











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.51 no.3-4 San José sep. 2003
Phaseolus lunatus (Fabaceae) del Valle Central de Costa Rica
Elida M. Vargas, 1,3 Emilio Castro, 1 Gabriel Macaya 2 & Oscar J. Rocha 1*
1 Escuela de Biología, Universidad de Costa Rica. Ciudad Universitaria "Rodrigo Facio", San José, Costa Rica.
2 Centro de Investigación en Biología Celular y Molecular, Universidad de Costa Rica. Ciudad Universitaria "Rodrigo Facio", San José, Costa Rica,
3 Departamento de Ciencias Naturales (Sección de Biología), Universidad de Costa Rica. Ciudad Universitaria "Carlos Monge", Sede de Occidente, San Ramón, Costa Rica.
Recibido 06-IX-2001. Corregido 19-VI-2002. Aceptado 08-VIII-2002.
Abstract
We studied the morfological diversity in fruits and seeds in 38 wild populations of Phaseolus lunatus var. lunatus (lima beans) in the central valley of Costa Rica. In order to do so, measured the lenght and width of the fruits and the lenght, width and thickness of seeds. We also calculated the ratio between these traits and determined the weight of 100 seeds. In general, we found significant variation between populations for all variables. When we grouped the 38 populations into eight geographical regions within the study area, we found significant differences between regions. However, the levels of variation between populations within geographical regions was larger than that found between geographical regions. These findings suggested that there is no clear relationship between these variables and the geographical grouping established in this study. The implications of these findings for the establishment of strategies for in situ conservation of wild populations of lima beans are discussed.
Key words: gene pool, genetic diversity, in situ conservation, Leguminosae, Lima beans, Mesoamerica, morphological variation.
La distribución de la variación genética dentro y entre distintas áreas geográficas es de gran interés para la conservación de la diversidad biológica (Hamrick y Godt 1989, Frankel et al. 1995, Guarino et al. 1995, Prance 1997). Esto es especialmente importante para especies de interés comercial, donde la diversidad genética constituye una fuente de genes que pueden ser incorporados en programas de mejoramiento genético tanto en plantas como en animales (Prance 1997). Este es el caso de los parientes silvestres de las plantas cultivadas, que forman parte del acervo genético primario de los cultivos, y cuyos genes pueden ser fácilmente incorporados a variedades mejoradas (Ford-Lloyd y Jackson 1986).
La diversidad de los caracteres morfológicos constituye la espina dorsal de los estudios sobre la diversidad genética en cualquiera de sus niveles (Chandran y Pandya 2000). En el caso de las especies cultivadas, este tipo de información ha sido ampliamente utilizada para discriminar entre materiales procedentes de distintas regiones geográficas y ha permitido un manejo más eficiente de los bancos de germoplasma en los grandes centros internacionales (Ariyo 1993, Polignano et al. 1993, Annicchiarico y Pecetti 1994, Hoc et al. 2003a, b). Estos trabajos también han permitido establecer relaciones genéticas entre cultivares y razas locales de diferente origen y además, han permitido relacionar la diversidad observada con diferentes regiones geográficas y otros elementos del ambiente físico.
Se han realizado diversos estudios con el objetivo de medir la variación de las características morfológicas en poblaciones silvestres de plantas cultivadas. Letschert y Frese (1993) evaluaron la variación morfológica en condiciones controladas de 35 introducciones de remolacha silvestre (Beta vulgaris) procedentes de Sicilia. Ellos midieron 15 caracteres diferentes dentro de los cuales se incluyen el patrón de crecimiento de la planta y diversas características del tallo, las hojas y las flores y encontraron que existe poca variación en todos ellos. Sin embargo, los autores mostraron que existe un patrón de distribución geográfica que permite distinguir entre los grupos de la costa y los del interior de la isla. De manera similar, Clements y Cowling (1994) evaluaron la variación morfológica en 157 poblaciones silvestres de Lupinus angustifolius de la región egea con relación al centro de origen geográfico de esta especie. Estos autores evaluaron 22 caracteres diferentes y demostraron una amplia variación morfológica tanto entre como dentro de los sitios de recolección y entre localidades de Grecia. Sus resultados indican que la variación de varios de los caracteres examinados está relacionada con diferencias climáticas entre los sitios estudiados.
Otros trabajos realizados con especies no domesticadas revelan que la distribución de los caracteres morfológicos sigue patrones que se puede asociar a regiones geográficas (Nevo et al. 1982, 1983, 1988, 1991). Por ejemplo, Allen et al. (1996) analizaron la variación genética y morfológica en 12 poblaciones del lirio Erythronium montanun, representadas en diferentes regiones geográficas (Oregon, Washington y Columbia Británica) del área oeste de América del Norte. Sus resultados, muestran diferencias entre regiones y entre poblaciones dentro de cada región para algunos de los caracteres estudiados, pero los autores no pudieron identificar ningún patrón geográfico de variación que estuviera bien definido. Por su parte, Wulff (1991) en un estudio realizado con tres especies del género Plantago, encontró una relación entre el sistema de cruzamiento y el nivel de variación de los caracteres morfológicos. Además, sus resultados indican que el grado de diferenciación genética entre las poblaciones es mayor para caracteres morfológicos que para marcadores isoenzimáticos. Esto es, la diferenciación se expresa más claramente en los caracteres morfológicos que en los marcadores moleculares, dado que los primeros presentan una gran plasticidad fenotípica que responde a las diferencias del ambiente.
Uno de los principales criterios que se han utilizado para explicar el origen y la diversidad del frijol lima es el estudio de la variación de las características morfológicas en semillas. Se ha demostrado que el morfotipo "big lima" que presenta semillas relativamente grandes, pertenece al reservorio genético andino, mientras que los morfotipos "potato" (con semillas pequeñas) y "sieva" (tamaño intermedio) pertenecen al reservorio de Mesoamérica (Baudoin 1988, Debouck et al. 1989, Esquivel et al. 1990, Lioi et al. 1991). Esquivel et al. (1990) estudiaron el origen y la distribución del frijol lima en Cuba utilizando diferentes caracteres morfológicos de las semillas en 173 poblaciones. Con base en la variación observada para 11 caracteres diferentes, estos autores lograron clasificar a estos frijoles en siete grandes grupos: silvestre, maleza, potato, potato- sieva, sieva, sieva-big lima y big lima. Si bien es cierto que ya se tenía información acerca de los principales morfotipos como sus formas intermedias en esta especie, es importante hacer notar que la variación en las semillas fue suficiente para distinguir entre los diferentes grupos. A pesar de ello, no se ha evaluado el nivel de variabilidad morfológica que existe en poblaciones silvestres de frijol lima que crecen dentro de una misma región.
En este estudio se analiza la distribución de la variación de diversas características morfológicas de los frutos y semillas procedentes de diferentes poblaciones silvestres de P. lunatus que crecen en el Valle Central de Costa Rica. Además, se analiza la distribución de esta variación en función de las ocho regiones geográficas definidas en este estudio.
Materiales y metodos
Area de estudio. El estudio se realizó en las regiones altas del Valle Central y la vertiente del Río Grande de Tárcoles, Costa Rica. Comprende diferentes localidades de las provincias de Cartago, Heredia, Alajuela y San José. Estas localidades forman parte del Valle Central donde crecen naturalmente poblaciones silvestres de P. lunatus. En total, el área de estudio tiene una extensión de aproximadamente 2100 km2 y es la región donde se concentra el mayor porcentaje de la población humana del país (Fig. 1).
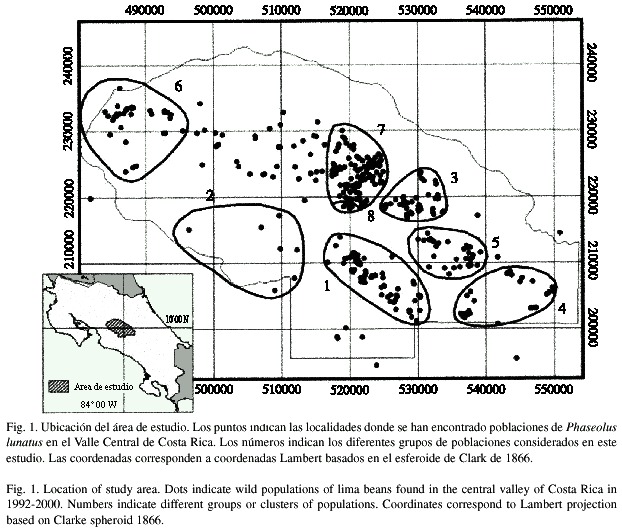
Poblaciones escogidas. Entre 1992 y 1993 se realizó un censo en el área de estudio y se localizaron unas 450 poblaciones silvestres de frijol lima (Rocha et al. 1997). Para efectos del presente estudio, se define una población como un grupo de plantas separadas al menos por 500 m de otras plantas de la misma especie. Del total de las 450 poblaciones ubicadas inicialmente, se seleccionó una muestra de 38 que fueran representativas del área en estudio. En el Cuadro 1 se presenta la ubicación de estas poblaciones. Tomando en cuenta las condiciones climatológicas y la distribución geográfica, las 38 poblaciones se agruparon en ocho regiones, cada una de las cuales está representada por al menos dos poblaciones diferentes (Fig. 1).
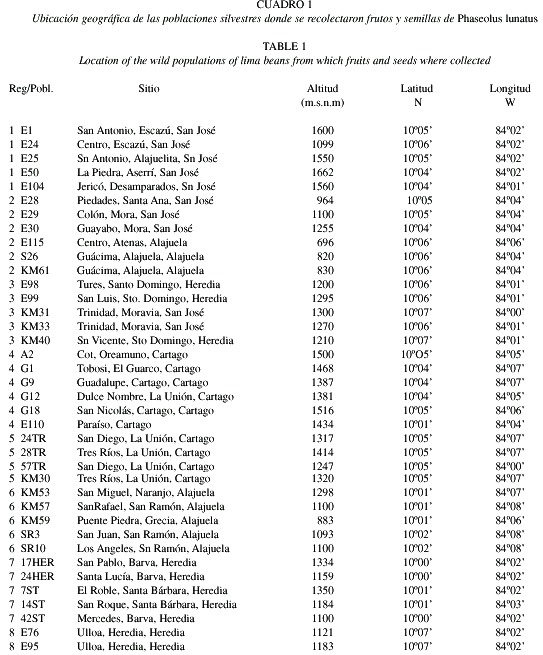
Caracteristicas morfológicas. Para cada población silvestre seleccionada, se hizo una descripción detallada de su ubicación y se colectó el mayor número posible de vainas maduras. Las semillas se separaron de las vainas el mismo día de recolección o a más tardar el día siguiente. Antes de separar las semillas se midió el largo y ancho de 30 vainas por población. Las vainas se seleccionaron al azar. Por cada vaina que se midió, se seleccionó una semilla al azar. A cada semilla se le midió el largo, el ancho y el grosor. En las mediciones de las diferentes características morfológicas de las vainas y semillas se utilizó un calibrador digital, marca "MAX-CAL".
Una vez realizadas todas las mediciones, se separaron las semillas de las vainas y se colocaron en cajas petri destapadas, las que se guardaron dentro de cajas plásticas tapadas a las que previamente se les adicionó aproximadamente un kilogramo de sílica gel. En estas condiciones las semillas se secaron durante 30 días a temperatura ambiente reemplazando la sílica gel cuando fuera necesario. Después de este tratamiento las semillas secas fueron trasladadas a bolsas plásticas y se sellaron herméticamente para evitar la entrada de agua. Estas bolsas se almacenaron en cajas plásticas con tapa a 4 Cº. Por cada población se seleccionaron al azar, dos o tres muestras de 100 semillas secas cada una y se pesaron en una balanza analítica.
Resultados
Los resultados de este estudio indican niveles de variación importantes en torno a los valores promedio las vainas y semillas de P. lunatus en el Valle Central de Costa Rica (Cuadro 2). El estudio de la distribución de la variación de todas las variables morfológicas se realizó mediante un análisis de variancia multivariado (Proc. GLM con la opción MANOVA del SAS, Anónimo 1985). Los resultados del mismo se presentan en el Cuadro 3. En este análisis, se estudió el efecto de la identidad de la población y la región geográfica en que la población se encuentra ubicada sobre las variables morfológicas medidas. La distribución de las poblaciones en ocho regiones, se hizo tomando en cuenta diversos criterios tales como la ubicación geográfica, el tipo de suelos y la altitud donde crecen naturalmente las poblaciones silvestres de frijol lima. También se tomaron en cuenta las condiciones del clima (temperatura y precipitación) para cada zona, lo mismo que las características fenológicas de las poblaciones.

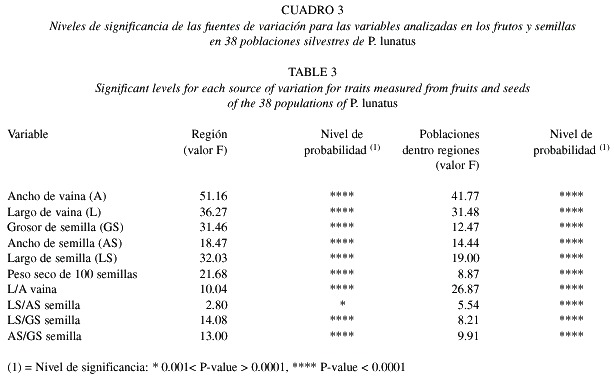
Variacion de las caracteristicas morfológicas. El análisis de diferentes variables físicas en las vainas y semillas de frijol lima demuestra que existe una amplia variación morfológica no sólo entre y dentro de las poblaciones estudiadas, sino también entre regiones. Los resultados del análisis de variancia multivariado revelan que existen diferencias significativas entre las distintas regiones origen de las poblaciones (Región) y entre las poblaciones de una misma región (Poblaciones (Región)) en todas las variables estudiadas (Cuadro 3).
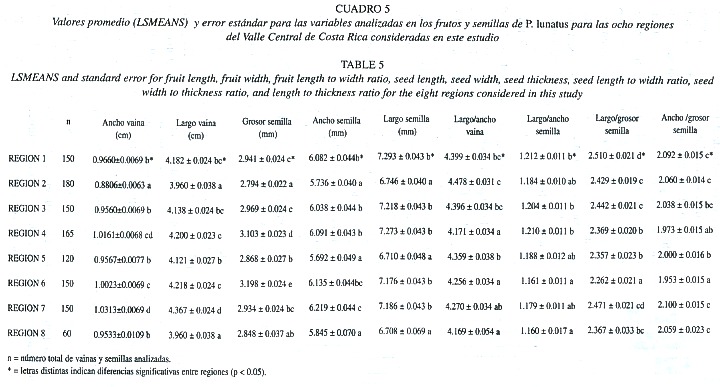
En las vainas, el valor promedio para el ancho varió entre 0.750 ± 0.015 cm (en la región 2) y 1.167 ± 0.015 cm (en la región 7) (Cuadro 4). A pesar de los valores extremos que presentan estas dos poblaciones, en aproximadamente el 74% de las poblaciones el ancho de las vainas varió entre 0.907 y 1.167 cm. Si se comparan estos resultados con el promedio de 0.97 ± 0.13 obtenido al agrupar todas las poblaciones (Cuadro 2), se observa que en promedio, el ancho de las vainas en el frijol lima es de 1.0 cm aproximadamente.
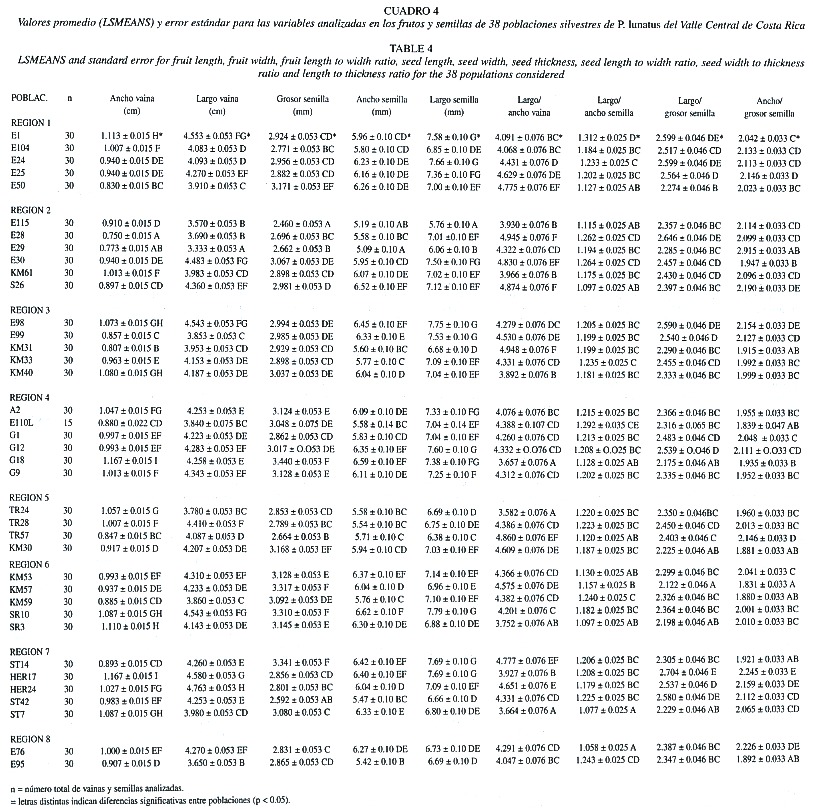
El largo de las vainas promedio de cada población varía entre 3.333 (en la región 2) y 4.763 cm (en la región 7) (Cuadro 4), sin embargo, la mayoría de poblaciones presentan en promedio más de 4.0 cm de longitud. Estos resultados coinciden con el valor de 4.15 ± 0.42 cm (Cuadro 2) calculado a partir de las las 1125 vainas que se midieron en las 38 poblaciones estudiadas.
Si se agrupan las poblaciones por regiones, se observa que, tanto para el ancho como para el largo de las vainas, las regiones 2 y 8 muestran , en promedio, valores más pequeños que las demás regiones (Cuadro 5, Figura 2). Por otra parte, las regiones 4, 6 y 7 presentan, en promedio, los valores más grandes. En ambos casos las regiones 3 y 5 presentaron, en promedio, valores intermedios entre estos dos extremos.
La proporción largo/ancho de vaina se calculó con el objetivo de determinar de manera aproximada, la forma de los frutos. El análisis de esta proporción también muestra diferencias significativas entre las ocho regiones, indicando variación en la forma de los frutos. En 38 de las poblaciones estudiadas, los valores para esta relación oscilaron entre 4.047 y 4.948. Estos resultados indican que en la mayoría de poblaciones (79%) las vainas son por lo menos cuatro veces mas largas que anchas (Cuadro 5). Cuando se analiza la variación por regiones, se observa que las regiones 4 y 8 presentan en promedio los valores más pequeños, es decir frutos menos alargados, mientras que para la región 2 se muestra el valor más grande (Figura 2). Sin embargo, debido a que para las ocho regiones los promedios varían entre 4.2 y 4.5 (Cuadro 5), se puede decir que, en términos generales, la forma de los frutos se mantiene relativamente constante entre regiones.
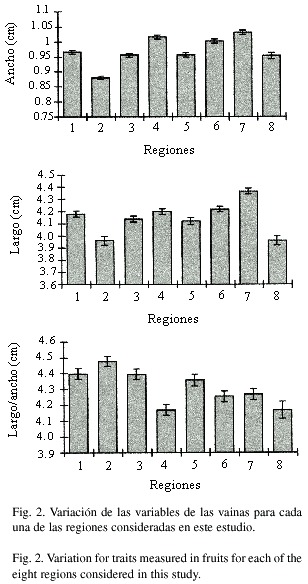
En las semillas, las tres variables que se utilizaron para evaluar la variación morfológica dentro y entre poblaciones silvestres de frijol lima son el largo, el ancho y el grosor. En términos generales, se considera que la variación morfológica de las semillas es poco afectada por las condiciones ambientales. En las 38 poblaciones analizadas, el ancho de las semillas varió entre 5.09 y 6.62 mm, el largo entre 5.76 y 7.79 mm y el grosor entre 2.46 y 3.44 mm según la población de origen (Cuadro 3). Si se unen todas las poblaciones y se calculan los promedios a partir de una muestra de 1 125 semillas para cada una de las variables (Cuadro 4), se observa que en promedio el grosor de las semillas es de 2.97 ± 0.36 mm, el ancho es de 6.00 ± 0.66 mm y el largo de 7.07 ± 0.69 mm. En términos generales, las semillas en esta especie son relativamente pequeñas y delgadas. Con base en el promedio para el ancho y largo, se puede decir que la mayoría de semillas son ligeramente más largas que anchas. Sin embargo, como la variación en muchas de estas semillas es de aproximadamente un milímetro, a simple vista es difícil detectarla.
Si se comparan estas tres características morfológicas entre regiones, se puede observar que las regiones 2, 5 y 8 muestran, en promedio, los valores más pequeños para el ancho, largo y grosor de las semillas (Figura 3). Las otras regiones se ubican en los extremos o en posiciones intermedias y varían según la característica morfológica que se analice. Nuevamente, se demuestra que las diferencias entre valores extremos para estas variables morfológicas son estadísticamente significativas, aún cuando en la mayoría de las comparaciones entre pares de regiones estas diferencias no son significativas (Cuadro 3).
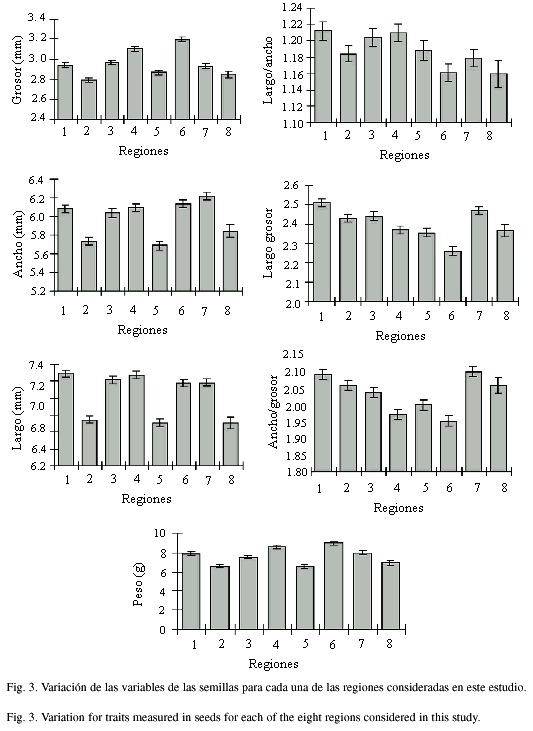
Con el objetivo de determinar la forma de las semillas, se calcularon las proporciones largo/ ancho, largo/grosor y ancho/grosor entre poblaciones y entre regiones. En las 38 poblaciones estudiadas, estas proporciones variaron en promedio entre 1.077 y 1.312 mm para el largo/ ancho, entre 2.122 y 2.646 mm para el largo/ grosor y 1.839 y 2.915 mm para el ancho/grosor (Cuadro 4). Valores muy cercanos a uno para la relación largo/ancho, indican que el largo y ancho en las semillas son muy semejantes y que por lo tanto la forma de las semillas es cuadrada. Estos resultados se corroboran fácilmente con sólo observar y comparar las semillas en diferentes poblaciones silvestres de frijol lima. Al agrupar las poblaciones por regiones se observa que para la relación largo/ ancho de las semillas, las regiones 6 y 8 presentan los valores más pequeños, mientras que las regiones 1 y 4 muestran los valores más grandes (Figura 3). Sin embargo, tanto para esta variable como para el largo/grosor y ancho/grosor el ámbito de variación observada entre regiones es relativamente pequeño (Cuadro 5, Figura 3). Estos resultados indican que, si bien es cierto se mantiene una tendencia de las regiones 2, 5 y 8 de presentar en promedio valores mas pequeños para el grosor, ancho y largo de las semillas, cuando se hace la relación entre estas variables, la proporción entre la forma y tamaño de las semillas se mantiene más o menos constante entre regiones.
Una variable adicional de las semillas que sirve para caracterizarlas física y químicamente es el peso de las mismas. En general, el peso en las semillas está relacionado no sólo con su tamaño, sino también con la cantidad y tipo de constituyentes que almacenan. Tal y como se indicó anteriormente, las poblaciones silvestres de frijol lima que crecen en el Valle Central de Costa Rica presentan semillas relativamente pequeñas. Por lo tanto, para calcular el peso seco, se pesaron 100 semillas por cada repetición. Hasta donde fue posible se hicieron tres repeticiones por población, sin embargo, si las semillas no eran suficientes se hicieron únicamente dos repeticiones por población.
En el Cuadro 6, se compara la variación en el peso de las semillas entre poblaciones. A pesar de la poca variación observada al comparar entre el largo y ancho de las semillas, el peso seco promedio de 100 semillas varió entre 4.61 y 11.88 g. Si se agrupan todas las poblaciones, el valor promedio que se obtiene para las 87 repeticiones es de 7.6 ± 1.3 g (Cuadro 2). Al comparar los pesos promedio entre regiones, al igual que para las otras cinco características analizadas hasta ahora, las regiones 2, 5 y 8 presentan los valores más bajos para el peso (Figura 3). En el otro extremo, las regiones 4 y 6 muestran los pesos más altos. En una posición intermedia se ubican las regiones 1, 3 y 7. Es importante destacar que estas tendencias, observadas para el peso promedio de 100 semillas, coinciden con muchos de los resultados obtenidos para las otras variables morfológicas.
Discusion
El objetivo de este estudio fue determinar los niveles de variación en las dimensiones de frutos y semillas de 38 poblaciones silvestres de Phaseolus lunatus en el Valle Central de Costa Rica. Los resultados de este estudio indican que existe un alto nivel de variación en los caracteres de los frutos y semillas de Phaseolus lunatus en el área considerada en este trabajo. Los datos indican que existe una amplia variación en el tamaño de los frutos y semillas y esta variación se observa tanto entre regiones, como entre poblaciones y aún entre semillas de una misma población. Esto es sobresaliente si se toma en consideración que las 38 poblaciones silvestres de frijol lima incluidas en este estudio se distribuyen en un área relativamente pequeña (2100 km2 ) y dentro de un mismo centro de diversidad (Mesoámerica).
La mayoría de los estudios sobre la variación en el tamaño de las semillas de P. lunatus, se han hecho con base en el peso de las mismas (Maquet 1995, Esquivel et al. 1990) . Sin embargo, en este trabajo no sólo se evaluaron cuatro características morfológicas de las semillas (largo, ancho, grosor y peso), sino que se realizaron un gran número de repeticiones para cada variable. El hecho de evaluar diferentes características morfológicas y un gran número de repeticiones por cada variable en cada una de las 38 poblaciones seleccionadas nos permite predecir, con bastante seguridad que una variación similar se podría encontrar en el resto de poblaciones silvestres que crecen en esta región de Costa Rica.
Varios autores han propuesto que el tamaño de las semillas es una de las características morfológicas más estables en las plantas (Harper 1977). Ellos proponen que la planta madre hace una distribución equitativa de los recursos en la progenie. Tanto por ser el producto directo de la reproducción sexual (la cual le confiere variabilidad y mayores posibilidades de adaptación a la progenie) como por ser el principal medio para la dispersión de la especie, un tipo de selección estabilizadora debería favorecer la constancia en el tamaño de las semillas dentro de una misma especie (Harper et al. 1970). Sin embargo, numerosos estudios demuestran que el tamaño de las semillas puede variar dentro de las poblaciones y dentro de plantas en una misma especie (Janzen 1977, Cavers y Steel 1984, Winn y Gross 1993, Sakai y Sakai 1995, Sakai y Sakai 1996). Especialmente, algunos autores afirman que en especies que presentan crecimiento indeterminado el peso de las semillas es más variable (Wulff 1986, Obeso 1993, Rocha y Stephenson 1995). En el caso de las poblaciones estudiadas, todas silvestres y con crecimiento indeterminado, se encontró alta variabilidad lo que concuerda con lo predicho por estos mismos autores. Esto parece indicar que, aún cuando el peso de las semillas está determinado genéticamente, las diferencias en la disponibilidad de recursos del medio ambiente o durante la ontogenia de la planta tienen un efecto importante sobre el tamaño de las semillas.
En otras especies del género Phaseolus, el peso de las semillas varía de manera inversa en relación al número de semillas por fruto (Nakamura 1988, Rocha y Stephenson 1990). Además, en el caso de P. vulgaris, el número de semillas por vaina es menor en aquellos frutos que provienen de autofertilizaciones y las semillas son más grandes que aquellas que provienen de fertilización cruzada (Nakamura 1986, 1988). Por otro lado, en P. coccineus, el peso de las semillas es también afectado por el genotipo de la planta que las produce y por el número de semillas en el fruto (Rocha y Stephenson 1990). En P. lunatus, donde cada población presenta una diversidad genética muy baja (Maquet et al. 1996, Vargas et al. 2001) y existe una alta diferenciación entre poblaciones, no es sorprendente encontrar una alta heterogeneidad entre poblaciones en el peso de las semillas.
Se conoce muy poco sobre la relación que pueda existir entre la variación geográfica y el tamaño y peso de las semillas. Winn y Gross (1993) encontraron muy poca variación en el peso de las semillas de Prunella vulgaris a lo largo de un gradiente latitudinal. En forma natural, las poblaciones de frijol lima se distribuyen desde los 800 hasta los 1700 m.s.n.m. En este ámbito de distribución altitudinal, las poblaciones que componen las regiones 2 y 6 están ubicadas a una menor altura que las demás poblaciones, mientras que las poblaciones de las regiones 1 y 4 están a la mayor altura. A pesar de estas diferencias tan marcadas, no se encontró ninguna tendencia que nos permita relacionar la altitud con la variación en el peso de las semillas. Cabe destacar que, a pesar del aparente aislamiento geográfico entre algunas de las ocho regiones definidas en este estudio, existe mayor variación entre las poblaciones de cada región que entre regiones. Esto no necesariamente significa que los factores ambientales tienen una contribución muy pequeña en la determinación del tamaño de las semillas, sino más bien puede indicar todo lo contrario. Esto por cuanto dentro de cada región propuesta existen diferencias en elevación, precipitación promedio anual, temperatura promedio anual y otras variables microambientales como tipo de suelo, etc, que pueden tener un papel muy importante en la determinación del tamaño de las semillas.
Los factores que pueden contribuir a la variación morfológica de las semillas entre y dentro de las poblaciones son muy diversos. Se sabe muy bien que para la mayoría de las especies existe una relación directa entre el tamaño de las semillas y el tamaño de las plántulas (Harper 1977, Rocha y Stephenson 1991, 1995). Es obvio, que un mayor tamaño en los primeros estadios de desarrollo le confiere ventajas a la planta. No obstante, también es cierto que es más fácil la dispersión de semillas pequeñas, sobre todo en especies como el frijol lima que no son dispersadas por animales. En condiciones de estrés por falta de nutrimentos, las plantas también pueden hacer una distribución desigual de recursos para la progenie. En
estas circunstancias, a pesar de que parte de las semillas morirán por falta de recursos, la planta se asegura de que al menos una parte de la progenie tendrá recursos suficientes para germinar y desarrollarse. Sin embargo, la mayoría de las plantas producen mas flores y óvulos que el número de frutos y semillas que estas llegan a producir (Stephenson 1981, Rocha y Stephenson 1991). Esto se asocia al hecho de que en esas especies la reproducción está limitada por la disponibilidad de recursos. Por ese motivo, el tamaño y el número de frutos y semillas es una respuesta tanto a las condiciones ambientales que afectan la disponibilidad de recursos como al genotipo de la planta.
Los resultados obtenidos en este trabajo coinciden con lo reportado por otros autores (Rocha et al. 1997, 2002, Vargas et al. 2001). Maquet et al. (1996) estudiaron la variación de isoenzimas en 21 poblaciones silvestres de frijol Lima en el Valle Central de Costa Rica. Sus resultados indican que existe poca variación dentro de las poblaciones y mucha variación entre las poblaciones (Gst = 0.803), además, también se encontró una marcada deficiencia de heterocigotos. Los autores señalan que esta distribución de la variación genética es típica de las especies que presentan altos niveles de autofertilización, lo que parece ser respaldado por el valor del índice de fijación reportado en ese estudio (F = 0.842). Pero Rocha et al. (1997) proponen que estas poblaciones pueden estar sufriendo severos cuellos de botella como consecuencia de las perturbaciones causadas por las prácticas agrícolas de las zonas en que habitan. Rocha et al. (1997, 2002) demostraron que las poblaciones silvestres de P. lunatus experimentan frecuentes episodios de extinción local y recolonización y proponen que este proceso debe ser considerado para explicar los niveles de diferenciación genética entre las poblaciones, ya que aceleran los procesos de deriva genética.
Otros autores han examinado el posible efecto de las extinciones locales y las subsiguientes recolonizaciones sobre la diferenciación genética de las poblaciones (Slatkin 1977, 1987, Wade y McCauley 1988, McCauley 1993, Gliddon y Gaudet 1994). Slatkin (1977) señala dos factores que determinan el resultado de estos procesos. En algunos casos pueden ocurrir procesos de muestreo no aleatorio entre los individuos de la población durante la producción de los propágulos que recolonizan un sitio, previamente a la extinción, causando un severo cuello de botella y la pérdida de alelos en la población. En otros casos, la dispersión de propágulos desde las poblaciones vecinas contribuye a restaurar la diversidad genética y evita la diferenciación entre las poblaciones. Esto permite concluir que la magnitud y la dirección de los efectos de las extinciones locales y la recolonización dependen del papel que juegen la deriva génica y el flujo génico entre las poblaciones (Slatkin 1977, 1987, Wade y McCauley 1988, McCauley 1993). Mas aún, Gliddon y Goudet (1994) concluyen que las poblaciones que sufren este tipo de procesos deben ser estudiadas utilizando un enfoque de metapoblaciones, y que este enfoque es particularmente útil para generar predicciones que contribuyan a resolver problemas propios de la conservación biológica. Los trabajos de Rocha et al. (1997, 2002) sustentan esas conclusiones.
En conclusión, los resultados de este estudio demuestran que existe bastante variación en el tamaño de los frutos y semillas en las poblaciones silvestres de Phaseolus lunatus localizadas en el Valle Central de Costa Rica. Además, la mayor parte de la variación se distribuye entre poblaciones y no entre las regiones geográficas definidas en este estudio. Esto no significa que los factores ambientales jueguen un papel pequeño en la determinación del tamaño de los frutos y semillas, sino más bien, parece indicar que las regiones geográficas definidas en este estudio son demasiado arbitrarias y carecen de homogeneidad. Sin embargo, este patrón de variación también puede ser el resultado de un flujo genético muy amplio en el pasado y que el aislamiento actual aún no ha alcanzado a generar diferencias genéticas entre las regiones. Estos resultados tienen relevancia para el desarrollo de estrategias de manejo de estas poblaciones con el fin de evitar erosión genética en ellas, indicando que es más importante conservar un mayor número de poblaciones antes que pocas poblaciones de gran tamaño. Esto por cuanto la variación entre poblaciones es significativa. Por otro lado, también es importante distribuir las poblaciones seleccionadas a todo lo largo y ancho del área de estudio para poder proteger la variación existente entre las diferentes regiones geográficas.
Agradecimientos
Los autores agradecen a M.E. Zaldivar, P. Hoc, Walter Marín y Julián Monge por recomendaciones y críticas realizadas a versiones anteriores de este manuscrito. A L. Castro, G. Aguilar y A. Quirós por su ayuda en el trabajo de campo y laboratorio. Al Belgian Administration for Development Cooperation (BADC) por el respaldo financiero y al International Plant Genetic Resources Institute (IPGRI) por la administración de este proyecto y su apoyo incondicional.
Resumen
Se estudió la diversidad morfológica de frutos y semillas de 38 poblaciones silvestres de Phaseolus lunatus var. lunatus (frijol lima) en el Valle Central de Costa Rica. Para ello se determinó la variación en el largo y ancho de vaina, el largo, ancho y grosor de las semillas, la razón entre estas variables y el peso promedio de 100 semillas. En términos generales, se puede afirmar que existen diferencias significativas entre las poblaciones analizadas para todas estas variables. Cuando se agruparon las 38 poblaciones en ocho regiones geográficas dentro del área de estudio, se encontró que existían diferencias significativas entre distintas regiones, pero la variación entre las poblaciones de cada región era mayor que aquella encontrada entre regiones. Estos resultados sugieren que no existe asociación entre estas variables y la región geográfica. Se analizó la utilidad de estos resultados para el desarrollo de estrategias para la conservación in situ de las poblaciones silvestres de Phaseolus lunatus.
Referencias
Allen, G., J. Antos, A. Worley, T. Suttill & R. Hebda. 1996. Morphological and genetic variation in disjunct populations of the avalanche Lily Erythronium montanum. Can. Jour. Bot. 74: 403-412 [ Links ]
Annicchiarico, P. & L. Pecetti. 1994. Morpho-physiological traits as descriptors for discrimination of durum wheat germplasm. Genet. Res. Crop Evol. 41: 47-54. [ Links ]
Anónimo, 1985. SAS users guide: statistics. SAS Institue, Cary, N.C. [ Links ]
Ariyo, O.J. 1993. Genetic diversity in west african okra (Abelmoschus caillei) (A. Chev.) Stevels – Multivariate analysis of morphological and agronomic characteristics. Genet. Res. Crop Evol. 40: 25-32. [ Links ]
Baudoin, J. 1988. Genetic resources, domestication and evolution of lima bean (Phaseolus lunatus), pp. 393-408. In P. Gepts (ed.). Genetics Resources of Phaseolus Beans. Kluwer Academic, Dordrecht, The Netherlands. [ Links ]
Cavers, P. & M. Steel. 1984. Patterns of change in seed weight over time on individual plants. Amer. Nat. 124: 324-335. [ Links ]
Chandran, K. & S.M. Pandya. 2000. Morphological characterization of Arachis species of section Arachis. Plant Genet. Res. News. 121: 38-41. [ Links ]
Clements, J. & A. Cowling. 1994. Patterns of morphological diversity in relation to geographical origins of wild Lupinus angustifolius from the aegean region. Genet. Res. Crop . Evol. 41: 109-122. [ Links ]
Debouck, D., A. Maquet & C. Posso. 1989. Biochemical evidence for two different gene pools in lima beans, Phaseolus vulgaris L. Ann. Rep. Bean Improvement Coop. 32: 58-59. [ Links ]
Esquivel, M., L. Castiñeiras, K. Hammer. 1990. Origin, classification, variation and distribution of lima bean (Phaseolus lunatus) in the light of cuban material. Euphytica 49: 89-97. [ Links ]
Ford-Lloyd, B.V. & M. Jackson. 1986. Plant genetic resources: an introduction to their conservation and use. Edward Arnold, London, U.K. 152 p. [ Links ]
Frankel, O.H, A.D.H. Brown & J.J. Burdon. 1995. The conservation of plant biodiversity. Cambridge University, Cambridge, U.K. [ Links ]
Gliddon, C. & J. Goudet. 1994. The genetic structure of metapopulations and conservation biology. pp. 107- 114. In V. Loeschke, J. Tomiuk & S.K. Jain (eds.). Conservation Genetics. Birkhauser, Germany. [ Links ]
Guarino, L.,V. R. Rao & R. Reid. 1995. Collecting plant genetic diversity, technical guidelines. CAB International, Cambridge, U.K. 748 p. [ Links ]
Hamrick, J.L. and M.J.W. Godt. 1989. Allozymer diversity in plants. pp. 43-63. In A.D.H. Brown, M.T. Clegg, A.L. Kahler & B.S. Weir (eds.). Plant population genetics, breeding, and genetic resources. Sinauer Associates, Sunderland, Massachussetts, USA. [ Links ]
Harper, J. 1977. Population biology of plants. Academic Press, New York, New York, USA. [ Links ]
Harper, J., P. Lovell, K. Moore. 1970. The shapes and sizes of seeds. Ann. Rev. Ecol. Syst. 1: 327-356. [ Links ]
Hoc, P.S., S.M. Espert, S.I. Drewes & A.D. Burghardt. 2003a. Polimorfismo en Phaseolus vulgaris var. aborigineus (Fabaceae). Evidencias que indican hibridación natural. Rev. Biol. Trop. 51(2): 369-380. [ Links ]
Hoc, P.S., S.M. Espert, S.I. Drewes & A.D. Burghardt. 2003b. Polimorfismo en Phaseolus vulgaris var. aborigineus (Fabaceae). Evidencias que indican hibridación natural. Rev. Biol. Trop. 51(4): en prensa. [ Links ]
Janzen, D. 1977. Variation in seed size within a crop of a Costa Rica Mucuna andreana (Leguminosae). Amer. J. Bot. 64: 347-349. [ Links ]
Letschert, J. & L. Frese. 1993. Analysis of morphological variation in wild beet (Beta vulgaris ) from Sicily. Genet. Res. Crop. Evol. 40: 15-24. [ Links ]
Lioi, L., M. Esquivel, L. Castañeiras & K. Hammer. 1991. Lima bean (Phaseolus lunatus) landraces from Cuba: electrophoretic analysis of seed storage proteins. Biol. Zent. bl. Heft 1: 76-79. [ Links ]
McCauley, D.E. 1993. Evolution in metapopulations with frequent local extinction and recolonization, pp. 109- 134. In D. Futuyma & J. Antonovics (ed.). Oxford Surveys in Evolutionary Biological. Oxford Univ., Oxford. [ Links ]
Maquet, A. 1995. Study on Genetic Diversity of the Legume Phaseolus lunatus Using Morphophysiological and Protein Markers. These de Doctorat. Gembloux, Faculté Universitaire des Sciences Agronomiques, Belgium, 188 p. [ Links ]
Maquet. A., I. Zoro, O. Rocha, J.P. Baudoin. 1996. Case studies on breeding systems and its consequences for germoplasm conservation. Genet. Res. Crop Evol. 43: 309-318. [ Links ]
Nakamura, R. 1986. Maternal investment and fruit abortion in Phaseolus vulgaris. Amer. Jour. Bot. 73: 1049-1057. [ Links ]
Nakamura, R. 1988. Seed abortion and seed variation within fruits of Phaseolus vulgaris: pollen donor and resource limitation effects. Amer. Jour. Bot. 75: 1003-1010. [ Links ]
Nevo, E., E. Golenberg & A. Beiles. 1982. Genetic diversity and environmental associations of wild wheat, Triticum dicoccoides, in Israel. Theor. Appl. Genet. 62: 241-254. [ Links ]
Nevo, E., A. Beiles & N. Storch. 1983. Microgeographic edaphic differentiation in hordein polymorphisms of wild barley. Theor. Appl. Genet. 64: 123-132. [ Links ]
Nevo, E., A. Beiles & T. Krugman. 1988. Natural selection of allozyme polymorphims: a microgeographical differentiation by edaphic, topographical, and temporal factors in wild emmer wheat (Triticum dicoccoides). Theor. Appl. Genet. 76: 737-752. [ Links ]
Nevo, E., I. Noy-Meir, A. Beiles, T. Krugman & M. Agami. 1991. Natural selection of allozyme polymorphisms: microgeographical spatial and temporal ecological differentiations in wild emmer wheat. Israel J. Bot. 40: 419-449. [ Links ]
Obeso, J. 1993. Seed mass variation in the perennial herb Asphodelus albus: sources of variation and position effect. Oecologia 93: 571-575. [ Links ]
Polignano, G.B., P.Uggenti &G. Scippa. 1993. The patterns of genetic diversity in faba beans collections from Ethiopia and Afghanistan. Genet. Res. Crop Evolut. 40: 71-75. [ Links ]
Prance, G.T. 1997. The conservation of botanical diversity. pp 3-14. In N. Maxted, B.V. Ford-Lloyd & J.G. Hawkes (eds.). Plant conservation biology: the in situ approach. Chapman and Hall, Reading, U.K. [ Links ]
Rocha, O.J. & A.G. Stephenson. 1990. Effect of ovule position on seed production, seed weight, and progeny Perfomance in Phaseolus coccineus L. (Leguminosae). Amer. J. Bot. 77: 1320-1329. [ Links ]
Rocha, O.J. & A.G. Stephenson. 1991 Effects of nonrandom seed abortion on progeny performance in Phaseolus coccineus L. Evolution 45: 1198-1208 [ Links ]
Rocha, O.J. & A.G. Stephenson. 1995. Regulation of flower, fruit, and seed production: Phaseolus coccineus a study case, pp. 245-262. In P.C. Hoch & A.G. Stephenson (eds.). Experimental and molecular approaches to plant biosystematics. Monograph in Systematic Botany from the Missouri Botanical Gardens. Vol 53. [ Links ]
Rocha, O., G. Macaya & J. Baudoin, 1997. Causes of local extintion and recolonization, determined by 3 years of monitoring wild populations of Phaseolus lunatus L. in the Central Valley of Costa Rica. Plant Genet. Res. News. 112: 44-48. [ Links ]
Rocha, O.J., J. Degreff, D. Barrantes, E. Castro & G. Macaya. 2002. Metapopulation dynamics of lima beans (Phaseolus lunatus) in the Central Valley of Costa Rica, pp. 205-215. In J.M.M. Engels, A.H.D. Brown & M.T. Jackson (eds.). Managing Plant Genetic Resources. CABI International, UK. [ Links ]
Sakai, S. & Sakai, A. 1995. Flower-dependent variation in seed size: theory and a test. Amer. Nat. 145: 918-934. [ Links ]
Sakai, S., A. Sakai. 1996. Why is there variation in mean seed among plants within single populations? Test of the fertilization efficiency hypothesis. Amer. J. Bot. 83: 1454-1457. [ Links ]
Slatkin, M. 1977. Gene flow and genetic drift in a species subject to frequent local extinctions. Revista 12: 253-262. [ Links ]
Slatkin, M. 1987. Gene flow and the geographic structure of natural population. Science 236: 87-92. [ Links ]
Stephenson, A.G. 1981. Flower and fruit abortion: proximate cause and ultimate functions. Ann. Rev. Ecol. Syst. 12: 253-279. [ Links ]
Vargas, E.M., G. Macaya, J.P. Baudoin & O.J. Rocha. 2001. Case studies in breeding systems and its consequences for germplam conservation: Electrophoretic mobility of phaseolins in wild populations of lima beans (Phaseolus lunatus) in the Central Valley of Costa Rica. Genet. Res. Crop Evol. 48: 109-120. [ Links ]
Wade, M.J & D.E. McCauley. 1988. Extinction and recolonization: Their effects on the genetic differentiation of local populations. Evolution 42: 995-1005. [ Links ]
Winn, A., & K. Gross. 1993. Latitudinal variation in seed weight and flower number in Prunella vulgaris. Oecología 93: 55-62. [ Links ]
Wulff, R. 1986. Seed size variation in Desmodium paniculatum. I. Factors affecting seed size. J. Ecol. 74: 87-89. [ Links ]
Wulff, K. 1991. Analysis of allozyme variability in three Plantago species and a comparison to morphological variability. Theor. Appl. Genet. 81: 119-126. [ Links ]

