











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.51 no.3-4 San José Set. 2003
ontogenetic variation (Serpentes, Elapidae: Leptomicrurus)
William W. Lamar
College of Sciences, The University of Texas at Tyler. Tyler, Texas 75799, USA.
Received 12-IV-2003. Corrected 29-VIII-2003. Accepted 01-IX-2003.
Abstract
Leptomicrurus renjifoi is described from tropical semi-deciduous forest of the eastern Colombian llanos. It is one of the smallest species in the genus, is most similar to L. scutiventris, and it may be distinguished from known congeners by a combination of color, pattern, and scale characters. Evidence for the recognition of Leptomicrurus is convincing, although its members were recently thought to comprise a closely related assemblage within Micrurus. A supposedly aberrant specimen of L. scutiventris may indicate clinal or ontogenetic variation in pattern.
Key words: Reptilia, Squamata, Serpentes, Elapidae, Leptomicrurus renjifoi, Leptomicrurus scutiventris, Leptomicrurus, Micrurus, clinal variation, Colombia, South America.
The genus Leptomicrurus was erected by Schmidt (1937) to accomodate two species of exceptionally slender, short-tailed coralsnakes, L. collaris (Schlegel) and L. narduccii (Jan). These are distinguished from members of the genus Micrurus on the basis of having the mental scale in contact posteriorly with the anterior chinshields, and possessing a body pattern not including rings or bands. Leptomicrurus was interpreted variously with regard to L. collaris (Schmidt 1937, Brongersma 1966, 1967), and Schmidts genus was relegated to a junior synonym of Micrurus when specimens of Leptomicrurus with some complete rings and specimens of Micrurus with anterior chinshields in contact with the mental were discovered (Romano 1972).
Leptomicrurus has been revalidated and its three species redefined (Roze and Bernal-Carlo 1988). Micrurus karlschmidti Romano was synonymized with Elaps scutiventris Cope, both names becoming subjective junior synonyms of Leptomicrurus scutiventris (Cope) (Roze and Bernal-Carlo 1988). The reasons for resurrecting Leptomicrurus are those of Schmidt coupled with certain hemipenial and osteological characters (Roze and Bernal-Carlo 1988). Roze and Bernal-Carlo also pointed out that mental-chinshield contact in Micrurus is rare, and rings when present on specimens of Leptomicrurus are anomalous, uncommon, and poorly defined.
These data support an hypothesis of close relationship among the three species assigned to Leptomicrurus, and they provide sufficient evidence for generic distinction. Roze and Bernal-Carlo (1988) was not followed by Campbell and Lamar (1989) because it was published after they had gone to press (contra Harvey et al. 2003). However, these authors did note that Micrurus (= Leptomicrurus) narduccii might represent a composite of more than one species and that Micrurus (= Leptomicrurus) karlschmidti possessed a lower range of ventral scales. A detailed morphological description is provided herein because none exists for any member of the genus.
Although the herpetofauna of the llanos of Colombia and Venezuela is thought to be relatively well known, portions of this region remain poorly inventoried. Given the difficulty of encountering fossorial and leaf-litter inhabitants in gallery forests, the discovery of two unidentified specimens of Leptomicrurus housed in the University of Texas at Arlington Collection of Vertebrates was not surprising. They represent a distinctive species of the genus Leptomicrurus, accordingly described as
Leptomicrurus renjifoi sp. nov.
Figs. 1 and 2

Holotype: Collection of Vertebrates, The University of Texas at Arlington UTA R-3490, a gravid adult female (Fig. 1), from the south side of the Río Tomo, near its juncture with the Río Orinoco, 97 km SSW of Puerto Carreño, Comisaría del Vichada, Colombia, 115 m, collected by Flavio Barney in August, 1969.
Paratype: UTA R-3491, a topotypic subadult female, same locality data as holotype; 156 m; collected in November, 1969.
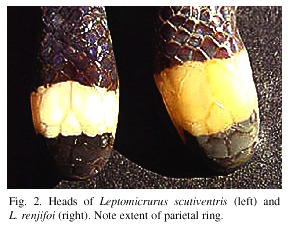
Definition and diagnosis: This bicolored species differs from all congeners by possessing a pattern of black rings separated by equally long (or longer) pale pinkish orange rings, a pale pinkish orange parietal ring, a black nuchal ring that passes posterior to the parietal tips and the seventh supralabial, pale tail rings that are neither red nor orange, and 243 or fewer ventrals in females. For further differences that distinguish L. renjifoi from its congeners, see Remarks and the Key to Leptomicrurus.
Description of holotype: An adult female, 427 mm in total length; tail length 19.7 mm (4.6% of total); head length 10.0 mm from front face of rostral to posterior end of mandible; head width 5.9 mm at broadest point (posterior edge of parietal ring); head scarcely distinct from neck; snout moderately truncated in dorsal view; eye small, snout about 3.0 times as long as horizontal distance across eye; pupil subelliptical; rostral about 1.2 times broader than high; internasals 1.2 times wider than long, laterally contacting anterior and posterior nasals; prefrontals moderate, slightly wider than long, laterally contacting posterior nasal and preocular; median prefrontal suture 0.5 times as long as frontal; frontal about 1.4 times longer than wide; parietals about 1.6 times longer than wide, median suture about 91.4% of frontal length; nostrils large, located in anterior portion of posterior nasals, dorsally oriented; one preocular; two postoculars; temporals 1+1, about 2.0 times longer than wide, separating supralabials 5, 6, and 7 from parietal; supralabials 7/7, 1 and 2 contacting anterior nasal, 2 and 3 contacting posterior nasal, 3 contacting preocular, 3 and 4 entering orbit, 4 and 5 contacting lower postocular, 5 and 6 contacting anterior temporal, 6 (largest) and 7 contacting posterior temporal; mental 1.2 times broader than long, posteriorly in contact with anterior chinshields; anterior chinshields well developed, about twice as long as wide, median suture equal to about half of scale length; posterior chinshields distinct from gulars, about twice as long as wide, median suture equal to about 3/4 of scale length; posterior chinshields separated from first ventral by 4 gulars plus 3 preventrals; infralabials 7/7, 1-4 contacting anterior chinshields, fourth largest and contacting posterior chinshields; dorsal scales disposed in 15 smooth rows throughout length of body; dorsal scales in 6 rows at level of tenth subcaudal; no apical pits apparent; ventrals 238 (following Dowling 1951); anal divided; subcaudals 16, paired; cap scale about 1.5 times as long as preceding subcaudal; tail short, thick, with tip acutely rounded.
In life the color pattern consisted of alternating black and pale pinkish orange rings on the head, body, and tail, a condition unique in the genus. In preservative (ethanol after formaldehyde) the snout is black, this color extending posteriorly and including the rostral, anterior and posterior nasals, supralabials 1 and 2 (except for the posterior lower corner of supralabial 2), the dorsal half of supralabial 3, a tiny portion of supralabial 4 (just below eye), the internasals, prefrontals, preoculars, the anterior half of the supraoculars, and about the anterior one fifth of the frontal. Black pigment is present ventrally on the anterior portion of the mental. The pale yellowish white parietal ring comprises the rest of the head and lower jaw and is immaculate save for slight duskiness along the frontal and interparietal sutures.
The black nuchal ring, 8 middorsal scales long, begins just posterior to the parietal scale tips and passes just posterior to the supralabials. Owing to a posterior ventral invasion of the parietal ring color, the anterior portion of the black nuchal ring is reduced laterally to a length of 5 1/2 dorsal scales and ventrally to a length of 4 1/2 ventral scales. Including the nuchal, there are 28 + 1/2 black body rings, the last one being situated over the vent and base of the tail. Three of the black body rings are reduced to dorsal blotches. There are 1/2 + 1 black tail rings and the last ring is incomplete ventrally. Exclusive of the parietal ring, there are 28 pale yellowish white body rings; these are immaculate ventrally and laterally, slightly dusky along the scale sutures dorsally, and contain an occasional black scale mid dorsally. Including the tail tip, there are two pale yellowish white tail rings. The iris is black.
Variation: The type series consists of two females. The holotype has a total length of 427 mm. The paratype is 182 mm overall, with a tail length of 8.8 mm, comprising 4.8% of the total length. The frontal width is 0.63-0.78 (=0.70) of the frontal length; the average cephalic index (width/length X 100) is 42.6. The eye is contained 2.72-3.07 (=2.8) times in the length of the snout. There are 238-243 (=240.5) ventrals and 16-19 (=17.5) subcaudals. The paratype possesses only one left postocular and only 18 subcaudals on the left side of the tail.
Black pigment on the snout of the paratype is practically identical in extent to that of the holotype; on the lower jaw it includes the entire mental and most of the adjacent first infralabial scales. The parietal ring is immaculate, pale yellowish white, with no darker dusting along the supracephalic scale sutures.
The paratype possesses 27 + 1/2 black body rings (including the nuchal), two of which are reduced to dorsal bands; there are 1/2 + 1 black tail rings. These black rings alternate with 27 pale yellowish white rings on the body plus two on the tail. The black rings are 2-7 (usually 4-4 1/2) mid body dorsals and 1 1/2-4 (usually 3-3 1/2) ventrals in length. The lower dorsal counts are because of a dorsal constriction on some black rings. The pale rings are 2 1/2-8 (usually 4-5) mid body dorsals and 4-10 (usually 4 1/2-6) ventrals long. The higher dorsal counts are adjacent to constricted black rings.
Overall the paratype closely resembles the holotype in color, but the pale rings are more immaculate dorsally. Ventrally the pale rings are immaculate with the exception of two that are suffused with a soft gray color. In life all pale rings were pinkish orange, like those of the holotype.
Etymology: The patronym renjifoi honors Juan Manuel Renjifo, a Colombian herpetologist who has contributed to our knowledge of venomous snakes of that region, and whose photographs of wildlife rank among the worlds finest. The gender is masculine.
Distribution and habitat: This species is known only from the type locality (but see Remarks for further discussion). The habitat is distinctive as compared with the mesophytic forests inhabited by the most similar congener, Leptomicrurus scutiventris. The eastern llanos of Colombia is a complex of savannas and forests that have been divided collectively into a dozen types (Vincelli 1981). Annual temperature averages 26.5 o C, and rainfall is strongly seasonal, with about 2100 to 2394 mm falling from April to November (Defler 1979, Anonymous 1976). Drought-like conditions are prevalent during the dry season (Brunnschweiler 1972).
The type locality is in non-inundated, semi-deciduous gallery (riparian) forest (Beard 1955) with an average canopy height of 20-22 m. Dominant vegetation includes Jessenia polycarpa, Calophyllum lucidum, Amaioua guianensis, Enterolobium schomburgkii, Terminalia amazonia, Guatteria gracilipes, Miconia holosericea, Parkia pendula, Licania subarachnophylla, Hymenaea courbaril, and Euterpe precatoria (Vincelli 1981). The forest consists of 100-110 ha surrounding a 100 m high granitic outcropping of the Guyana Shield (Defler 1979), and bordered by a Stipa/sedge savanna (Defler 1983, Vincelli 1981). The forest floor is gravelly soil covered with leaf litter.
Remarks: Leptomicrurus narduccii is of doubtful sympatry with L. renjifoi, and L. collaris is restricted to northeastern South America. The range of L. scutiventris closely approaches that of L. renjifoi from the south (Roze and Bernal-Carlo 1988, Roze 1996).
Rarely, specimens of L. narduccii and L. scutiventris may be found with a few anterior body rings but invariably these are short (1 dorsal scale in length), indistinct projections from the ventral blotches. The pale cephalic ring in L. collaris and L. scutiventris fails to cover the posterior portion of the parietals, while that of L. narduccii fails to cover the anterior portion of the parietals. The pale cephalic ring of L. renjifoi completely encompasses the parietals. Unlike its three congeners, there is no red or orange pigment in either the pale cephalic or the caudal rings of L. renjifoi. Hoge and Romano (1966) described Leptomicrurus schmidti from Tapurucuara, Uaupés, Amazonas, Brazil. These authors noted a tendency of the anterior ventral blotches to be joined by short dorsal bands to form rings in this species. This taxon was renamed Micrurus karlschmidti by Romano (1972) before it was synonymized with L. scutiventris by Roze and Bernal-Carlo (1988), who suggested that if the population of L. scutiventris from northwestern Brazil and adjacent Colombia should prove to be distinctive, the name L. schmidti would be available.
A tiny specimen of L. scutiventris in the Naturhistorisches Museum of Vienna (NMW 9173) from São Pedro de Olivença, Amazonas, Brazil, is unusual in possessing short (1 dorsal scale length), indistinct, pale rings connecting the majority of ventral blotches dorsally. It is a female with 270 ventrals and a total length of 155 mm (Roze and Bernal-Carlo 1988). As suggested by Roze and Bernal-Carlo (1988), it might indicate a distinctive population of L. scutiventris. Owing to the similar lepidosis shared by L. scutiventris and L. renjifoi, it might also be argued that the patterns of the latter and NMW 9173 are the product of north-south clinal variation in L. scutiventris. However, two specimens (ICN 03-04) from Amanavén, at the mouth of the Río Guaviare, Vichada, Colombia, are typical of L. scutiventris (Roze and Bernal-Carlo 1988, Roze 1996). Amanavén lies in mesophytic forest a scant 128 km S of the type locality of L. renjifoi, and well north of the São Pedro de Olivença site.
Whether the tendency to form rings is prevalent in this population, and how that might relate to L. renjifoi is debatable. Nothing is known about reproduction in Leptomicrurus, and neonates, with the exception of NMW 9173, have not been observed. Ontogenetic pattern change like that observed for some populations of Oxyrhopus petolarius involving progressive melanism beginning posteriorly might account for the presence of anterior rings in some specimens of L. narduccii and L. scutiventris. This would also explain the ring condition in specimen NMW 9173.
Behavior: Both specimens of L. renjifoi thrashed from side to side and raised and curled the tail when molested. The holotype was gravid and contains two eggs.

Acknowledgements
Specimens and other courtesies were provided by: Jonathan A. Campbell, The University of Texas at Arlington (UTA); James R. Dixon and Kathryn Vaughn, Texas A & M University (TCWC); William E. Duellman and John R. Simmons, the University of Kansas (KU); Darrel R. Frost and Linda Ford, the American Museum of Natural History (AMNH); John D. Lynch, the late Federico Medem, and Olga Victoria Castaño, the Universidad Nacional de Colombia (ICN); William B. Montgomery and Juan Silva-Haad. The late Flavio Barney, René Pérez, and Miguel Mesía helped with field work. Additional information was provided by Thomas Defler, the late Flavio Barney, Janis A. Roze, and Paul Vincelli. Louis W. Porras, Mahmood Sasa, and Alejandro Solórzano made comments about the manuscript. Randy Mallory assisted with photograph.
Resumen
Leptomicrurus renjifoi es una nueva especie de serpiente elápida del bosque tropical semi-decíduo de los llanos orientales de Colombia. Es una de las especies más pequeñas del género, se asemeja más a L. scutiventris, y se distingue de sus congéneres por una combinación de color, patrón, y carácteres de escamación. Existe suficiente evidencia morfológica para reconocer Leptomicrurus como género independiente, aunque hasta hace poco sus miembros fueron considerados como un grupo compacto dentro del género Micrurus.
References
Anonymous. 1976. Distribución de la precipitación media anual en Colombia (mapa preliminar). Inst. Colombiano de Hidrología, Meteorología, y Adecuación de Tierras (HIMAT), Santa Fe de Bogotá. [ Links ]
Beard, J.S. 1955. The classification of tropical American vegetation types. Ecology 36: 89-100. [ Links ]
Brongersma, L.D. 1966. Note on Leptomicrurus collaris (Schlegel) (Reptilia, Serpentes). Zool. Meded. Leiden 41: 243-254. [ Links ]
Brongersma, L.D. 1967 [dated 1966]. The poisonous snakes of Surinam. Mem. Inst. Butantan 33: 73-79. [ Links ]
Brunnschweiler, D. 1972. The Llanos: frontier of Colombia. Latin American Studies Center, Monograph 9, Michigan St. Univ., 71 pp. [ Links ]
Campbell, J.A. & W.W. Lamar. 1989. The venomous reptiles of Latin America. Cornell Univ. Press, Ithaca, New York, 425 pp. [ Links ]
Defler, T.R. 1979. On the ecology and behavior of Cebus albifrons in eastern Colombia: I. Ecology. Primates 20: 475-490. [ Links ]
Defler, T.R. 1983. A remote park in Colombia. Oryx, Vol. XVII (January 1983): 15-17. [ Links ]
Dowling, H.G. 1951. Aproposed standard system of counting ventrals in snakes. British J. Herpetol. 1: 97-99. [ Links ]
Harvey, M.B., J. Aparicio E. & L. González A. 2003. Revision of the venomous snakes of Bolivia: Part I. The coralsnakes (Elapidae: Micrurus). Carnegie Museum Publications in Zoology. [ Links ]
Hoge, A.R. & S.A.R.W.D.L. Romano. 1966 [dated 1965]. Leptomicrurus in Brasil (Serpentes - Elapidae). Mem. Inst. Butantan 32: 1-8. [ Links ]
Romano, S.A.R.W.D.L. 1972 [dated 1971]. Notes on Leptomicrurus Schmidt (Serpentes, Elapidae). Mem. Inst. Butantan 35: 111-115. [ Links ]
Roze, J.A. 1996. Coral snakes of the Americas. Biology, identification, and venoms. Kreiger Publishing, Malabar, Florida, 328 p. [ Links ]
Roze, J.A. & A. Bernal-Carlo. 1988 [dated 1987]. Las serpientes corales venenosas del género Leptomicrurus (Serpentes, Elapidae) de Suramérica con descripción de una nueva subespecie. Boll. Mus. Reg. Sci. Nat. Torino. 5: 573-608. [ Links ]
Schmidt, K.P. 1937. The history of Elaps collaris Schlegel, 1837-1937. Field Mus. Nat. Hist. Publ., Zool. Ser. 20: 361-364. [ Links ]
Vincelli, P.C. 1981. Estudio de la vegetación del Territorio Faunistico "El Tuparro." Cespedesia 10(37-38): 7-51 [ Links ]
Specimens examined:
Leptomicrurus collaris – SURINAME: Brownsberg, 675 m, AMNH 131309.
Leptomicrurus narduccii – COLOMBIA: Putumayo: Puesto de Bombeo Guamuez, 100 m, KU 140414; ECUADOR: Napo: Misahualli (personal collection); Lago Agrio, 330 m, KU 126057.
Leptomicrurus scutiventris – COLOMBIA: Amazonas: Leticia, 80 m, (J. Silva-Haad collection); Vichada: Amanavén, mouth of Río Guaviare, 140m, ICN 03- 04; PERU: Loreto: Padre Cocha, Río Nanay, 170 m (personal collection); San Andrés, Río Momón, 170 m (personal collection).

