











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.49 no.3-4 San José dic. 2001
(Perciformes: Coryphaenidae), del litoral de Sinaloa, Nayarit
y Baja California Sur, México
J.V. Madrid y R. Beltrán-Pimienta.
Instituto Nacional de la Pesca. Centro Regional de Investigación Pesquera de Mazatlán, Sinaloa, México. A.P. 711. Mazatlán, Sinaloa, México, Tel. 5269880049, fax 5269880002. juanchomvera@yahoo.com
Recibido 23-III-2001. Corregido 20-III-2001. Aceptado 6-IV-2001.
Abstract
Dolphin fish population data were recorded from landings in Sinaloa, Nayarit and Baja Califonia Sur, Mexico, in 1997. Of 3 211 organisms only 2 812 produced complete biometric data. The maximal length and weight were 192 cm and 30 Kg. The three sampling sites and the climatic periods also were associated with different size (P < 0.05). The covariate analysis of weight and length relationships indicated significative differences (P < 0.05) for total data and by sex. Condition factor was higher throughout the study period in Mazatlán, and the variation was higher, in the spring. The sexual ratio was in Mazatlán was constant, while there were more males in Nayarit in the fall and the spring and in Los Cabos in the spring. There may be population differences, between the central mexican Pacific Coast and the Peninsula of Baja data.
Key words: Dolphin fish, population comparison, sizes, Gulf of Califonia
Introducción
Un problema en el estudio de peces pelágicos, es la incertidumbre sobre los límites en la distribución de las diferentes poblaciones. Es común dar por hecho que las capturas de una misma especie en lugares distantes, corresponde a la misma población, y las diferencias en las tallas, reclutamientos, madurez y desoves son atribuidas a las variaciones en la temperatura, surgencias o corrientes marinas y a las interacciones de éstas con la disponibilidad de alimento, el crecimiento y la mortalidad. Sin embargo las diferencias podrían estar relacionadas a la existencia de diferentes poblaciones de la misma especie (Oxenford y Hunte 1986, Mahon y Hunte 1999).
Además de las dificultades para definir los límites de las poblaciones, existe en el Pacífico mexicano, una pesquería artesanal del dorado al margen de la ley, que no permite tener registros sistemáticos de las capturas, pues Coryphaena hippurus esta reservado a la pesca deportiva en México (Anónimo 1992). Las capturas anuales artesanales llegan a representar hasta el 55% del total de las obtenidas por la flota artesanal en el litoral Pacífico central mexicano (Beltrán et al. 1995).
Considerando la importancia que tiene este recurso pesquero y para contribuir al conocimiento de la estructura y la dinámica poblacional del dorado (C. hippurus), que habitan en la región de la boca del Golfo de California, nuestros objetivos fueron: comparar las frecuencias de tallas, las relaciones entre el peso y la longitud, los factores de condición y las proporciones sexuales, utilizando el enfoque de analizar todos los aspectos posibles del ciclo de vida; esta estrategia ha sido empleada para las poblaciones de dorados del Atlántico (Oxenford y Hunte 1986).
Materiales y métodos
Los datos provienen de capturas con línea de mano durante 1997 y desembarcadas en Playa Sur, Mazatlán, La Peñita de Jaltemba en Nayarit y Los Cabos en Baja California Sur, México. Los muestreos en Los Cabos, fueron tomados de las capturas de la flota deportiva. Los muestreos de Nayarit y Sinaloa, se realizaron de las capturas con embarcaciones menores que operan en dichos lugares. Los sistemas de captura son semejantes
De cada organismo se registró la longitud furcal y el peso eviscerado, a partir de éstos se calcularon los máximos, los promedios y los mínimos. El sexo se determinó con base al dimorfismo sexual que presenta la especie.
Los datos se agruparon en periodos climáticos (Fiedler 1992 y Fiedler et al. 1992): de enero a marzo, el invierno; de abril a junio, primavera; de julio a septiembre, el verano y de octubre a diciembre, el otoño.
A partir de los datos agrupados, se calcularon las relaciones peso y longitud (transformados a ln x+1), por medio de correlación lineal, las que fueron posteriormente comparadas por un análisis de covarianza. Para calcular el factor de condición, se utilizó el modelo de
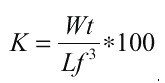
adonde Wt = peso total y Lf = longitud furcal.
Los intervalos de clase de las frecuencias de longitudes (1 cm) organizadas por periodos climáticos, zonas de estudio y totales, fueron comparados con la prueba de Kruskal-Wallis. Estos intervalos, también fueron comparados mediante un análisis de varianza de Friedman, intercambiable con X 2. Una prueba no-paramétrica del Signo, cuya significancia se alcanza con el 50% de probabilidad, para la hipótesis de nulidad, fue utilizada para comparar zonas.
Área de estudio: La zona de estudio se encuentra ubicada en la entrada del Golfo de California, (entre 105° 15' a 110° 05' W y 21 a 23° 30' N). La región se encuentra bajo la influencia de las corrientes marinas de California, la Corriente Norecuatorial, la Contracorriente Norecuatorial y la Corriente Costera de Costa Rica (Baugmarter y Christensen 1985, Xie y Hsieh 1995, Badan 1997). Climáticamente esta ubicada en la transición de la zona tropical y subtropical.
Resultados
El número de organismos analizados fue de 3 211. Solo en 2 812 se registraron datos de peso, longitud y sexo simultáneamente (Cuadro 1). La composición de las longitudes y pesos máximos, medias y mínimos se pueden observar en el Cuadro 1, donde además se presentan las proporciones sexuales por zona.
La mayor longitud furcal fue de 192 cm y 30 kg. en peso, tal como se puede observar en los datos del otoño, en Los Cabos, Baja California Sur (Cuadro 1). La menor talla fue de 40 cm y el menor peso 0.5 kg., en verano y primavera simultáneamente, en Mazatlán, Sinaloa.
mexicana, durante el año de 1997
Table 1
Mexican coast during 1997
| N | Lmáx | Lmedia | Lmín | Wmáx | Wmedio | Wmín | ♀:♂ | |
| Mazatlán | ||||||||
| Primavera | 919 | 129 | 72.2 | 45 | 17.0 | 3.3 | 0.5 | 0.45: 0.55 |
| Verano | 376 | 128 | 74.5 | 40 | 19.0 | 3.8 | 1.0 | 0.44: 0.56 |
| Otoño | 114 | 90 | 52.7 | 45 | 7.0 | 1.6 | 1.0 | 0.45: 0.55 |
| Nayarit | ||||||||
| Primavera | 118 | 103 | 68.9 | 47 | 6.0 | 2.7 | 0.85 | 0.39: 0.61 |
| Verano | 85 | 139 | 61.9 | 40 | 20.0 | 2.3 | 0.70 | 0.37: 0.63 |
| Otoño | 135 | 140 | 95.7 | 54 | 21.0 | 6.7 | 1.10 | 0.56: 0.44 |
| B. Cal. Sur | ||||||||
| Primavera | 414 | 137 | 97.3 | 47 | 23.7 | 8.9 | 1.0 | 0.64: 0.36 |
| Verano | 159 | 140 | 88.0 | 65 | 23.5 | 6.4 | 2.5 | 0.40: 0.60 |
| Otoño | 492 | 192 | 89.4 | 58 | 30.0 | 6.2 | 1.5 | 0.37: 0.63 |
N = es el número de organismos, Lmáx = es la longitud furcal máxima, Lmedia = es la longitud furcal media y Lmin = es la longitud furcal mínima, W = se refiere al peso y los subíndices son máximos, medios y mínimos respectivamente. La columna final son las proporciones sexuales. Las longitudes están en cm y el peso en Kg.
N = number of organism, Lmáx = maximum furcal length, Lmedia = median furcal length, Lminimum = furcal length, W = weight, subindexes ara maximum, median and minimum respectively. The last column are the sexual proportions. Lengths are in cm and weight in kg.
Las proporciones sexuales se mantuvieron constantes en Mazatlán. En La Peñita de Jaltemba en Nayarit, siempre hubo más hembras que machos, pero en el otoño fue al contrario. En los Cabos, Baja California Sur, hubo cerca del doble de machos en la primavera y en el otoño hubo casi el doble de hembras.
Los tres sitios difieren en la estructura de tallas (Kruskal-Wallis, P < 0.05, Fig. 1), al igual que las estaciones climáticas (Kruskal-Wallis, P < 0.001, Fig. 2). En cada sitio la comparación múltiple no fue significativa.
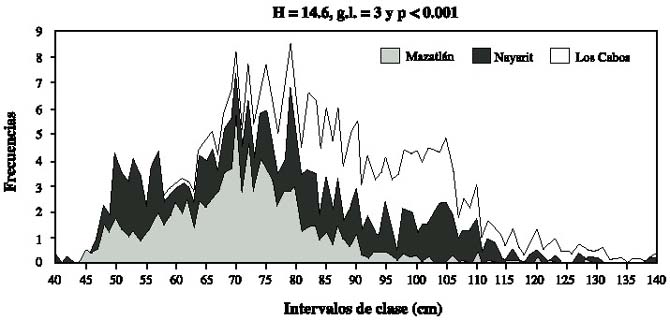
la costa noroccidental mexicana,durante el año de 1997.
Fig.1.Histogram of the percentual frequency of the lenght in 1 cm intervals of Coryphaena hippurus from the northwest
Mexican coast during 1997.
El análisis de varianza de dos-vías de Friedman (equivalente a X²), para los intervalos de frecuencias de tallas de un cm y de los 3 sitios, también produjeron diferencias significativas (X² = 8.5, N = 161, g.l.= 2 y P < 0.013). Tampoco se detectaron diferencias en la comparación múltiple.
Se observaron diferencias significativas, para la prueba del signo, entre Mazatlán vs Los Cabos (Z = 2.19, P < 0.028), y Nayarit vs Los Cabos (Z = 2.27, P < 0.024), pero no entre las poblaciones de Nayarit y Mazatlán (Z = 0.75, P < 0.45).
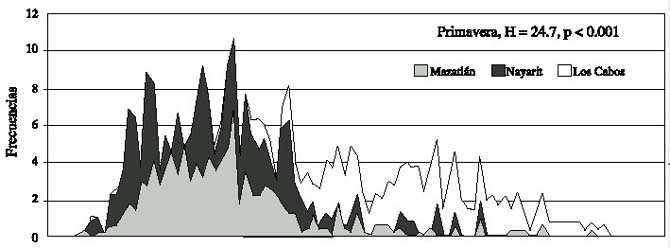
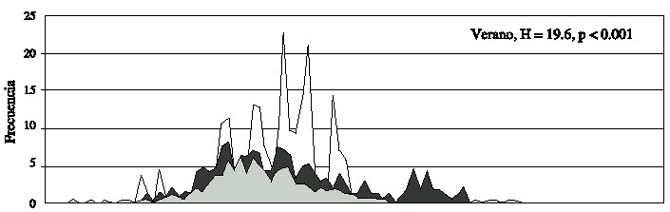
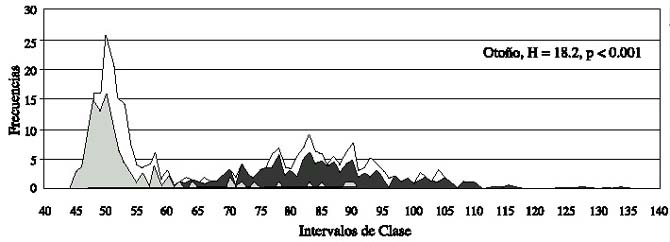
Fig.2.Histograms of the frequency distribution of the length for the three study areas,grouped by climatic period during 1997.
El análisis de covarianza de las relaciones entre el peso y la longitud, para los totales de las tres zonas de estudio, indica diferencias significativas (F = 765, P < 0.001, Fig. 3). Tampoco se pudo constatar, por comparacion múltiple diferencias entre sitios. Las elevaciones no presentaron diferencias significativas.
Esta misma prueba, produjo en la primavera de 1997 diferencias significativas en los tres sitios de muestreo para las pendientes (F = 196, P < 0.001) y para las elevaciones (F = 25, P < 0.001), por lo cual se rechaza en ambos casos las hipótesis de nulidad, asimismo las comparaciones múltiples, no detectaron diferencias significativas. Para el verano, las tres zonas de muestreo también presentaron diferencias significativas para las pendientes (F = 126, P < 0.001) y para las elevaciones (F = 352, P < 0.001), asimismo, las comparaciones múltiples no detectaron diferencias significativas, a pesar de los valores de F.
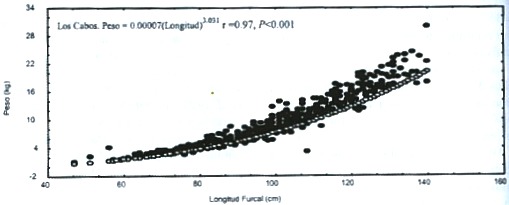
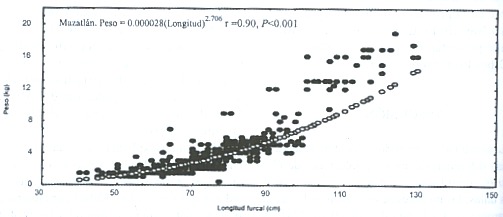
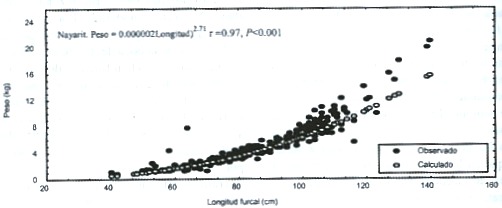
Fig.3.Weight and length relationship of both sexes of Coryphaena hippurus for each study area in the vestibular region of
the Gulf of California.
Las relaciones de peso y longitud de los machos de las tres zonas, también presentaron diferencias significativas para las pendientes (F = 402, P < 0.001), pero no fueron significativas entre las elevaciones (F = 1.4E-11, P > 0.05). Para las hembras de las tres zonas también presentaron diferencias significativas para las pendientes (F = 455, P < 0.001) y tampoco fueron significativas las elevaciones (F = 1E-11, P > 0.05).
Para el factor de condición (Cuadro 2), se puede observar que los mayores valores se presentaron en la primavera, en las zonas de Mazatlán y Nayarit. En términos globales en Mazatlán se presentaron los mayores valores y en Los Cabos los menores valores.
noroccidental mexicana (Sinaloa, Nayarit y Baja California Sur)
y para tres épocas climáticas de 1997
Table 2
Condition factors for Coryphaena hippurus in three study areas in the northwest Mexican
coast (Sinaloa, Nayarit and Baja California Sur)
and for three climatic season of 1997
| Zona de estudios | Primavera | Verano | Otoño |
| Los Cavos, B.C.Sur | 1.32 + 0.17 | 0.49 + 0.05 | 0.90 + 0.09 |
| Mazatlán | 6.82 + 2.05 | 2.55 + 0.59 | 3.38 + 0.46 |
| Nayarit | 5.25 + 0.98 | 1.33 + 0.09 | 2.54 + 0.53 |
Discusión
Los resultados obtenidos permiten observar que hay diferencias entre las poblaciones de C. hippurus, como las que se pueden distinguir entre las tallas; las menores se presentan en los desembarques de La Peñita de Jaltemba y Mazatlán y los organismos mayores se presentan en Los Cabos. Estas diferencias son evidentes en la comparación de las distribuciones de frecuencia del verano (Fig. 1). Otras diferencias cualitativas se observan entre las proporciones de machos y hembras durante los tres periodos climáticos de 1997; en la zona de Mazatlán la proporción de hembras es casi constante y en Nayarit, es mayor la presencia de hembras. En el otoño en Nayarit, los machos son más numerosos que las hembras. En los Cabos las proporciones tuvieron mayor variación que en los otros sitios: en la primavera el número de machos fue casi el doble que el de hembras, invirtiéndose durante el verano. En el otoño nuevamente hay más hembras.
Diferencias significativas aparecieron en las pruebas pareadas de la estructura de tallas, lo que permite separar las poblaciones de los Cabos, de las de Nayarit y Mazatlán.
Diferencias significativas también aparecieron entre las relaciones del peso y la longitud furcal, las que pueden estar asociadas a la disponibilidad de recursos alimenticios y a la estructura de las poblaciones. Estas diferencias se presentan entre los sexos de esta especie (Massuti et al, 1999), que puede estar asociadas a las eficiencias para conseguir alimento.
Con respecto al factor de condición, las poblaciones tuvieron valores altos durante la primavera. En los resultados de Mazatlán, las varianzas son suficientemente grandes y pueden ser un indicador de la madurez gonádica durante este periodo, aunque se ha señalado actividad reproductiva durante todo el año, tanto en la zona de Los Cabos (García-Melgar 1995), como en Mazatlán (Barcelata–Ortiz 1990), lo que habría que revisar. Diferencias en los patrones de reproducción (diferencias en fecundidad y tamaño de huevos), han sido documentados para las poblaciones de dorado del norte y del sur del Atlántico Central Oeste (Oxenford y Hunte 1986, Oxenford 1999)
Buscando explicaciones a las diferencias encontradas, podemos observar que las tres áreas geográficas poseen características particulares. La zona de Los Cabos por ejemplo, carece de plataforma continental y las profundidades de 500 m son muy cercanas a la costa (Abítia-Cárdenas 1992). En la zona de Mazatlán y Nayarit es moderada. Por otro lado las masas de aguas superficiales y subsuperficales de la región de la boca del Golfo de California, se encuentran bajo la influencia de las corrientes marinas de California, la Corriente Norecuatorial, la Contracorriente Norecuatorial y la Corriente Costera de Costa Rica (Baugmarter y Christensen 1985, Xie y Hsieh 1995, Badan 1997), estas influencias varían en el tiempo; por ejemplo Los cabos queda separado oceanográficamente durante los inviernos y los años extraordinariamente fríos. La influencia del verano y el Niño-oscilación del Sur (Wyrtki 1966, Fiedler et al. 1992; Fiedler 1992, Fiedler et al. 1992, Ramp et al. 1997, Anónimo 1999), rompería constantemente estas separaciones. Santana-Hernández (2001), en su estudio sobre las abundancias de dorado, en las capturas de barcos palangreros, ha demostrado que las poblaciones de Los Cabos quedan relativamente aisladas (véase también Norton 1999, para los movimientos asociados a la Corriente de California).
Bajo la visión darwiniana las especies se encuentran formadas por poblaciones físicamente separadas, cuya cohesión se mantiene atravez de la migración. La idea dominante es suponer que la distribución es contínua, sin embargo muchas poblaciones sons discontínuas y se distribuyen en parches, manteniendo el flujo de genes, por medio de larvas, huevos o adultos, de manera discreta y extraordinaria, tal como se ha documentado para cambios de distribución relacionados a los eventos El Niño (Hubbs 1948, Walker 1966, Allen y Robertson 1994). La propuesta de discontinuidades en las poblaciones de dorado, ha sido desarrollada para el Atlántico Central (Oxenford y Hunte 1986) y apoyan a las diferencias que hemos observado. En las aguas del Mar de Japón, Kojima (1965), señala que los dorados presentan migraciones que pueden significar separaciones poblacionales. Por otra parte, se ha comentado que los cambios en la estacionalidad de las capturas en aguas internacionales, sugieren la posibilidad de que existan dos poblaciones, una que habita en el Pacífico Norte y la otra en el hemisferio sur (Kojima 1964, Patterson y Martinez 1991; Norton y Crooke, 1994)
Por lo anterior nuestros datos nos indican diferencias poblacionales en la región, en las que habría que profundizar por las implicaciones que tiene en el manejo futuro de este recurso tan importante en las costas del Pacífico central mexicano.
Resumen
Poblaciones de dorado (Pisces) fueron muestreadas en Sinaloa, Nayarit y Baja Califonia Sur, México, en 1997. Unos 3211 organismos fueron analizados. Las tallas máximas fueron de 192 cm y 30 Kg. Hay diferentes tallas asociadas a las zonas y épocas climática (P < 0.05). Las relaciones del peso y la longitud son diferentes (P < 0.05), para totales y sexos. El factor de condición fue alto en Mazatlán y hay gran variación en primavera. La proporción sexual fue constante en Mazatlán, y hubo mayoría de machos en Nayarit, en otoño y primavera y en primavera en Los cabos. Pueden existir diferencias entre poblaciones costeras del Pacífico y Baja California.
Palabras claves: Dorado, comparación poblacional, tallas, Golfo de California.
Referencias
Abítia-Cárdenas, L. A. 1992. Espectro trófico energético del marlin rayado Tetrapturus audax (Phillippi, 1887) y marlin azul Makaira mazara (Judan y Saydor, 1902) de la zona de Cabo San Lucas, Baja California Sur, México. Tesis de Maestría. Centro de Investigación en Ciencias marinas- Instituto. Politécnico Nacional, 76 p. [ Links ]
Allen, G. R. & R. D. Robertson. 1994. Fishes of the Tropical Eastern Pacific. Crawford House, Bathurst, Australia, 332 p. [ Links ]
Anónimo. 1992. Ley de Pesca y su reglamento. Secretaría de Pesca. Talleres "Q" Editores. México, D.F. 68 p. [ Links ]
Anónimo. 1999. Climate diagnostics bulletin. No 99/9 Climate prediction center. NOAA. Camp Spring, Maryland. [ Links ]
Badan, A. 1997. La corriente Costera de Costa Rica en el Pacífico Mexicano, p. 99-113. In F. M. Lavín (ed.). Contribuciones a la Oceanografía Física en México. CONACYT, México, D. F. [ Links ]
Barcelata-Ortiz, M., J. de la O-Girón. & L. González-Márquez. 1990. Aspectos reproductivos de las especies: Pez Vela Istiophorus platypterus y Dorado Coryphaena hippurus capturados en las costas de Mazatlán, Sinaloa en 1988. Tesis Licenciatura. Universidad Autónoma de Sinaloa, México. [ Links ]
Baumgartner, R.T. & N. Christensen Jr. 1985. Coupling of the Gulf of California to large-scale interannual climatic variability. J. Mar. Res. 43: 825-848. [ Links ]
Beltrán, P. R. 1995. Análisis sobre las capturas de dorado en Mazatlán, Sinaloa. Informe Técnico, Inst. Nal. Pesca, CRIP-Mazatlán, Sinaloa, 14 p. [ Links ]
Fiedler, C.P. 1992. Seasonal climatologies and variability of Eastern Tropical Pacific surface waters. NOAA Tech. Rep., NMFS 108. 65 p. [ Links ]
Fiedler, C.P., F.P. Chavez, D. W. Behringer & S.B. Reilly. 1992. Physical and biological effects of Los Niños in the eastern tropical Pacific, 1986-1989. Deep-Sea Res. 39: 199-219. [ Links ]
García-Melgar, G. C. 1995. Ciclo de reproducción del dorado (Coryphaena hippurus) en el área de Los Cabos B.C.S. México. Tesis de licenciatura. Universidad Autónoma de Baja California Sur, 66 p. [ Links ]
Hubbs, C.L. 1948. Changes in the fish fauna of western North America correlated with changes in ocean temperature. J. Mar. Res. 7: 459-482. [ Links ]
Kojima, S. 1964. On the distribution of the dolphin, Coryphaena hippurus L., in the Pacific Ocean and the Indian Ocean. Bull. Japan. Soc. Sci. Fish. 30: 472-477. [ Links ]
Kojima, S. 1965. Studies on the fishing conditions of the dolphin Coryphaena hippurus L., in the western regions of the Sea of Japan-X. Results on tagging experiment. Bull. Japan. Soc. Sci. Fish. 31: 575-578. [ Links ]
Massuti, E., B. Morales-Nin & J. Moranta. 1999. Otholits microstructure and growth patterns of dolphin Corypahena hippurus in Western Mediterranean. Fish. Bull. 97: 891-899. [ Links ]
Norton, G. J . 1999. Apparent habitat extension of dolphinfish (Coryphaena hippurus) in response to climate transient in the California Current. p. 239-260. In E. Massutí & B. Morales-Nin (eds.). Biology and fisheries of dolphinfish and related species. Scientia Marina, Barcelona, España. [ Links ]
Norton, G. J. & S. J. Crooke. 1994. Occasional availability of dolphin, Coryphaena hippurus, to southern California commercial passenger fishing vessel anglers: Observations and hypotheses. Rep. CALCOFI 35: 230-239 p. [ Links ]
Oxenford, A., H. 1999. Biology of dolphinfish (Coryphaena hippurus) in the western central Atlantic: a review. P. 277-301. In E. Massutí & B. Morales-Nin (eds.). Biology and fisheries of dolphinfish and related species. Scientia Marina, Barcelona, España. [ Links ]
Oxenford, H. A. & W. Hunte. 1986. A preliminary investigation of stock structure of the dolphin Coryphaena hippurus in the Western Central Atlantic. Fish. Bull. 84: 451-460. [ Links ]
Patterson, K.R. & J. Martínez. 1991. Exploitation of the dolphin-fish Coryphaena hippurus. L. off Ecuador: Analysis by lenght-based virtual population analysis. Fishbyte 9: 21-23. [ Links ]
Ramp, R.S., L.J. McClean, A.C., Collins, J.A., Semtner & A.S.K Hays. 1997. Observations and modeling of the 1991-1992 El Niño signal off central California. J. Geophys. Res. 102: 5553-5582. [ Links ]
Santana-Hernández, H. 2001. Estructura de la comunidad de pelágicos mayores capturados con palangre en el Pacífico mexicano (1983-1996) y su relación la temperatura superficial del mar. Tesis doctoral. Universidad de Colima, México 122 p. [ Links ]
Walker, N. 1966. The origins and affinities of the Galapagos shorefishes, p. 72-74. In R. I. Bowman (ed.). The Galapagos Proceedings of the Symposia of the Galapagos Scientific Project. University of California. [ Links ]
Wyrtki, K. 1966. Oceanography of the eastern equatorial Pacific Ocean. Oceanograph. Mar. Biol. Ann. Rev. 4: 33-68. [ Links ]
Xie, L. & W.W. Hsieh. 1995. The global distribution of wind-induced upwelling. Fish. Ocean. 4: 52-67. [ Links ]

