











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.51 no.1 San José Mar. 2003
Abstract
Decapod community inhabiting the rocky littoral of eight bays from Socorro island, Revillagigedo Archipelago, was studied during March 1992. Samples were collected from one-square meter plots placed following transects drawn perpendicularly to the shoreline along the intertidal zone. Species richness and abundance were determined in each bay, as well as the similarity between bays using Morisitas index. Organisms collected belong to six families, 20 genera and 26 species of the Brachyura and Anomura infraorders. Xanthidae was the family with the highest number of species, and the highest species richness was recorded at the V. Lozano bay. Pachygrapsus transversus, Xanthodius cooksoni and Calcinus explorator were the most abundant species in nearly all localities. Dendrogram obtained define two groups of bays at a similarity level above 0.67, given the similarity in the crabs species richness and their abundance between bays in the island.
Key words: brachyurans, anomurans, richness, abundance, rocky coast, Mexico.
The Revillagigedo Archipelago, Mexico, is constituted by the Clarion, San Benedicto, Roca Partida and Socorro islands. The latter is the largest of all, and because of its volcanic origin, it offers a wide variety of habitats for the development of various kinds of benthonic invertebrates. Research works on the carcino-logical fauna for this Archipelago are scarce. The studies by Rathbun (1918, 1925, 1930), included some species collected in any of these islands, while those by Hernández-Aguilera et al. (1986) and Hernández-Aguilera and Martínez-Guzmán (1992) refer to decapods and stomatopods and were conducted at Clarion. Socorro Islands carcinofauna is less well studied, research works were carried out more than 40 years ago and they basically provide descriptions and lists of species. In her publications on the carcinological fauna of the Pacific Ocean, Rathbun describes some species that were collected in Socorro Island.
Afterwards, Garth (1958) reports majid species collected during the Allan Hancock Societys expeditions in this island; likewise Haig (1960) includes the species collected in these same expeditions in his monograph on porcellanids. Adem et al. (1960) notify the presence of the Xanthidae and Grapsidae families at the southern portion of the island. Garth (1992) publishes a study of brachyuran crabs from the Revillagigedo islands, emphasizing their insular endemism, based on the 1934 and 1939 collection trips carried out by the "Velero" ship. Last, Hendrickx (1995a, b) includes some of the species present in the island in his review about decapod crabs of the eastern tropical Pacific.
To date no quantitative descriptions of the carcinofauna inhabiting Socorro Island have been reported. For this reason, this paper reports the species richness, abundance and distribution of brachyuran and anomuran crabs found inhabiting the rocky coast of the bays located to the north and south of this island as an attempt to broaden the ecological knowledge of these taxonomical groups in the island.
Materials and methods
Study area: Socorro Island (Fig. 1), with an extension of 150 km 2 , is located at 18º47 North Lat. and 110º 59 West Long., 480 km from the southern edge of Baja California and 716 km off Manzanillo, Colima (Adem et al. 1960). It has several bays: three to the south (V. Lozano, Braithwaite and Binners), one to the southwest (Grayson), one to the northwest (Blanca) and one to the north (North Bay), the latter two with two inlets each. All bays are frequently washed by strong waves and are inhabited at its central portion by small coral reef colonies at depths between 4 and 8 m. Rest of the islands littoral is inaccessible because of its cliffs and rocky outcrops. Abrief description of the main features in these bays follow:
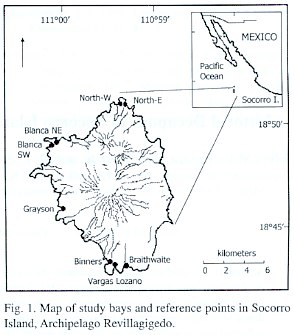
Vargas Lozano. Constituted by fragmented and eroded biogenic elements and by sand, gravel and igneous rocks of variable dimensions. At one end it is delimited by a cliff with fissures, cavities and large rocks with fragments of coral or gravel under them, and forming some small intertidal pools. The other end is made of large, relatively unstable rocks.
Binners. It includes a beach integrated by coarse coral reef elements along with gravel of igneous and biogenic origin. On one end there is an area of large eroded rocks with rocky out-crops; on the other the bay changes to a wide and low rocky coast with intertidal pools, fissures and cavities.
Braithwaite . An open bay that includes at its central portion large eroded rocks. It is delimited at both ends by compacted rocks, which include areas of rocky substrate in plateaus with a smooth slope, delimiting a wide supra- and mesolittoral fringe washed by the sea. The wavefront shows numerous crevices, caverns, cavities and intertidal pools.
Grayson. Small inlet that includes small crevices, cavities and a small sand plain with an intertidal zone constituted by large unstable stones along a steep slope. It is surrounded by abrupt cliffs and loose crags.
Blanca. Constituted by two inlets separated from one another by a prominent rocky out-crop.
The northeastern inlet (Blanca NE), is relatively shallow with a smoothly-sloped sub-strate including dead coral reef elements, flat stones cover by filamentous algae and abundant organic matter. The southwestern inlet (Blanca SW) is a small bay that includes large stones with small tide pools and rocky out-crops at its central portion.
North Bay. It comprises two small bays separated by a volcanic formation. The eastern bay (North-E) has a moderate slope and a sub-strate made of fine sand. At one of its ends there is an abrupt high cliff formed by nonconsolidated materials, at the other end the coast is made of a low rocky outcrop. The western bay (North-W) is sandy, at one end is limited by a fringe of large round stones, crags and rocky outcrops including crevices, cavities and intertidal pools with a permanent water exchange. Avertical cliff limits the other end.
One expedition was conducted to Socorro Island during March, 1992 and included all bays mentioned above. In each bay, sampling included the supralittoral and high- and mid-mesolittoral
zones. It was done along transects placed perpendicularly to the coastline, separated from one another by 20 m approximately. Each transect included several one-square meter plots located at 2 m intervals (Miles and Miles 1966, Holme and McIntyre 1984). The number of plots sampled in each bay depended on the extension of the bay as well as on the conditions of tides and surges that prevailed during sampling (Mille-Pagaza et al. 1994). It is worth mentioning that during this period a wide portion of the intertidal zone was accessible because of the occurrence of a particularly pronounced low tide.
Sampling was done manually by lifting rocks and crushing portions of dead coral.
Determination of crab species was based on identification keys and descriptions by Rathbun (1918, 1930), Garth (1958), Haig (1960, 1966), Brusca (1980), García-Madrigal (1994) and Hendrickx (1995b).
Species richness, defined as the number of species present (Krebs 1994, Stiling 1999), total density (number of organisms/100 m2 ) and relative abundance of each species were determined. Similarity among bays was calculated by means of the Morisitas index, and these values were used to classify the bays, grouping them according to the Unweighed Per-Group Method using Arithmetic Averages (UPGMA) (Ludwig and Reynolds 1988, Krebs 1989).
Results
Organisms registered belong to four families, 17 genera and 19 species of the Brachyura infraorder, and two families, three genera and seven species of the Anomura infraorder. Taxonomical groups with the highest species richness (Table 1) were Majoidea, Xanthidae and Grapsidae with seven, seven and four species, respectively. Among anomurans, porcellanids showed the highest species richness, with six species. Bays with the highest species richness were V. Lozano and Blanca NE, with 15 and 16 species, respectively (Table 1).
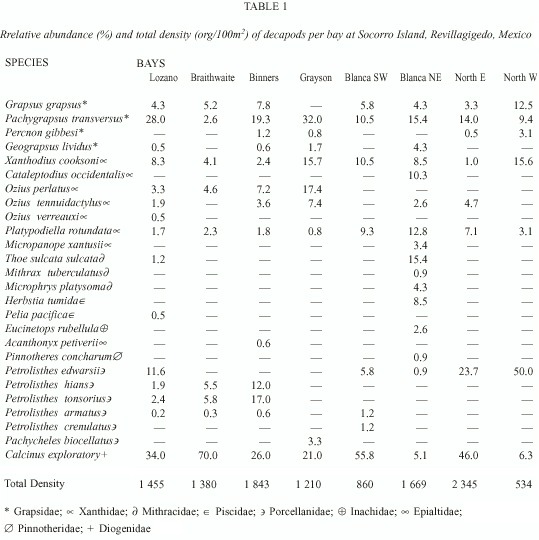
In general, grapsids and xanthids had the widest distribution in the island, with several species present in all bays. By contrast, Pinnotheridae had the most restricted distribution, being found only at Blanca NE with a very low abundance. Species with the widest distribution were Grapsus grapsus (Linnaeus 1758) and Pachygrapsus transversus (Gibbes 1850), Xanthodius cooksoni (Miers 1877), Platypodiella rotundata (Stimpson 1860) and Calcinus explorator Boone 1932.
The highest total density was registered at Binners and North-E (1 843 and 2 345 org./100 m2 , respectively). Grapsidae represent the dominant family in the island, with the Diogenidae anomurans also dominating in some bays. Xanthids tend to be less dominant, having intermediate values, whereas majids present very low abundance in general (Tabla 1).
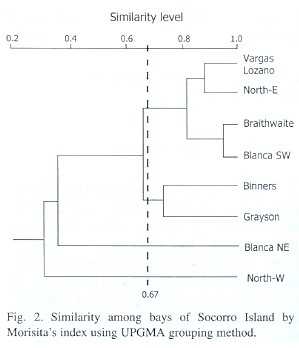
Dendrogram (Fig. 2) obtained from the Morisitas index showed that two groups are formed at 0.67 level, the first one also with two subgroups, one including Blanca SW and Braithwaite, with the highest similarity (0.95) and the other including North-E and V. Lozano with a similarity of about 0.85. Another group, with a lower similarity (0.75), was constituted by Grayson and Binners.
Last, Blanca NE and North-W bays appear as the most dissimilar ones.
Discussion
Species richness recorded in March 1992, particularly at the V. Lozano bay, can be attributed to habitats complexity (Abele 1974, Birch 1981, Raffaelli and Hawkins 1997, Krohne 1998). This bay provides protection and food to several grapsid, xanthid and porcellanid crabs. P. transversus is one of the species that stand out, developing abundantly in fissures and cavities, from which it emerges at low tide to feed on algae adhered to rocks, like other species of this genus in several other localities (Abele 1974, Ricketts et al. 1985). Xanthids remain among gravel or pieces of coral below rocks, where they feed on detritus; and porcellanids find shelter among crevices or under rocks, where they filter diatoms and organic detritus as they do at northern locations of the American Pacific coast (Ricketts et al. 1985). At Blanca NE the highest diversity was achieved by majids in relation to a particular habitat constituted by a zone of super-imposed flat rocks, with abundant filamentous algae and that retain slime among which these crabs live, constituting very characteristic associations (Sanders 1968).
In Socorro Island there are species that inhabit different habitats, like C. explorator and P. transversus, which can be found below and above rocks and even at intertidal pools. By contrast, there are species which prefer a particular habitat type, as in the case of majids, as well as Cataleptodius occidentalis (Stimpson, 1871) and Pinnotheres concharum, that were found associated with flat rock-algae-slime, or Acanthonyx petiveri Milne Edwards 1834 that was only collected at Binners bay in intertidal pools with algal growth, possibly forming part of another kind of association, or Pachycheles biocellatus and Petrolisthes crenulatus, only present in one of the bays associated with fractions of coral reef below rocks.
Species richness found in this work (26 species) does not coincide with the data reported by Garth (1992), who mentions 36 brachyuran species for Socorro Island collected during several expeditions. This is attributed to the fact that Garth included species found at the infralittoral fringe. Composition by genera partially coincides with data reported by Hernández-Aguilera et al. (1986) and Garth (1992), since they only mention Acanthony x, Herbstia, Cataleptodius, Ozius and Geograpsus as part of the carcinofauna of Clarion. Despite the fact that Gecarcinus planatus was frequently seen in all bays, this species was not included in any of our plots. Likewise, it is worth noting that Xanthodius hebe s, mentioned by Llinas-Gutiérrez et al. (1993) as an inhabitant of intertidal pools, was neither found in any bay nor it even appears included in Garths lists (1992).
Differences in total density among bays can also be explained as a function of habitat complexity (Abele 1974, García-Madrigal 1994, Hendrickx 1996). Those inlets with highest values present intertidal pools, super-imposed flat rocks with light waves and also various kinds of algae that offer protection and food to crabs, as well as a wide variety of microhabitats. By contrast, inlets with lower densities showed a large amount of nearly smooth superimposed rocks that are constantly hit by strong waves and show a scarce plant cover. This is markedly perceivable at Grayson and North-W bays, where perturbations derived from the constant movement of round stones restricts the development of majids and favor species like P. transversus or C. explorator, which reach high densities under these conditions.
Most species show a clear relationship between abundance and the particular habitat type they inhabit (Crane 1947, Abele 1974, Villalobos-Hiriart et al. 1989, Hendrickx and Vázquez-Cureño, 1998). Thus, the Petrolisthes species better develop in areas with superim-posed rocks, relatively protected from strong waves. Particularly, P. edwarsii reached up to 556 org./100 m2 at North-W (50% of total abundance). By contrast, xanthids were more abundant at the lowest zone of rocks, among gravel, as in the case of X. cooksoni and some species of the genus Ozius that under these conditions recorded up to 414 and 210 org./100 m2 , respectively. Most majids species also show a marked preference for places protected by rocks or intertidal pools with abundant algae and organic matter, although they are always present at very low densities, as mentioned by Crane (1947) for several islands in the Pacific ocean and by García-Madrigal (1994) for the Maruata bay, Michoacan. However, Micropanope xantusii, mentioned as an inhabitant of coral reef exclusively (Garth 1960, 1961), was found under rocks among filamentous algae in this study. This close relationship between crab abundance and habitat availability was also observed at Blanca NE and North-W bays, where C. explorator showed the lowest densities due to absence of coral reef fragments and gravel under rocks, present at other bays where this hermit crab develops abundantly. Species with the lowest density varied in each bay according to their physiographic features.
Species recorded at all bays include P.transversu s, classified as an amphi-american tropical cosmopolitan species (Crane 1947, Martínez-Guzmán and Hernández-Aguilera 1993), as well as X. cookson i, mentioned as an endemic insular species (Garth 1992). G. grapsus and C. explorator were seen in all bays. The former is widely distributed in rocky shores of the Mexican Pacific coast (Crane 1947, Garth 1960, Villalobos-Hiriart et al. 1989, Hendrickx 1995a and b) and the latter constitutes a conspicuous element of the fauna in the Cabo San Lucas area, classified by Garth (1960) as having an insular character. Platypodiella rotundata, found under mid-sized rocks among gravel, has also been mentioned for Clarion (Hernández-Aguilera et al. 1986, Hernández-Aguilera and Martínez-Guzmán 1992), Maruata, Michoacan (García-Madrigal 1994), Clipperton (Hertlein and Emerson 1957, Garth 1965) and Galapagos islands, Ecuador (Villalobos-Hiriart et al. 1989). This is indicative of the wide distribution of these species, as well as of the fact that the composition of fauna in this island includes both continental and insular species.
The presence of Percnon gibbesi, Geograpsus lividus, Petrolisthes armatus and P. crenulatu s, both in northern and southern bays, could indicate that their distribution comprises the whole island with low densities, since they have been recorded along the Pacific coast from Baja California and the Gulf of California down to Chile and in Clarion, Maria Madre, Clipperton and Galapagos (Crane 1947, Hertlein and Emerson 1957, Garth 1960, Villalobos-Hiriart et al. 1989, Garth 1992, Hernández-Aguilera and Martínez-Guzmán 1992).
Socorro Island is added to the distribution range of species of Cataleptodius, Ozius, Herbstia and Acanthonyx, which were not considered by Garth (1992) despite their being found down to Chile and Panama. This omission could be due to the scarce information regarding littoral fauna in most parts of the island.
The list of species found in this work, compared with that of Clarion Island (Crane 1947, Garth 1961, Hernández-Aguilera et al. 1986, Garth 1992, Hernández-Aguilera and Martínez-Guzmán 1992), confirm that these islands share a large number of species: four grapsids, five xanthids, five majids, one diogenid and four porcellanids, most probably due to its closeness.
Llinas-Gutiérrez et al. (1993) mention that "the Revillagigedo Archipelago can be considered as a tropical zone with interannual variations that define it as a transition area; therefore, its fauna is constituted by tropical and temperate elements". According to this, it could be expected that the crab fauna of Socorro Islands littoral show the same type of components. However, the fauna is fundamentally Panamic (18 species), in addition to other four species (Thoe sulcata sulcata , Eucinetops rubellula, Petrolisthes armatus and P. crenulatus) that belong to the Mexican province defined by Briggs (1974), and two insular species, X. cooksoni and C. explorator (Garth 1992). These faunistic elements indicate that Socorro island is clearly an element of the Panamic province as indicated by Briggs (1974), where the temperate component mentioned by Llinas-Gutiérrez et al. (1993) is missing.
Composition and abundance of brachyurans and anomurans in the different bays indicate that there is a relative homogeneity throughout the area, with some differences derived from habitat complexity in some bays, as clearly reflected in the dendrogram (Fig. 2). Similarity between V. Lozano and North-E can be partially explained in terms of their similar physiographic features, with scarce intertidal pools and superimposed rocks with fissures and cavities and with a scarce plant cover, where the abundance of one or two species prevails. A similar situation is detected at Braithwaite and Blanca-SW, both having intertidal pools covered by algae that determine the occurrence of a higher crab diversity with a clear predominance of C. explorator over the remaining species. Binners bay was grouped with Grayson, even when their physiographic conditions differ, which can be explained as a result of the high densities that P. transversus showed compared to other species. The remaining bays are clearly separated, Blanca-NE because of the high majid diversity and North-W because its inhospitable conditions limit the occurrence of a high decapod diversity.
It is possible to state that the diversity of littoral crabs at Socorro Island varies along its rocky coast, depending on habitat complexity and resource availability. This becomes evident both in differences in species richness and in species relative abundance. Furthermore, variations through time in these parameters suggest the existence of a complex dynamics in crab community structure inhabiting the littoral fringe, influenced by variations in environmental conditions.
Acknowledgments
We wish to thank José L. Villalobos H. who provided literature and for his assistance in the species identification, and Francisco Cruz Vázquez for his assistance in collecting organisms and species identification. We thank anonymous reviewers for their valuable constructive comments and suggestions.
Resumen
Se estudió la comunidad de cangrejos del litoral rocoso de ocho bahías de la Isla Socorro, Archipiélago Revillagigedo en marzo de 1992. Las muestras fueron colectadas en cuadrantes de un m2 a lo largo de transectos perpendiculares a la costa en la zona intermareal. Se determinó la riqueza específica, la abundancia por especie para cada bahía, así como la similitud entre bahías mediante el índice de Morisita. Los organismos colectados pertenecen a seis familias, 20 géneros y 26 especies de los infraordenes Brachyura y Anomura. La familia con el mayor número de especies fue Xanthidae y la mayor riqueza específica se registró en la bahía V. Lozano. Pachygrapsus transversus, Xanthodius cooksoni y Calcinus explorator fueron las más abundantes en casi todas las localidades. En el dendrograma se definen dos agrupaciones con nivel de similitud superior a 0.67, dada la semejanza en la riqueza específica de cangrejos y las abundancias entre las bahías.
References
Abele, L.G. 1974. Species Diversity of Decapod Crustaceans in Marine Habitats. Ecology 55: 156-161. [ Links ]
Adem, J., E. Cobo, L. Blázquez, F. Miranda, A. Villalobos, T. Herrera, B. Villa & L. Vázquez. 1960. La Isla Socorro, Archipiélago de las Revillagigedo. Monog. Inst. Geofísica/2. UNAM. México. 234 p.
Birch, D. W. 1981. Dominance in Marine Ecosystems. Am. Nat. 118(2): 262-273. [ Links ]
Briggs, J.C. 1974. Marine Zoogeography. McGraw-Hill. New York. 473 p. [ Links ]
Brusca, R.C. 1980. Intertidal invertebrates of the Gulf of California, Univ. of Arizona. 513 p. [ Links ]
Crane, J. 1947. Eastern Pacific Expeditions of the New York Zoological Society. Intertidal Brachygnathous crabs from the West coast of tropical America with special reference to ecology. Zoologica. 22: 47-78.
García-Madrigal, M.S. 1994. Brachyura (Crustacea: Decapoda) de la Bahía de Maruata, Michoacán, México. Contr. Zool. Consultoría e Investigación Ambiental (1): 1-58. México.
Garth, J.S. 1958. Brachyura of the Pacific coast of America. Oxyrhyncha. Allan Hancock Pac. Exp. 21:1-854. [ Links ]
Garth, J.S. 1960. Distribution and Affinities of the Brachyuran Crustacea. In The biogeography of Baja California and adjacents seas. Part II. Marine biotas. Syst. Zool. 9:105-123. [ Links ]
Garth, J.S. 1961. Eastern Pacific Expeditions of the New York Zoological Society. XLV. Non-Intertidal Brachygnathous crabs from the West Coast of Tropical America. Part 2: Brachygnatha Brachyrhyncha. Zoologica: 46: 133-159.
Garth, J.S. 1965. The brachyuran decapod crustaceans of Clipperton Island. Proc. Calif. Acad. Sci, 33: 1-64. [ Links ]
Garth, J.S. 1992. The brachyuran crabs of the Revillagigedo Islands, Colima, México, with remarks on insular endemism in the Eastern Tropical Pacific. Proc. San Diego Soc. Nat. Hist. 24: 1-6.
Haig, J. 1960. The Porcellanidae (Crustacea, Anomura) of the eastern Pacific. Allan Hancock Pac. Exp. 24: 1-400. [ Links ]
Haig, J. 1966. Porcellanid crabs (Crustacea, Anomura) Results Scient. Comp. Calypso 7: 351-358. [ Links ]
Hendrickx, M.E. 1995. Checklist of brachyuran crabs (Crustacea: Decapoda) from the eastern tropical Pacific. Bull. Inst. Royal Scienc. Nat. De Belgique 65: 125-150. [ Links ]
Hendrickx, M.E. 1995. Cangrejos. pp. 565-636. In W. Fisher, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem (eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-Oriental. Vol. 1. Plantas e Invertebrados. Roma, Italia.
Hendrickx, M.E. 1996. Habitats and biodiversity of decapod crustaceans in the SE Gulf of California, Mexico. Rev. Biol. Trop. 44: 603-617. [ Links ]
Hendrickx, M.E. & L.A. Vázquez-Cureño. 1998. Composition and Zoogeographical Affinities of the Stomatopod and Decapod Crustacea Fauna Collected During the Ceemex P4 Cruise in the Gulf of Tehuantepec, México. Bull. Inst. Royal Scienc. Nat. De Belgique 68: 135-144. [ Links ]
Hernández-Aguilera, J.L., I. López-Salgado & P. Sosa-Hernández. 1986. Fauna carcinológica insular de México I. Crustáceos estomatópodos y decápodos de la Isla Clarión. Inv. Ocean. 3: 183-250. [ Links ]
Hernández-Aguilera, J.L. & L.A. Martínez-Guzmán. 1992. Notas acerca de la distribución de los estomatópodos y decápodos de aguas someras de Isla Clarión, Archipiélago Revillagigedo, Colima, México. Proc. San Diego Soc. Nat. Hist. 19: 1-6.
Hertlein, G.L. & W.K. Emerson 1957. Additional notes on the invertebrate fauna of Clipperton Island. Am. Mus. Nov. 1859: 1-9. [ Links ]
Holme, N. A. & A.D. McIntyre 1984. Methods for the Study of Marine Benthos. Blackwell, Oxford. 387 p. [ Links ]
Krebs, C. J. 1989. Ecological Methodology. Harper and Row. New York. 654 p. [ Links ]
Krebs, C. J. 1994. Ecology. The Experimental Analysis of Distribution and Abundance. Addison-Wesley. New York. 900 p. [ Links ]
Krohne, D.T. 1998. General Ecology. Wadsworth. New York. 722 p. [ Links ]
Ludwig, J.A. & J.F. Reynolds. 1988. Statistical Ecology. Wiley. New York. 337 p. [ Links ]
Llinas-Gutiérrez, J., D. Lluch-Cota, A. Castellanos, & A. Ortega-Rubio. 1993. La Isla Socorro, Revillagigedo, pp. 520-534. In S.I. Salazar-Vallejo and N.E. González (eds.). Biodiversidad marina y costera de México. Comisión Nacional de Biodiversidad y Centro de Investigaciones de Quintana Roo, México.
Martínez-Guzmán, L. & J.L. Hernández-Aguilera. 1993. Crustaceos Estomatópodos y Decápodos del Arrecife Alacrán, Yucatán, pp. 609-629. In S.I. Salazar-Vallejo and N.E. González (eds.). Biodiversidad Marina Costera de México. Comisión Nacional de Biodiversidad y Centro de Investigaciones de Quintana Roo, México. [ Links ]
Miles, P.M. & Miles H.B. 1966. Hultons Biological Field Studies Seashore Ecology. Hulton Educational Limited. London. 96 p. [ Links ]
Mille-Pagaza, S., A. Pérez-Chi & O. Holguin-Quiñones. 1994. Fauna malacológica bentónica del litoral de Isla Socorro, Revillagigedo, México. Cienc. Mar. 20: 467-486. [ Links ]
Raffaelli, D. & Hawkins S. 1997. Intertidal Ecology. Chapman and Hall. Great Britain. 356 p. [ Links ]
Rathbun, M.J. 1918. The Grapsoid crabs of America. Smithsonian Inst. U. S. Nat. Mus. 97: 461p.
Rathbun, M.J. 1925. Oxymatous and allied crabs of America. Smithsonian Inst. U.S. Nat. Mus. 129: 585 p.
Rathbun, M.J. 1930. The cancroid crabs of America. Smithsonian Inst. U.S. Nat. Mus.152: 609 p. [ Links ]
Ricketts, E.F., J. Calvin & J.W. Hedgpeth 1985. Between Pacific Tides. Stanford Univ. California. 614 p. [ Links ]
Sanders, H.L. 1968. Marine Benthic Diversity. A Comparative Study. Am. Nat. 102: 493-532. [ Links ]
Stiling, P. 1999. Ecology. Theories and Applications. Prentice Hall. New Jersey. 638 [ Links ]p.
Villalobos-Hiriart, J.L., J.C. Nates Rodríguez, A. Cantú Díaz Barriga, M.D. Valle Martínez, P. Flores Hernández, E. Lira Fernández & P. Schmidtsdorf Valencia. 1989. Listados Faunísticos de México I. Crustáceos estomatópodos y decápodos inter-mareales de las islas del Golfo de California, México. Inst. Biol. Univ. Nal. Autón. México, 114 p.
1 Departamento de Zoología, Lab. de Ecología. Escuela Nacional de Ciencias Biológicas. I. P. N. México, D. F. CP 11340 México. FAX (5) 396 35 03. aperezchi@hotmail.com

