










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.3-4 San José Dec. 2001
y su impacto sobre las hormigas en selvas del piedemonte
amazónico de Colombia
Tatiana Sanjuán1, Luis Guillermo Henao 2 y Germán Amat 1
2 Fundación Inguedé, Apartado Aéreo 41595, Bogotá, Colombia. Corel: inguede@andinet.com
Recibido 25-I-1991. Corregido 02-II-2001. Aceptado 08-II-2001.
Abstract
Entomopathogenic fungi of the genus Cordyceps were sampled in the foothills of the Colombian Amazon region (450 – 600 msnm altitude, January and May, 1998, i.e. dry and wet seasons, respectively). Healthy and Cordyceps-parasitized ants from a nearly pristine, a naturally disturbed and a human-disturbed forests were collected on 100 quadrants (1 m2) per forest. Litter, shrubs and trunks were explored as substrates, and samples were gathered in vertical strata from 50 cm to 2 m high. Aztec was the predominant ant genus in near pristine forest whereas Camponotus, Pheidole and Crematogaster were dominant in disturbed forests. In the naturally disturbed forest we found 759 individuals of Cordyceps unilateralis and 69 of C. lloydii var. binata parasitizing Camponotus spp (Formicidae: Formicinae), whereas C. kniphofioides var. ponerinarum and C. australis were found only in six cases infecting the ants Paraponera clavata and Pachycondila crassinoda (Formicidae: Ponerinae). In the human disturbed forest it were found 34 individuals of C. unilateralis on Camponotus. In the near pristine forest we did not find parasitized ants. The Camponotus spp ants parasitized by C. unilateralis and C. lloydii var binata were found predominantly in the substratum foliage up to 1 m. It was concluded that the incidence of Cordyceps–ant interactions is not influenced by the conservation status of the forest. Instead, relative humidity and presence/absence of host ants appear to be key factors. Also, the micro-spatial distribution of the interactions seems to follow a definite pattern. This work observations support the hypothesis on coevolution of dispersion mechanisms in Cordyceps and ant feromonal codes.
Key words: Cordyceps, ants, entomopathogenic fungi, tropical rain forest, biological control, Camponotus, fungal ecology
Cordyceps es el teleomorfo (forma sexual) de un hongo entomopatógeno perteneciente a la división Ascomycotina, parásito obligado de poblaciones de artrópodos y considerado como el más diverso del trópico. Junto a este hongo se han encontrado géneros relacionados como anamorfos (forma asexual) en los que se destacan Acremonium, Hirsutella, Hymenostilbe, Paecilomyces, Polycephalomyces, Sporothrix y Verticillium, los cuales presentan un gran pleomorfismo que les confiere una gran ventaja para dispersar esporas bajo una variedad de condiciones ambientales (Evans et al. 1988).
Cordyceps en hormigas tiene un ciclo de vida que se prolonga entre seis y 20 días, iniciándose con la adherencia de la espora a la cutícula. Una vez establecido el parásito, el individuo infectado presenta una considerable reducción de su actividad con desplazamientos asincrónicos y erráticos. En las siguientes 24 a 48 horas la hormiga muere y el hongo forma una estructura de resistencia, el esclerocio, en el celoma del insecto. Hacia la etapa final de la patogénesis, dependiendo de las condiciones ambientales, emerge el sinema del anamorfo o el estroma maduro de Cordyceps a través de las uniones del tórax con las extremidades y el pronoto (Evans 1982).
El primer ejemplar de Cordyceps fue descrito por Link en 1833, sobre una larva de Melolontha melolonthae (Coleoptera) (Mains 1958). Kobayasi ha realizado el tratamiento sistemático más completo sobre el género Cordyceps, dando a conocer una diversidad global de 232 especies (Kobayasi 1982), mientras Evans y Samson (1982, 1984) fueron los primeros en estudiar a Cordyceps en artrópodos de agroecosistemas de Ghana y selvas tropicales de Ecuador y Brasil encontrando seis especies de Cordyceps en hormigas. En la presente investigación se evaluó si el estado de conservación de los bosques estudiados es un factor determinante de la diversidad de Cordyceps, se evaluó su impacto en la mirmecofauna local y se identificó el patrón de distribución espacial de la interacción Cordyceps - hormiga.
Materiales y metodos
Zona de estudio: El muestreo se efectuó bajo la jurisdicción del municipio de Villagarzón, vereda Alto Chalhuayaco en el Piedemonte Putumayense (Amazonia colombiana), ubicado entre las estribaciones de la Cordillera Oriental y el lomerío amazónico a 0º50'54" latitud norte y 76º47'29.5" longitud oeste, en el piso térmico templado con altitudes entre 400 y 600 m y precipitaciones anuales entre 3800 y 4800 mm, de origen orográfico. El promedio de temperatura anual es 24 ºC y la humedad relativa posee valores hasta del 90 % correspondiente, según las Zonas de Vida de Holdridge, a "bosque muy húmedo tropical" (Bmh-T). Se estima que el Piedemonte Amazónico posee una de las mayores concentraciones de diversidad biótica del norte de Suramérica (Halffter 1992) y aun conserva un 80 % de su cobertura vegetal natural.
Selección y descripción de los bosques: Los muestreos se cumplieron en tres tipos de bosque, diferenciados principalmente por el grado de intervención, las condiciones microclimáticas, edáficas, fisiográficas, las especies dominantes, el estado de la hojarasca, el grado de epifitismo y el uso actual.
Recolección de hongos: En cada uno de los bosques se ubicaron 50 parcelas de 1 m2 al azar por época de muestreo. Se recolectaron hormigas parasitadas por hongos hasta 2 m por encima del suelo y 10 cm por debajo del suelo, revisando ramas, arbustos, troncos, hojarasca y el envés de las hojas.
Las muestras fueron descritas macroscópicamente y se fijaron con alfileres a cajas de cartón con icopor (polietileno) donde se secaron y se conservaron con gel de sílice. En el laboratorio, se describieron los ejemplares para la identificación taxonómica de las especies de Cordyceps. Los cuerpos fructíferos fueron rehidratados con KOH al 1 % y se realizaron cortes con tinción de azul de lactofenol para observación microscópica. El material recolectado está depositado en seco en la colección entomológica del Instituto de Ciencias Naturales (ICN-MHN), de la Universidad Nacional de Colombia, sede Bogotá.
La recolección de muestras micológicas se basó en los métodos empleados por Evans y Samson en Ghana, Brasil y Ecuador (Evans 1974, 1982).
Recolección de hormigas: La mirmecofauna se muestreo teniendo en cuenta la estratificación del bosque. En el estrato rasante se instalaron trampas de caída o "pitfall" (20 unidades), además del empleo de trampas de recolección de hojarasca o "winkler" y la recolección manual en troncos muertos y vivos. En el estrato herbáceo se empleó una red para captura de insectos y se hicieron 100 réplicas rozando superficialmente la vegetación en tres transectos lineales; complementariamente, se instalaron trampas "Malaise" (tres unidades por sitio). La mirmecofauna del estrato arbustivo / subarbóreo se recolectó mediante agitación foliar por un minuto con tres réplicas, además de recolecciones manuales en cortezas de árboles, inflorescencias e infrutescencias. La recolección de la mirmecofauna arbórea se llevo a cabo mediante trampas cebadas (40 unidades por sitio) que consistieron en frascos cilíndricos plásticos perforados, de 3 cm de longitud y 7 mm de diámetro, con carne de pescado en su interior. El material colectado se conserva en líquido (etanol al 70%) y está depositado en la colección entomológica del Instituto de Ciencias Naturales (ICN-MHN), de la Universidad Nacional de Colombia, sede Bogotá.
Estratificación: Durante la época seca se observó una tendencia de las diferentes especies de Cordyceps a ocupar un nicho particular en la estructura vertical del bosque con perturbación natural (B2), por lo que en el período lluvioso se procedió a cuantificar la cantidad de hormigas infectadas por este hongo. En cada unidad de muestreo las parcelas se estratificaron cada 50 cm en 2 m verticales. Los estratos y sustratos se clasificaron así:
Estratos: uno (0-0.50 m), dos (0.50-1 m), tres (1 - 1.5 m) y cuatro (1.5 - 2 m).
Sustratos: Hoja: Ápice o envés de plantas epífitas, folíolos de helecho o palma.
Raíces epífitas: Raíces aéreas de epífitas muy pequeñas sobre troncos y ramas de arbustos. Musgo: Briófitos epífitos que cubren la estructura de las plantas del sotobosque. Corteza: Tronco de árboles.
Los datos fueron sometidos a una prueba de bondad de ajuste de Kolmogorov para probar si existía un patrón determinado de distribución del hongo parasitando hormigas y con la prueba no paramétrica de Kolmogorov – Smirnov se trató de determinar si había alguna preferencia por estrato o sustrato por parte de las hormigas infectadas.
Resultados
Caracterización de los bosques: La consideración de ocho parámetros bióticos y abióticos permitió identificar marcadamente los tres tipos de bosque: con perturbación antrópica, con perturbación natural y poco perturbado (Cuadro 1).
Caracterización de los tipos de bosque de acuerdo con parámetros bióticos y abióticos
Table 1
Characterization of forest types according with biotic and abiotic parameters
| Parámetro | Bosque perturbado antrópicamente | Bosque con perturbación natural | Bosque poco perturbado |
| Grado de intervención | Alto. Causado por la tala de árboles y siembra de especies vegetales domesticadas. | Medio. Causado por la caída de arboles, flujo y reflujo del río y trocha humana. | Bajo. Causado por la caída de árboles y deslizamientos. |
| Topografía | Meseta ondulada, pendientes entre el 15-20 % | Meseta ondulada, pendientes menores del 10 %. | Meseta disectada, pendientes mayores del 40 %. |
| Nivel freático | Medio | Alto | Bajo |
| Humedad relativa | Oscilación diurna en 10 horas: 84-94 %. | Oscilación diurna en 10 horas: 90-98 % | Oscilación diurna en 10 horas: 80-94 %. |
| Temperatura | Oscilación diurna en 10 horas: 22-27 ºC. | Oscilación diurna en 10 horas: 23-25 ºC. | Oscilación diurna en 10 horas: 24-26 ºC. |
| Condiciones edáficas | Franco areno arcilloso. Niveles de materia orgánica bajos, con procesos de humificación y mineralización proporcionales. | Arcilloso. Niveles de materia orgánica altos, con procesos de humificación más que de mineralización. | Franco areno arcilloso. Niveles de materia orgánica bajos, con procesos de mineralización más que de humificación. |
| Especies vegetales dominantes | Cecropia ficifolia (Yarumo) Bactris gasipaes (chontaduro) Heliconia spp. (platanillos) | Cecropia sciadophylla (yarumo de monte) Socratea exhorriza (chonta) Heteropsis spruceana (yaré) | Socratea exorhiza (chonta) Attalea regia (bombona) |
| Grado de epifitismo | Medio. Está distribuido en su mayoría alrededor de las bases de los yarumos y árboles de mayor porte. Bejucos ausentes. | Alto. Se presentan en todos los estratos. Presencia de bejucos de bajo porte. | Bajo. Se presenta en árboles cercanos al nacimiento de la quebrada. Presencia de bejucos de porte grueso. |
| Característica de la hojarasca | Escasa. Capa de 0–3 cm de espesor. | Abundante. Capa de 0–20 cm de espesor. | Escasa. Capa de 0–3 cm de espesor. |
Especies de Cordyceps parásitas de hormigas: En la época seca se colectaron 207 ejemplares de hormigas parasitadas por hongos y en la época lluviosa 661 ejemplares. En ambos períodos el número total de especies de Cordyceps identificadas fue cuatro, además de siete anamorfos, también se encontraron dos especies de Torrubiella aun sin identificar (Cuadro 2).
Table 2
| Especies de Cordyceps | Anamorfos | Especies de hormigas |
| Cordyceps unilateralis (Tull.) Sacc. | Hirsutella formicarum | Camponotus sp3 Camponotus sp4 Crematogaster sp. Cephalotes atratus |
|
| Sporothrix insectorum | Camponotus sp3 |
|
| Tilachlidium liberianum | Camponotus sp3 Camponotus sp4 |
|
| Polycephalomyces sp. | Camponotus sp3 |
| Cordyceps lloydii var. binata (Faw.) |
| Camponotus sp3 |
| Cordyceps kniphofioides var. ponerinum (Evans y Samson.) | Hirsutella stilbelliformis var. ponerinum | Paraponera clavata |
| Cordyceps australis (Speg.) Sacc. |
| Paraponera clavata Pachycondyla crassinoda |
|
| Gibellula formicarum | Crematogaster sp. |
|
| Hymenostilbe sp. | Cephalotes atratus |
| Especies de Torrubiella |
|
|
| Torrubiella sp.1 |
| Camponotus sp3 |
| Torrubiella sp.2 |
| Camponotus sp3 Camponotus sp4 |
Evaluación de la mirmecofauna: En la región estudiada se presentó una riqueza de 28 géneros y aproximadamente 120 especies de hormigas (Cuadro 3). Los géneros predominantes por bosque de acuerdo con su abundancia relativa son: Azteca, en el bosque poco perturbado y que se encuentra ausente en los demás bosques; Camponotus, en el perturbado naturalmente y está asociado con Crematogaster en los estratos rasante, herbáceo y arbustivo. Crematogaster, Camponotus y Pheidole predominan en el bosque perturbado antrópicamente y son los taxones más abundantes representando más del 50% de la abundancia total de este sitio (Cuadro 3). De los 28 géneros presentes, Pachycondyla, Cephalotes, Paraponera, Crematogaster y Camponotus cuentan con especies parasitadas por Cordyceps o sus anamorfos o Torrubiella (Cuadro 2 y 3).
Géneros de hormigas presentes en los diferentes tipos de bosque del Piedemonte Amazónico colombiano y
su abundancia relativa (%)
Table 3
Genera of ants in different in forest types of the Amazonian Colombian foothill and their relative abundance (%)
| Bosque poco perturbado (B1) | Bosque perturbado naturalmente (B2) | Bosque perturbado antrópicamente (B3) | |||
| Azteca | 32 | Camponotus | 43 | Crematogaster | 25 |
| Pheidole | 9 | Crematogaster | 13 | Pheidole | 23 |
| Pachycondyla | 9 | Paratrechina | 13 | Camponotus | 11 |
| Cyphomyrmex | 7 | Solenopsis | 9 | Dolichoderus | 9 |
| Ectatomma | 7 | Linepithema | 7 | Acromyrmex | 8 |
| Crematogaster | 6 | Pheidole | 6 | Pachycondyla | 7 |
| Dolichoderus | 6 | Odontomachus | 4 | Paratrechina | 5 |
| Solenopsis | 6 | Megalomyrmex | 1 | Solenopsis | 4 |
| Paratrechina | 5 | Pachycondyla | 1 | Odontomachus | 3 |
| Gnamtogenys | 4 | Paraponera | 1 | Cyphomyrmex | 2 |
| Megalomyrmex | 3 | Cyphomyrmex | 1 | Ectatomma | 1 |
| Camponotus | 2 | Dolichoderus | 1 | Pseudomyrmex | 1 |
| Lepthotorax | 1 | Gnamtogenys | 1 | Wasmania | 1 |
| Pseudomyrmex | 1 | Cephalotes | 0 | Allomerus | 0 |
| Wasmania | 1 | Zacriptocerus | 0 | Dyscotherea | 0 |
| Zacriptocerus | 1 | Hypoponera | 0 | Gnamtogenys | 0 |
| Myrmelachista | 0 | Strugmigenys | 0 | Pathytireus | 0 |
|
|
|
|
| Tetramorium | 0 |
Los ceros son aproximados.
Presencia y ausencia de las especies de Cordyceps y géneros relacionados en bosques con diferentes regímenes de perturbación: Durante los muestreos realizados en época seca y lluviosa se registraron 868 hormigas parasitadas por hongos, de los cuales 834 casos se encontraron en 100 m2 del bosque perturbado naturalmente (B2), mientras el bosque perturbado antrópicamente (B3) tuvo 34 hormigas parasitadas en la misma área y el bosque menos perturbado (B1) no presentó hormigas parasitadas por algún hongo entomopatógeno (Cuadro 4).
en los dos períodos climáticos (B1, bosque poco perturbado; B2, perturbado
naturalmente y B3, perturbado antrópicamente)
Table 4
forest; B2, natural-disturbed; B3, human-disturbed)
| Especies de Cordyceps y anamorfos | Época seca | Época lluviosa | ||||
|
| B1 | B2 | B3 | B1 | B2 | B3 |
| Cordyceps unilateralis | - | +/127 | +/17 | - | +/537 | +/17 |
| Hirsutella formicarum | - | + | + | - | + | + |
| Sporothrix insetorum | - | + | - | - | + | - |
| Tilachlidium liberinum | - | + | - | - | + | + |
| Polycephalomyces sp. | - | + | - | - | + | - |
| Cordyceps kniphofioides var. ponerinum | - | +/2 | - | - | +/2 | - |
| Hirsutella stilbelliformis var. ponerinum | - | + | - | - | - | - |
| Cordyceps lloydii var. binata | - | +/34 | - | - | +/35 | - |
| Cordyceps australis | - | +/1 | - | - | +/1 | - |
| Especies de Torrubiella |
|
|
|
|
|
|
| Torrubiella sp. 1 | - | +/17 | - | - | +45 | - |
| Torrubiella sp. 2 | - | +/9 | - | - | +24 | + |
| TOTAL | 0 | 11/190 | 17 | 0 | 10/644 | 17 |
Número de presencias(+) o ausencias (-) / Número de hormigas parasitadas
Calculando índices de similaridad de presencia / ausencia para los dos bosques que presentan la interacción hongo entomopatógeno – hormiga se encontró que en la época seca, el bosque perturbado naturalmente (B2) es semejante al perturbado antrópicamente (B3) en un 14 %, mientras en temporada lluviosa es afín en un 50 %. Comparando el B2 en ambas épocas del año se halló una alta similaridad ( 88 %) , mientras que en B3 la similitud solo fué del 44 % (Cuadro 4). Esto demuestra que hay una clara diferencia espacio – temporal entre la composición de hongos del bosque perturbado naturalmente con el perturbado antrópicamente, pero en los mismos bosques en diferentes épocas del año existe una tendencia a mantenerse constante la composición de los hongos.
Estratificación vertical de la interacción Cordyceps / hormigas: La distribución de las hormigas muertas a causa de Cordyceps, en diferentes estratos y sustratos, responde a un patrón determinado (Kolmogorov p> 0.0.5). Se observó una tendencia de las hormigas atacadas por Cordyceps a ubicarse a menos de 50 cm de altura (estrato uno) y en el sustrato hoja (Cuadro 5). En cuanto a la distribución de las diferentes especies de Cordyceps en los estratos, se aprecia una tendencia de Cordyceps unilateralis y Cordyceps lloydii a estar a menos de 50 cm de altura (Cuadro 6). Con respecto a los sustratos, C. lloydii se localiza en hojas, mientras C. unilateralis tiende a ubicarse en los sustratos hoja y raicilla (Cuadro 6).
Hormigas infectadas por Cordyceps en los diferentes estratos y sustratos
Table 5
Ants infected for Cordyceps on the different strata and substrata
| Sustrato / Estrato | Hoja | Raicilla | Musgo | Corteza | Total |
| 1 (0.5 m) | 153 | 86 | 14 | 27 | 283 |
| 2 (1 m) | 79 | 48 | 14 | 5 | 146 |
| 3 (1.5 m) | 62 | 21 | 12 | 1 | 96 |
| 4 (2 m) | 85 | 20 | 12 | 2 | 119 |
| Total | 380 | 175 | 52 | 35 | ● 644 |
● Número de individuos
(K = 0.5131, P< 0.5**)
Distribución de las diferentes especies de Cordyceps en los distintos estratos y sustratos
Table 6
Species distribution of Cordyceps on the different strata and substrata
| Estrato | C. unilateralis | C. lloydii | C. australis | C. kniphofioides |
| 1 (0.5 m) | ● 261 | ● 18 | 1 | 1 |
| 2 (1 m) | 140 | 6 | 0 | 1 |
| 3 (1.5 m) | 93 | 4 | 0 | 0 |
| 4 (2 m) | 112 | 7 | 0 | 0 |
| Sustrato |
|
|
|
|
| Hoja | * 345 | 35 | 0 | 0 |
| Raicilla | 175 | 0 | 0 | 0 |
| Musgo | 52 | 0 | 0 | 0 |
| Corteza | 32 | 0 | 1 | 2 |
● (K-S = 0.08, P < .05).
* (X2 = 102.9, 3 g.l.,P < .001).
C. unilateralis (Cuadro 2) es la especie que posee más anamorfos y éstos se presentan en los diferentes estratos y sustratos. Las pruebas estadísticas señalan que las hormigas parasitadas con Hirsutella formicarum reiteran alturas aproximadas a 50 cm , mientras Sporothrix insectorum y Tilachlidium liberianum tienden a localizarse hasta 1 m de altura (estratos uno y dos) (Cuadro 7). De igual forma se analizó la disposición en los sustratos; existe una diferencia significativa (X2 = 81.199, P< 0.001), lo que indica que los anamorfos de C. unilateralis se ubican en un sustrato particular, especialmente H. formicarum, que se asocia a los sustratos raicilla y hoja y S. insectorum al sustrato hoja (Cuadro 7).
Distribución de los anamorfos de C. unilateralis en los diferentes estratos y sustratos
Table 7
Distribution of C. unilateralis anamorphs on diferent strata and substrata
Estrato | H. formicarum | S. insectorum | T. liberanium | Polycephalomyces sp. |
| 1 (0.5 m) | ● 102 | ● 34 | * 10 | 0 |
| 2 (1 m) | 38 | 43 | 6 | * 2 |
| 3 (1.5 m) | 8 | 31 | 8 | 0 |
| 4 (2 m) | 13 | 16 | 10 | 2 |
| Sustrato |
|
|
|
|
| Hoja | 36 | ◊ 89 | ◊ 24 | ◊ 4 |
| Raicilla | ◊ 78 | 24 | 5 | 0 |
| Musgo | 22 | 8 | 4 | 0 |
| Corteza | 25 | 3 | 1 | 0 |
* T. liberianum vs. Polycephalomyces (K-S = 0.1, P < 0.05)
◊ (X2 = 81.199, 3 g.l., P < 0.0011)
Discusión
Especies de Cordyceps del Piedemonte Amazónico: Las especies de este género, encontradas sobre hormigas del bosque muy húmedo tropical corresponden al menos a cuatro de las especies reportadas en el Neotrópico por Evans y Samson (1982, 1984) y Samson et al. (1982) en sus trabajos realizados en la zona del piedemonte amazónico ecuatoriano y en las selvas de Brasil, respectivamente. De acuerdo con las abundancias relativas de las diferentes especies, se corrobora que C. unilateralis predomina en hormigas del trópico en ambas épocas del año (Fig. 1); seguida por C. lloydii, Cordyceps kniphofioides y Cordyceps australis. Se desconoce por qué no se presentó C. doiana (Kob.) en hormigas de la subfamilia Ponerinae, especie ya registrada para Colombia y muy abundante en selvas con características similares a las del área de estudio (Kobayasi 1981).
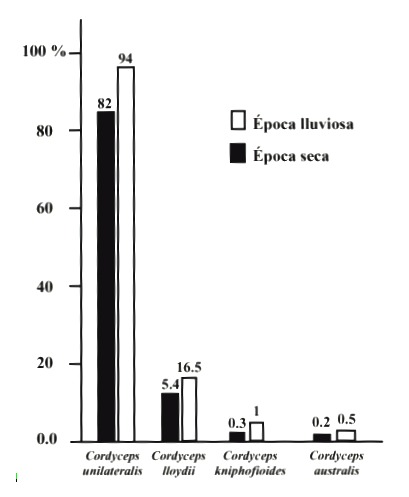
Fig. 1. Abundancias relativas de las especies de Cordyceps en los dos periodos de muestreo en el
piedemonte Amazónico Colombiano.
Fig. 1. Relative abundance of species of Cordyceps in the two sampling periods (wet and dry season)
in the Amazonian Colombian foothill.
Relación entre la mirmecofauna local y Cordyceps: La fauna de hormigas en la región estudiada presenta una riqueza genérica y específica alta si se tienen como referencia estudios locales en ambientes de selva húmeda tropical (Fernandez y Schneider 1989). La microdistribución y los altos valores de riqueza / abundancia observados en los bosques estudiados indican una riqueza trófica en el bosque con perturbación natural. La estructuración de las hormigas en grupos funcionales basada en la categorización de Andersen (1990) señala gremios especialistas como Azteca, oportunistas como Paratrechina, crípticos como Zacriptocerus y Wasmannnia y depredadores solitarias como Paraponera, Pachycondyla y Ectatomma. Esta categorización gremial refuerza la hipótesis de la saturación de nichos (Cuadro 3) (Andersen 1990).
La aparición de claros generados por la caída de árboles o la tala del bosque induce a la colonización de hábitats por especies generalistas como Dolichoderus, Camponotus, Crematogaster, Pheidole y Solenopsis (Cuadro 3). Esta situación de perturbación crea, en consecuencia, una confrontación entre individuos de estas especies por defender sus nidos; sí además son especies dominantes, entonces las especies competidoras que procuran los mismos recursos se convierten en el mayor regulador de sus poblaciones (Hölldobler & Wilson 1990).
Los géneros más abundantes y diversificados, que son Camponotus y Crematogaster (con seis y dos especies respectivamente), están clasificados funcionalmente como generalistas y dominantes (Hölldobler y Wilson 1990), lo que significa grupos de individuos abundantes, muy activos, con alta agresividad y capaces de explotar un alto número de recursos del ecosistema. Al parecer, ni Crematogaster ni Camponotus poseen especies antagonistas que las controlen, por ello se sugiere que la regulación demográfica podría explicarse en la interacción patógeno - hospedero. Por consiguiente, este sería un factor causal de la epizootia desencadenada por C. unilateralis, contrariamente a lo observado en el bosque menos perturbado, donde las poblaciones existentes de Camponotus a primera vista se mantienen dentro de su equilibrio biológico y no necesitan de un elemento regulador adicional.
Regulación de la interacción Cordyceps / hormigas y el estado de conservación del bosque: El haber encontrado un mayor número de especies de Cordyceps en bosques con algún grado de perturbación en selvas tropicales no coincide con los resultados obtenidos por Evans (1974) en Ghana, quien comparó la diversidad de hongos entomopatógenos de artrópodos en bosques primarios con condiciones similares al bosque poco disturbado (B1) de este trabajo; en este tipo de bosque el mismo autor encontró el mayor número de casos de infectividad de Cordyceps sobre hormigas. La razón de la abundancia de los hongos entomopatógenos en los bosques tropicales primarios es la existencia del denso dosel, lo que permite que el microclima se mantenga estable (Evans 1982, Evans & Samson 1984). A medida que el dosel desaparece, la radiación solar penetra con mayor intensidad ocasionando aumentos de temperatura y disminución de la humedad relativa que son los factores que permiten el éxito de la germinación de la espora del hongo (Evans, Samson & Latge 1988). Bajo esta hipótesis, se esperaría que el bosque menos perturbado (B1) tuviera la mayor abundancia de Cordyceps en hormigas, pero este no presentó ninguna hormiga parasitada. Es posible que las características de este bosque, con una pendiente mayor del 40 %, un drenaje rápido y un nivel freático profundo, propicien una humedad relativa muy variable y más baja que en los otros bosques (Cuadro 1). Por su parte, el bosque con intervención antrópica presenta la mayor temperatura diaria y una humedad relativa media; en estos hábitats se registraron 17 casos de Cordyceps parasitando hormigas, además de ortópteros y larvas de coleópteros. Es probable que el drenaje lento y el nivel freático medio, además de la características arcillosas del suelo, aumenten el régimen de humedad con relación al bosque más conservado (Cuadro 1). Regímenes microambientales como estos son altamente restrictivos para las poblaciones de hongos como Cordyceps.
En este trabajo, como en otros realizados con hormigas de la Amazonia (Andrade 1980, Evans y Samson 1984), se ha evidenciado una gran afinidad de C. unilateralis por Camponotus spp., en particular en aquellas especies que corresponden a bosques con algún nivel de perturbación (Evans 1974, Andrade, 1980). Las dos morfoespecies de Camponotus parasitadas por C. unilateralis (sp3 y sp4) sólo se hallaron en los bosques que presentaban algún régimen de disturbio ya sea natural o antrópico. Las morfoespecies que se colectaron en el bosque menos disturbado no se encontraron parasitadas en los otros bosques lo que significaría que la infectividad de Cordyceps es altamente selectiva. Al parecer el grado de perturbación del bosque no afecta directamente la diversidad de Cordyceps en hormigas, sino factores como la humedad relativa y la composición de la mirmecofauna, los cuales modularían la diversidad de la interacción (Fig. 2).
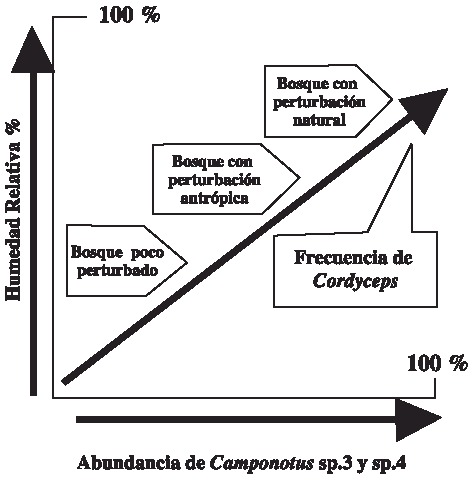
Fig. 2. Modelo explicativo de la interacción Cordyceps / hormiga y su relación con el estado de
conservación del bosque, la humedad relativa y la abundancia de la hormiga hospedera, Camponotus spp.
Fig.2. Model of Cordyceps / ant interaction, and conservation status of the forest, relative humidity and
the abundance of the host ant Camponotus spp.
Patrones de distribución espacial de Cordyceps y sus anamorfos en el bosque con perturbación natural: La distribución de las interacciones a nivel de estrato y sustrato es muy localizada. Las conductas de los individuos infectados sugieren la presencia de hormonas (de agonía) que pueden originar desplazamientos masivos y recurrentes, este es un factor que puede explicar la disposición gregaria de hormigas infectadas por Cordyceps.
Las hormigas infectadas por C. unilateralis se encontraron asociadas a los sustratos hoja y raicilla a bajas alturas (entre 0 y 1m) de tal forma que el estroma del hongo puede dispersar sus ascósporas libremente sobre el suelo o sobre hormigas que por allí pasen. Hormigas del género Camponotus, infectadas por C. unilateralis, fueron encontradas en Brasil adheridas al envés de las hojas de hierbas y arbustos y escondidas siempre de la luz solar directa (Evans 1982, Evans y Samson 1982, 1984). Al parecer este funcionamiento está asociado a patrones altruistas de cada especie de Camponotus, expresados mediante el traslado de cadáveres lejos de los centros de actividad de las obreras, los que posteriormente son devorados por sus congéneres antes de la esporulación del hongo. De otra parte, en Ghana, Evans (1974) halló especies de Camponotus infectadas que se exponían al sol ubicándose en ramas y en el haz de las hojas como respuesta al comportamiento fotofílico del hongo. Muchas especulaciones al respecto se han dado; no obstante, para la mayoría de autores este comportamiento responde a las toxinas del hongo que logra producir comportamientos anormales dentro de la hormiga (K. Jaffe com. pers.).
Los anamorfos de C. unilateralis, al igual que su forma sexual, se asocian a Camponotus ubicados a ciertas alturas y sustratos de acuerdo con sus mecanismos de dispersión de esporas (Cuadro 7). Los individuos infectados con H. formicarum, por ejemplo, se ubicaron en raicillas y hojas a una altura de 50 cm; este patrón quizás se debe a la menor incidencia de luz a esta altura y porque sus esporas mucilaginosas podrían desplazarse mejor a través del agua. S. insectorum alcanza a ubicarse hasta un metro de altura asociado al envés de las hojas; debido a su resistencia a la desecación y a la capacidad que posee de dispersar esporas a través del aire. Mientras T. liberianum presenta una inclinación por alturas menores a un metro, pero las observaciones obtenidas no permiten concluir si existe un sustrato específico para este anamorfo.
Cordyceps lloydii var binata, se halló siempre en el envés de hojas de consistencia suculenta, de tal manera que se le facilitó la dispersión de las esporas, la protección contra la desequedad y el establecimiento del inóculo (Cuadro 6). Las especies de Cordyceps encontradas sobre hormigas de la subfamilia Ponerinae (Cuadro 2 y 6) se hallaron en el suelo o adheridas a la corteza de bejucos, como el "Yaré" (Heteropsis spruceana) y otras lianas; contrariamente a lo que describen Evans y Samson (1982) quienes sostienen que las ponerines Neotropicales siempre se esconden bajo la hojarasca y que sólo el estroma de Cordyceps emerge como respuesta a un estimulo fotofílico . En los ejemplares del Putumayo el ascenso de estas hormigas enfermas a la parte alta del "Yaré" se explica por que este hábito está ligado al comportamiento de hormigas solitarias como las "congas" (Paraponera clavata) que forrajean en el suelo y utilizan estos bejucos para ascender a nectarios florales, pero en el recorrido fallecen y terminan adheridas al bejuco o la corteza del árbol donde emerge el estroma de Cordyceps buscando la luz solar (Belk et al. 1989).
Es de interés anotar la constancia numérica de los individuos de Camponotus sp4 infectados por C. unilateralis y sus anamorfos durante las dos épocas de muestreo en el bosque con perturbación antrópica. En ambos períodos se recogieron en un mismo tronco entre 17 y 20 ejemplares. No se observó en ningún caso un centro de actividad de las hormigas o el nido. Un caso similar se presentó en Brasil, pero con Cephalotes atratus y Cordyceps kniphofioides que aparecieron en un mismo árbol durante un intervalo de muestreo de dos años (Evans y Samson 1982).
Finalmente, la disposición de los hongos en los diferentes sustratos y estratos permite postular que los mecanismos de dispersión de Cordyceps coevolucionaron con el comportamiento de las hormigas hospederas y es muy posible que códigos feromonales puedan regir estos complejos comportamientos inducidos por las toxinas de Cordyceps y el patrón altruista de las hormigas.
Agradecimientos
Este trabajo fue llevado a cabo gracias a la financiación del Fondo para la Protección del Medio Ambiente FEN Colombia y al apoyo del Instituto de Ciencias Naturales y el Departamento de Biología de la Universidad Nacional de Colombia. Agradecemos a la comunidad indígena Inga residente en la vereda "Alto Chalhuayaco" (Putumayo) y a la Corporación Autónoma para el Desarrollo Sostenible Sur de la Amazonia (CORPOAMAZONIA) por su apoyo logístico. Finalmente, un reconocimiento a Nivea Cristina Garzón por la lectura y sugerencias al manuscrito.
Resumen
Se estudió la diversidad del género Cordyceps, hongo entomopatógeno, en hormigas del piedemonte Amazónico colombiano (450-600 msnm, enero y mayo de 1998, épocas seca y lluviosa, respectivamente). La recolección de hormigas sanas y parasitadas por Cordyceps se llevó a cabo en tres bosques con distintos regímenes de perturbación: natural, antrópico y poco perturbado. Se establecieron 100 cuadrantes de 1 m2 por cada bosque. Las muestras fueron tomadas en hojarasca, arbustos y troncos como sustratos, y cada 50 cm hasta 2 m de altura los estratos verticales. El género Azteca fue el predominante en el bosque poco perturbado, mientras Camponotus, Pheidole y Crematogaster abundaron en los bosques con perturbación natural y antrópica. El bosque con perturbación natural presentó 759 individuos de C. unilateralis y 69 de C. lloydii var. binata parasitando hormigas Camponotus spp. (Formicidae: Formicinae). Se encontraron sólo seis casos de C. kniphofioides var. ponerinarum y C. australis en las hormigas Paraponera clavata y Pachycondyla crassinoda (Formicidae: Ponerinae). En el bosque perturbado antrópicamente se presentaron 34 ejemplares de C. unilateralis en Camponotus mientras que en el menos perturbado no se observaron hormigas parasitadas. Se halló que las hormigas Camponotus spp. que son parasitadas por C. unilateralis y C. lloydii var. binata se ubican preferencialmente en el sustrato hoja, hasta 1 m de altura. Se determinó que la incidencia de la interacción Cordyceps / hormiga no está influenciada por el estado de conservación del bosque, sin embargo, la variación de la humedad relativa y la presencia o ausencia de la hormiga hospedera son factores que influyen en la diversidad de Cordyceps en hormigas. Se encontró, además, que la distribución microespacial de la interacción sigue un patrón determinado, aportando más argumentos a la hipótesis de que los mecanismos de dispersión de Cordyceps coevolucionaron con la hormiga influenciando sus códigos feromonales.
Referencias
Andersen, A. N. 1990. The use of ant communities to evaluate change in Australian terrestrial ecosystems: a review and a recipe. Proc. Ecol. Soc. Aust. 16: 347-357. [ Links ]
Andrade, C. de. 1980. Epizootia natural causada por Cordyceps unilateralis (Hypocreales, Euascomycetes) em adultos de Camponotus sp. (Hymenoptera, Formicidae) na região de Manaus, Amazonas, Brasil. Acta Amazonica 10: 671-677. [ Links ]
Belk, M., H. Black & C. Jorgensen. 1989. Nest tree selective by tropical ant, Paraponera clavata. Biotropica 21: 173-177. [ Links ]
Evans, C. H. 1974. Natural control of arthropods, with special reference to ants (Formicidae) by fungi in the tropical high forest of Ghana. J. Appl. Ecol. 11:37-49. [ Links ]
Evans, C. H. 1982. Entomogenous fungi in tropical forest ecosystems: an appraisal. Ecol. Entomol. 7:47-60. [ Links ]
Evans, C. H. & R. A. Samson. 1982. Cordyceps species and their anamorphs pathogenic on ants in tropical forest ecosystems: I. The Cephalotes complex. Trans. Br. Mycol. Soc. 79: 431-453. [ Links ]
Evans, C. H. & R. A. Samson. 1984. Cordyceps species and their anamorphs pathogenic of ants (Formicidae) in tropical forest ecosystems II. The Camponotus (Formicinae) complex. Trans. Br. Myco. Soc. 82: 127-150. [ Links ]
Evans, C. H., R. A. Samson & J-P. Latge. 1988. Atlas of entomopathogenic fungi. Springer, Berlín. 186 p. [ Links ]
Fernandez, F. & L. Schneider. 1989. Las hormigas de la reserva natural de la Macarena. Rev. Col. Entomol. 15: 38-44. [ Links ]
Halffter, G. 1992. La diversidad biológica de Iberoamérica. Instituto de Ecología, México D. F. 388 p. [ Links ]
Holdobler, B. & F. O. Wilson. 1990. The ants. Harvard University, Boston. 896 p. [ Links ]
Kobayasi, Y. 1981. Revision of the genus Cordyceps and its allies 2. Bull. Natur. Muse. Tokyo Ser. B. 7: 123-129. [ Links ]
Kobayasi, Y. 1982. Keys to the taxa of the genera Cordyceps and Torrubiella. Trans. Mycol. Soc. Japan 23: 329-364. [ Links ]
Mains, E. B. 1958. North American entomogenous species of Cordyceps. Mycol. 50 :169-222. [ Links ]
Samson, R., H. Evans & S. Hoekstra. 1982. Notes of entomopathogenous fungi from Ghana. VI. The genus Cordyceps. Proc. Neth. Akad. Wetenscha., Ser. C85: 589-605. [ Links ]

