











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.49 no.2 San José jun. 2001
Abstract
Three cruises, one in cach of the main stream patterns in the area, were made on the central Mexican Pacific continental slope (May-June 1995, November 1995, and March 1996). Seven trawl sampling sites were defined per cruise, and in each site four bathymetric levels were sampled (20, 40, 60, and 80 m). During the warmest seasons, the highest abundance of Porichthys margaritatus was at 60 m. When the temperature of the bottom water was lower, in March 1996, the highest abundance of P. margaritatus moved towards 40 m. The depths with greatest abundance fluctuated between 18 and 21°C, indicating a narrow temperature preference range. A Gaussian model was used to explain the relation between abundance and temperatura.
Key words
Porichlhys margaritatus, bathymetric distribution, temperature-abundance relation, Western tropical
Pacific, unimodal response model, Gaussian distribution.
One of the most important numerical methods for community analysis in the last years has been the family of multivaiiate techniques bases on the reciprocal averaging algorithm, as it is reflected in the scientific literature. These rnethods are supported by the argument that all species occur in a characteristic, limited range of habitats. Within their range, they tend to be most abundant around their particular environmental optimum (ter Braak and Prentice 1988). Although the Unimodal Response Model (URM) has been widely accepted to explain the way that species are distributed throughout environmental gradient, only a few studies has explicitly focussed in this assumption.
We analyze the abundance variation of Porichthys margaritatus (Richardson, 1844) in relation to temperature and depth like a direct gradient analysis (ter Braak and Looman 1995). In accordance with Aguilar-Palomino et al. (1996), the species P. margaritatus is considered an important component of the demersal softbottom fauna in the central Mexican Pacific and is frequently found in the shrimp bycatch fishery.
Materials and Methods
Three cruises were made aboard the RIV BIP V, on the continental slope of the central Mexican Pacific, one in each of the current patterns (Wyrtki 1965). The survey area is approximately 410 km2 and its main characteristics are described in Godínez-Domínguez and González-Sansón (1998). The first cruise (DEM I) was from May to June 1995 when the California Current and the North Equatorial Countercurrent converge in front of Jalisco and Colima; the second in November 1995 (DEM II) under the strong influence of the North Equatorial Countercurrent, a tropical water mass; and the third in March 1996 (DEM III) during the effect of the California Current, a cold water mass. Seven trawl sampling sites were established along the coast during each cruise. At each site, four stations were defined by depth (20, 40, 60, and 80 m). The samples were taken with two nets during nocturnal trawls lasting one-half hour at a speed of two knots. A detailed description of the fishing gear and sampling procedures can be found in González-Sansón et al. (1997). The temperature of the water near the bottom was recorded with a CTD profiler (SBE-19).
To analyze the effects of depth and season on organism abundance, a Friedman nonparametric analysis of variance was made (Zar 1996). A nonlinear Gaussian model (ter Braak and Looman 1995) was fitted to explain the relation of the abundance of organisms and temperature of the water mass. The formula of the Gaussian response model is:
Results
A total of 84 sampling tows were made along the continental shelf off Jalisco and Colima. We trawled for about 42 h and over an area of 100 ha. About 29 000 organisms of P. margaritatus were caught during the three cruises.
During the DEM I Cruise the highest average value of organism abundance (641 org/ha) was at 60 m depth (Fig. 1a), and the lowest abundance value at 20 m (5.4 org/ha, X2r=13.14, P<0.05). Water temperature decreased with depth. The maximum average value was recorded at 20 m (25.2 °C) and the niinimum of 18.5 °C was at 80 m. Tbe highest average abundance during cruise DEM II was at 60 m (Fig 1b), similar to the previous cruise (607 org/ha). The lowest mean value was at 20 m (6.3 org/ha). Values of abundance at depths differed (X2r= 1 5.35, P<0.05). The bottom water temperature was inversely related to depth, from 27.1 °C at 20 m to 16.7 °C at 80 m. The difference of the average water temperature between depths was maximum in this season and was 10.6 °C. The highest average abundance in the third cruise (DEM III) was at 40 m with 1 125 org/ha (Fig 1c). The lowest abundance value was 145 org/ha at 80 m and the abundance among depths were significantly different (X2r= 8, P<0.05). This indicates a movement of the P. margaritatus population towards shallower waters. The bottom water temperature decreased with depth. The fluctuation among the average values was 4.5 °C. The highest average temperature was 21 °C at 20 m and the lowest was 15.5 °C at 80 m.
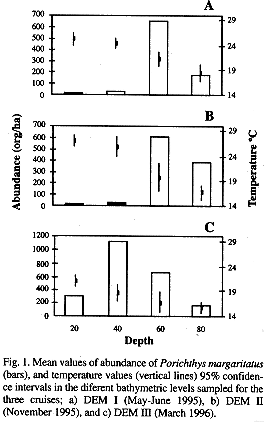
The highest abundance was in cruise DEM III when the water temperatura was lowest and the lowest abundance was found during DEM II with the warmest water (X2r= 8.5, P<0.05).
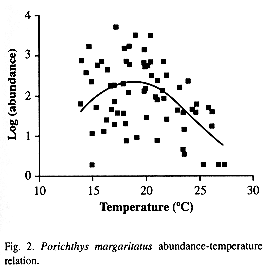
A direct relation between abundance and temperature means was observed only during the DEM III Cruise, because the relation observed during DEM I and II among both variables was nonlinear. Even though there was great heterogeneity of the abundance values (r=0.3), a significant (P<0.05) nonlinear relation was fitted to data (Fig. 2), where the range of temperature gradient considered was 14.4 °C (28.2 to 13.9 °C). The temperatura of maximum abundance (u) was 18.8 °C (C.I. 17.7 to 19.7 °C), the value of the tolerance (t) was 5.7 °C (C.I. 4 to 7.4 °C) and the maximum abundance (c) was 2.3 org/ha (C.I. 2.1 to 2.6 org/ha).
Discussion
Segregation patterns of bathymetrie distribution of soft-bottom demersal fauna have been reported in several studies (Bianchi 1991, 1992a, 1992b). A displacement of the greatest abundance of P. margaritatus towards shallower areas during the effect of the California Current in the area was observed. The depths with greatest abundance fluctuated between 21 °C in DEM I to 18.7 °C in DEM II. This indicates a narrow temperature preference range and could be the reason for displacement observed. We lack data of dissolved oxygen, however Bianchi (1991) suggests this factor could play an important role in the distribution of the demersal communities.
In agreement with some authors (Godínez-Domínguez and González-Sansón 1998; Filonov et al. 2000), coastal water showed tropical affinities during DEM I and II Cruises; the highest temperatura and a deeper thermocline (40-60 m). With the Califonia Current, the temperature was lower and the diermocline practically disappeared. Hence, the thertnocline could be related to P. margaritatus abundance, because the maximum abundance was located close to the thermocline.
A unimodal model to relate species to the environment has been widely used (ter Braak 1987), and this Gaussian relation becomes visible when a sufficient range of the environment variable is considered. We considered almost a 15 °C range (10-90 m) during a hydroclimatic cycle. Several authors suggest that other factors as competition (Gause 1934) and a complex set of variables included in a concept named "habitat suitability" (MacCall 1989) could influence species response to an environmental gradient. The abundance distribution of the benthic invertebrates where P. margaritatus are feeding varies similarly to P. margatitatus abundance in the same period (Godínez-Domínguez and González-Sansón 1998).
Aknowledgements
This work was financed by the University of Guadalajara, SEMARNAP permit No. 020298213-03. Our thanks to the efficient crew of the R/V Barco de Investigación Pesquera BIP V and to our colleagues of the "Demersal Project" and "Turtles Project" for their help. Thanks to Ellis Glazier for editing the English-language text.
Resumen
Se analizaron tres cruceros de pesca demersal, uno por cada pauta principal de circulación marina en la zona (mayo-junio de 1995, noviembre de 1995, y marzo de 1996). Se determinaron siete sitios de muestreo en cada crucero y en cada sitio se muestrearon cuatro estratos batimétricos (20,40, 60 y 80 m). Durante la época cálida, las mayores abundancias de Porychthys nwrgaritatus se encontraron en 60 m. Cuando la temperatura de fondo fue mínima durante marzo de 1996, las mayores abundancias se desplazaron a 40 m de profundidad. La temperatura de máximas abundancias fluctuaron entre 18 y 21 °C, lo cual indica un estrecho rango de preferencia térmica. Se propone un modelo Gaussiano para explicar la relación entre la abundancia y la temperatura.
References
Aguilar-Palomino, B., J. Mariscal-Romero, G. González. Sansón & L.E. Rodríguez-Ibarra. 1996. Ictiofauna demersal de fondos blandos de la plataforma continental de Jalisco y Colima, México.Primavera de 1995. Cien. Mar. 22: 469-481. [ Links ]
Bianchi, G. 1991. Demersal assemblages of the continental shelf and slope edge between the Gulf of Tehuantepec (Mexico) and the Gulf of Papagayo (Costa Rica). Mar. Ecol. Progr. Ser. 73: 121-140. [ Links ]
Bianchi, G. 1992a. Demersal assemblages of the continental shelf an upper slope of Angola. Mar. Ecol. Progr. Ser. 81: 101-120. [ Links ]
Bianchi, G. 1992b. Study of the demersal assemblages of continental shelf and upper slope off Congoand Gabon, based on trawl surveys of the RV "Dr Fridtjof Nansen". Mar Ecol. Progr. Ser. 85: 9-23. [ Links ]
Filonov, A., I.E. Tereshchenko, C.O. Monzón, M.E. González-Ruelas & E. Godínez-Domínguez. 2000. Resultados del estudio de la variabilidad estacional de los campos de temperatura y salinidad cerca de la zona costera de los estados de Jalisco y Colima, México. Cien. Mar. 26: 303-321. [ Links ]
Gause, 1934. The struggle for existence. Williams and Wilkin, Baltimore. [ Links ]
Godínez-Domínguez, E. & G. González-Sansón. 1998. Variación de los patrones de distribuciónbatimétrica de la fauna macrobentónica en la plataforma continental de Jalisco y Colima, México. Cien. Mar. 24: 337-351. [ Links ]
González-Sansón, G., B. Aguilar-Palomino, J. Arciniega-Flores, Garcia de Quevedo-Machaín, R., E. Godínez-Domínguez, V. Landa-Jaime, J. Mariscal-Romero, J.E. Michel-Morfín & M. Saucedo-Lozano. Variación espacial de la abundancia de la fauna de fondos blandos en la plataforma continental de Jalisco y Colima, México (primavera, 1995). Cien. Mar. 23: 93-110. [ Links ]
MacCall, A.D. 1989. Dynamic geography of marine fish populations. Washington Sea Grant Program. University of Washington, Seattie. 153 p. [ Links ]
ter Braak, C.J.F. 1987. General introduction. In C.J.F. ter Braak (ed.). Unimodal Models to Relate Species to Environment. Agricultural Mathematics Group,The Netherlands. 152 p. [ Links ]
ter Braak, C.J.F. & C.W.N. Looman, 1995. Regression. In R.H.G. Jongman, C.J.F. ter Braak & O.F.R. van Tongeren (eds.). Data in Community and Landscape Ecology. Cambridge University, London. 299 p. [ Links ]
ter Braak, C.J.F & I.C. Prentice. 1988. A theory of gradient analysis. Adv. Ecol. Res. 18: 271-317. [ Links ]
Wyrtki, K. 1965. Surface currents of the eastern tropical Pacific Ocean. Inter. Amer. Trop. Tuna Comm., Bull. IX : 271-294. [ Links ]
Zar, JH. 1996. Biostatistical Analysis. Prentice Hall, New Jersey. 662 p. [ Links ]
1Centro de Ecología Costera, Universidad de Guadalajara. México.
2 Current address; Departamento de Biologia Animal, Vegetal y Ecología. Universidad de A Coruña.
Campus da Zapateira s/n, A Coruña, España. 15071; egodinez@mail2.udc.es
3 Centro de Investigaciones Marinas, Universidad de la Habana. La Habana, Cuba.

