (2000) quienes afirman que las micorrizas poseen una mayor actividad de la fosfatasa alcalina intracelular; condición que puede estar determinada por la relación simbiótica de estos organismos con su hospedero, el cual produce fosfatasa ácida (Ridge y Rovira 1971, Shaykh y Roberts 1974, Juma y Tabatabai 1988, Tadano y Sakai 1991). Las diferencias significativas presentes entre los hongos ensayados para las actividades de las fosfatasas extra e intracelulares (ácidas y alcalinas) (Cuadro 1) indican, que estos organismos difieren en su capacidad para mineralizar el P en la rizosfera del maní.
La alta actividad de las fosfatasas extracelulares (ácidas y alcalinas), sugiere que para el momento de la determinación de las actividades de estas enzimas (después de 14 días de incubación), es posible que los niveles de P presentes en el micelio y en el medio de crecimiento, pudieran haber disminuido su concentración, provocando una mayor síntesis de fosfatasas ácidas y alcalinas extracelulares, ya que éstas pueden ser inducidas por las condiciones ambientales, en respuesta a los requerimientos de P por parte de los organismos (Dilly y Nannipieri 1988).
Aunque se ha informado que tanto el género Aspergillus como Penicillium, participan activamente en la mineralización del P orgánico del suelo (Agnihotri 1970, Tarafdar y Chonkar 1978, Alexander 1980, Tarafdar et al. 1988), los integrantes del género Penicillium (Penicillum sp., P. brevicompactum y P. chrysogenum) mostraron ser más eficientes para la síntesis de fosfatasas ácidas (extracelulares e intracelulares) y de fosfatasa alcalina extracelular (Cuadro 1). Estos resultados contrastan con los informes de Tarafdar et al. (1988), quienes afirman que entre las especies de Penicillium y Aspergillus aislados de suelos desérticos, las especies de Aspergillus eran más activas en la producción de fosfatasas. No obstante, corrobora los informes de Tarafdar (1988) en referencia a la menor capacidad de A. terreus de producir fosfatasa alcalina extracelular (Cuadro 1).
En líneas generales, las diferencias existentes con relación a la producción de fosfatasas ácidas y alcalinas (extracelulares e intracelulares), entre los diferentes hongos ensayados (Cuadro 1), están asociadas no sólo a los mecanismos de inducción y regulación enzimática (Jonner y Johansen 2000) o a las demandas nutricionales y a la disponibilidad de nutrimentos (Dighton 1991), sino también a las diferentes combinaciones e interacciones de las isoenzimas de las fosfatasas (Kroehler 1988); es decir, que estas diferencias pueden estar controladas poligenéticamente (Kropp 1990).
La magnitud de las actividades de las fosfatasas obtenidas in vitro (Cuadro 1), indica que las enzimas hifales pueden participar significativamente en la hidrólisis de los compuestos orgánicos de P en la rizosfera y facilitarle así la disponibilidad del P a las plantas, al contribuir a la actividad total de las fosfatasas en el suelo (Rao y Tarafdar 1993). La capacidad de estos hongos del suelo para emplear el P presente en la rizosfera del maní cultivado en suelo ácido, les confiere una gran ventaja adaptativa bajo estas condiciones.
La correlación negativa observada en la mayoría de los casos, entre las actividades de las fosfatasas ácidas extracelulares e intracelulares obtenidas in vitro (Cuadro 2), sugiere que su síntesis podría estar determinada por la concentración de P presente, dentro y fuera del micelio y por los mecanismos de inducción y represión (Calleja et al. 1980). Kroheler (1988) propone que un análisis del metabolismo interno y del contenido de P almacenado por los hongos estudiados, puede proveer una mejor información acerca de si la deficiencia del P en el medio o en el hongo, determinan la regulación de la producción de las actividades de las fosfatasas intracelulares y extracelulares.
La correlación positiva entre las fosfatasas alcalinas extracelulares e intracelulares (Cuadro 2), permite inferir, que la síntesis de la fosfatasa alcalina extracelular responde a las necesidades de liberación de estas fosfatasas al medio exterior, en respuesta a la disponibilidad del P.
La mayor actividad de la fosfatasa ácida, en la rizosfera del maní cultivado en suelo ácido (Cuadro 3), está relacionada, junto con la contribución de las raíces de las plantas y de la flora microbiana presente en esta zona, con la producción de esta enzima (Juma y Tabatabai 1988, Tadano y Sakai 1991, Tadano et al. 1993), actividad que resulta estimulada por la deficiencia de P característica de estos suelos (Gómez-Guiñán 1995).
La falta de correlación entre las actividades de las fosfatasas ácidas extracelulares obtenidas in vitro para la mayoría de los hongos aislados y las presentes en el suelo de la rizosfera del maní (Cuadro 4), indica que la actividad de esta enzima en el suelo está determinada por otros factores, tales como, el origen microbiano y vegetal de estas enzimas (Shaykh y Roberts 1974, Tarafdar y Classen 1988, Juma y Tabatabai 1988, Tadano et al. 1993), la adsorción y posible inactivación de las mismas por las arcillas, la materia orgánica y por la acción de las proteasas microbianas (Burns 1978, Dick et al. 1983, Nannipieri 1994, Hýsek y arapatka 1998).
La correlación negativa entre las actividades de las fosfatasas alcalinas producidas en el medio de cultivo, a excepción de Penicillium sp. y las presentes en el suelo de la rizosfera (Cuadro 4), indica que la capacidad de síntesis in vitro, de fosfatasas alcalinas extracelulares por estos organismos, no refleja la actividad de esta enzima en la rizosfera, tal vez debido a la producción de estas enzimas por otros organismos presentes en esta zona (Hýsek y arapatka 1998) y a los factores citados anteriormente (Burns 1978, Dick et al. 1983).
En general, las actividades de las fosfatasas ácidas y alcalinas producidas por los hongos en el medio de cultivo, fueron mayores a las actividades obtenidas para la zona de la rizosfera del maní con base en el peso seco (Cuadros 1 y 3). Si se toma en consideración, que la biomasa fúngica en el suelo está alrededor de 0.1-10 mg-1 g-1 de suelo seco (Joner y Johansen 2000), el potencial de estos hongos para contribuir a la mineralización del P en suelos con bajo contenido de este elemento (2.2 ppm) (Gómez-Guiñán 1995, 1998), es de significativa importancia.
Agradecimientos
Al Consejo De Investigación de la Universidad de Oriente y al FONACIT.
Resumen
El papel potencial de los hongos aislados de la rizosfera del maní, en la producción de fosfatasas ácidas y alcalinas extracelulares e intracelulares, fue evaluado in vitro. Las fosfatasas ácidas y alcalinas extracelulares, mostraron la mayor actividad y las especies de Penicillium fueron las más eficientes en su producción. Los análisis de correlación mostraron que las fosfatasas ácidas extracelulares e intracelulares producidas por A. niger, A. terreus, Penicillium sp. y P. brevicompactum se correlacionaron negativamente entre sí; mientras que las actividades de las fosfatasas alcalinas extracelulares e intracelulares, se correlacionaron positivamente. Las actividades de las fosfatasas ácidas extracelulares, producidas in vitro por la mayoría de los hongos ensayados, no se correlacionaron con la actividad de la fosfatasa ácida presente en la rizosfera del maní; sin embargo, las actividades de las fosfatasas alcalinas extracelulares producidas in vitro, se correlacionaron negativamente con las actividades de las fosfatasas alcalinas presentes en la rizosfera. La capacidad de producción de fosfatasas, por parte de los hongos aislados de la rizosfera del maní, sugiere el gran potencial de los mismos para contribuir a la mineralización del P en esta zona.
Referencias
Agnihotri, V.P. 1970. Solubilization of insoluble phosphatase by some soil fungi isolated from nursey seed-beds. Can. J. Microbiol. 16: 877-880. [ Links ]
Alexander, M. 1980. Introducción a la microbiología del suelo. Libros y Editoriales. México. 491 p. [ Links ]
Antibius, R.K., R.L. Sinsabugh & A.E. Linkins. 1992. Phosphatase activities and phosphorus uptake from inositol phosphate by ectomycorrhizal fungi. Can. J. Bot. 70:794-800. [ Links ]
Baligar, V.C., R.J. Wright & M.D. Smedly. 1988. Acid phosphatase activity in soil of the Appalachian region. Soil Sci. Soc. Amer. J. 52: 1612-1616. [ Links ]
Beever, R.E. & D.J.W. Burns. 1980. Phosphorus uptake, storage and utilization by fungi. Adv. Bot. Res. 8: 127-219. [ Links ]
Burns, R.G. 1978. Enzyme activity in soil: Some theoretical and practical considerations, pp. 295-340. In R.G. Burns (ed.). Soil Enzyme. Academic. Nueva York. [ Links ]
Calleja, M., D. Mousain, B. Lecouvreur & J. DAuzac. 1980. Influence de la carence phosphatée sur les activités phosphatases acides de trois champignons mycorhiziens: Hebeloma edurum Metrod., Suillus granulatus (L. et Fr.) O. Kuntze et Pisolithus tinctorius (pers.) Coker et Couch. Physiol. Veg. 18: 489-504. [ Links ]
Casida, L.E. 1959. Phosphatase activity of some common soil fungi. Soil Sci. 87: 305-310. [ Links ]
Curl, E.A. & B. Truelove. 1986. The rhizosphere. Springer- Verlag. Berlin. 288 p. [ Links ]
Dick ,W.A., N.G. Juma & M.A. Tabatabai. 1983. Effects of soils on acid phosphatase and inorganic pyrophosphatase of corn roots. Soil Sci. 136: 19-25. [ Links ]
Dighton, J. 1983. Phosphatase production by mycorrhizal fungi. Plant Soil. 71: 455-462. [ Links ]
Dighton, J. 1991. Acquisition of nutrients from organic sources by mycorrhizal autotrophics plants. Experientia 47: 362-369. [ Links ]
Dilly, O. & P. Nannipieri. 1998. Intracellular and extracellular enzyme activity in soil with reference to elemental cycling. Z. Pflanzenernahr. Bodenk. 161: 243-248. [ Links ]
Dinkelaker, B. & H. Marschner. 1992. In vivo demostration of acid phosphatase activity in the rhizosphere of soil-grown plants. Plant Soil 144: 199-205. [ Links ]
Ezawa, T., M. Saito & T. Yoshida. 1995. Comparison of phosphatase localization in the intraradical hyphae of arbuscular mycorrhizal fungi Glomus spp. and Gigaspora spp. Plant Soil 176: 67-63. [ Links ]
Gómez-Guiñán, Y. 1995. Efecto del abono orgánico tierra de filtro (TF) sobre las actividades de las fosfatasas y la disponibilidad del fósforo en la rizosfera del maní (Arachis hypogaea L.) cultivado en suelo ácido. Trabajo de Ascenso para optar a la categoría de Profesor Titular. Puerto La Cruz, Estado Anzoátegui. [ Links ]
Gómez-Guiñán, Y. & I. Nageswara. 1996. Efecto de la rizosfera del maní (Papilonaceae) sobre la flora micótica y la actividad de las fosfatasas. Carib. J. Sci. 32: 214-220. [ Links ]
Gómez-Guiñán, Y. 1998. IV Congreso Interamericano sobre el Medio Ambiente. Universidad Simón Bolivar. Caracas, Distrito Federal. pp. 253-257. [ Links ]
Hýsek, J. & B. arapatka. 1998. Relationship between phosphatase active bacteria and phosphatase activities in forest soils. Biol. Fertil. Soils 26: 112-115. [ Links ]
Ho, I. 1988. A simple method for assessing acid phosphatase activity of ectomycorrhizal fungi. Trans. Br. Mycol. Soc. 91: 346- 347. [ Links ]
Ho, I. & B. Zak. 1979. Acid Phosphatase activity of six ectomicorrhizal fungi. Can. J. Bot. 57: 1203-1205. [ Links ]
Joner, E.J. & A. Johansen. 2000. Phosphatase activity of external hyphae of two arbuscular mycorrhizal fungi. Mycol. Res. 104: 81-86. [ Links ]
Juma, N.G. & M.A. Tabatabai. 1988. Phosphatase activity in corn and soybean roots: Conditions for assay and effects of metals. Plant Soil 107: 39-47. [ Links ]
Kroehler, C. 1988. The effects of organic and inorganic phosphorus concentration on the acid phosphatase activity of ectomycorrhizal fungi. Can. J. Bot. 66: 750-756. [ Links ]
Kropp, B.R. 1990. Variation in acid phosphatase activity among progeny from controlled crosses in the ectomycorrhizal fungus Lacaria bicolor. Can. J. Bot. 68: 864-866. [ Links ]
Kyochi, O. & S. Yoshida. 1988. Phosphomonoesterase activity in cultivated soils amended with different levels of organic materials. Soil Sci. Plant Nutr. 34:239-296. [ Links ]
Nahas, E., J.F. Centurion & L.C. Assis. 1994. Microrganismos solubilizadores de fosfato e produtores de fosfatases de vários solos. Rev. Bras. Cien. Solo 18: 43-48. [ Links ]
Nannipieri, P. 1994. The potential use of soil enzyme as indicators for productivity, sustainability and pollution, pp. 238-244. In C.E. Pankhurst, B.M. Doube, V.V.S.R. Gupta and P.R. Grace (eds.). Soil biota management in sustainable farming systems. CSIRO. Australia. [ Links ]
Rao, A.V. & J.C. Tarafdar. 1993. Role of VAM fungi in nutrient- uptake and growth of clusterbean in an arid soil. Arid Soil Res. Rehab. 7: 275- 280. [ Links ]
Ridge, E.H. & A.D. Rovira. 1971. Phosphatase activity of intact young wheat roots under sterile and non-sterile conditions. New Phytol. 70: 1017-1026. [ Links ]
Shaykh, M.N. & L.W. Roberts. 1974. A histochemical study of phosphatase in root apical meristems. Ann. Bot. 38: 165-174. [ Links ]
Tabatabai, M.A. & J.M. Bremner. 1969. Use of p-nitrophenylphosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1: 301-307. [ Links ]
Tadano, T. & H. Sakai. 1991. Secretion of acid phosphatase by the roots of several crop species under phosphorus- deficient conditions. Soil Sci. Plant. Nutr. 37: 129-140. [ Links ]
Tadano, T., K. Ozawa, H. Sakai & H. Matsui. 1993. Secretion of acid phosphatase by root of crop plants under phosphorus deficient conditions and some properties of the enzyme secreted by Lupin roots. Plant Soil. 156: 95-98. [ Links ]
Tarafdar, J.C., A.V. Rao & K. Bala. 1988. Production of phosphatase by fungi isolated from desert soils. Folia Microbiol. 33: 453-457. [ Links ]
Tarafdar, J.C. & N. Classen. 1988. Organic phosphorus compounds as a phosphorus source for higher plants through the activity of phosphatase produced by plant roots and microorganisms. Biol. Fertil. Soil. 5: 308-312. [ Links ]
Tarafdar, J.C. & P.B. Chhonkar. 1978. Status of phosphatase in the root soil interface of leguminous and non-leguminous crops. Z. Pflanzenner Narchr. Bodenkd. 1941: 347-351. [ Links ]

