











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.49 no.2 San José Jun. 2001
Received 28-IX-2000. Corrected 4-X-2000. Accepted 9-10-2000
Abstract
Bahía Concepción (Gulf of California) was studied to describe some extremely low dissolved-oxygen values at the bottom of the bay. Surveys included measurements of vertical distribution of temperature, dissolved oxygen, nitrate, nitrite, phosphates and hydrogen sulfide. Thermal stratification from early spring to autumn and a well-mixed water column during the winter were present. Dissolved-oxygen concentration was homogeneous in the water column from winter to spring (5-7 mg L-1). From summer to autumn, bottom dissolved oxygen (> 20 m) decreased to reach conditions of hypoxia (1-2 mg L-1) or anoxia; nitrate, nitrite, and phosphates increased (up to 13, 1.7 and 2.2 *M) followed by a last short period of a deep hydrogen sulfide layer (up to 3.1 mg L-1). A back calculation to estimate the input of organic matter in the deep layer showed that 18 g C m-2y-1 needed to be trapped in the bottom to induce such an oxygen depletion. This period is thought to be unfavorable for bottom infauna in this bay.
Key words
Anoxic basin, organic matter inputs, hydrography, nutrients, Bahía Concepción, Gulf of California.
Seasonal anoxic or hypoxic conditions have often been observed in temperate coastal environments (May 1973, Santos and Simon 1980, Jørgensen 1990, Babenerd 1991, Prena 1994) and to a lesser extent in tropical marine environments (Vargas 1995). The frequency, severity, and length of hypoxic conditions are mainly a consequence of a hydrographic structure causing water stagnation (Jørgensen 1990), or by the inflow of deep oxygen-depleted waters from adjacent regions (Prena 1994). The decrease in dissolved oxygen in bottom waters together with a stable halocline or thermocline induces high periods of oxygen stress that may have severe ecological consequences on bottom fauna (Theede et al. 1969) and demersal fish (May 1973, Officer et al. 1984).
Along the Mexican Pacific coast, several events of fish mortality have been observed but never studied. In Bahía Concepción (Gulf of California), aperiodic defaunation has occurred, mainly in soft-bottom mollusks. Some unpublished reports briefly explained such mass mortalities by abnormal increases in water temperature. Preliminary studies have identified also an abnormal decrease of dissolved oxygen (Reyes Salinas 1994). To describe this process of anoxia, we have monitored water quality variables and the physical structure of the water column. On the basis of the data obtained, we analyzed for either natural or anthropogenic influences, as causes of the environmental disturbances in this bay.
Bahía Concepción: Bahía Concepción is one of the deepest and largest (282 km2) coastal bays on the western side of the Gulf of California, in an arid climate region (Fig. 1). A large shallow southern plain and a northern 10-m deep sill near the bay entrance border the bays central basin (30- to 35-m depth).
This bay has higher salinity (35.1 to 36.0, Gilmartin and Revelante 1978) than in the adjacent Gulf (< 35.2) because of high evaporation, particularly during summer. The terrestrial area is desertic, with an annual rainfall of about 113 mm. According to Gilmartin and Revelante (1978), Bahía Concepción has a total volume of 4553x106 m3 and an intertidal volume of 221x106 m3, giving a relatively low flushing index of 0.05.
The only urban development, the village of Mulegé, is 3 km north of the entrance of the bay (Fig. 1). It has about 3,000 inhabitants. The exploitation of shellfish, as Argopecten ventricosus, (Sowerby II), was the main activity in the bay (Villalejo-Fuente and Ochoa-Báez 1993), however, the annual yield was highly variable from year to year (1734 ± 1582 t, Sepesca, 1993). Extensive and local events of shellfish defaunation occurred mainly during the summer (Massó and Vélez pers. comm.), probably contributing to the inconsistency of the yield.
Materials and Methods
Hydrographic surveys were made in 1993 in March, June, September, October, and December, and in 1994, in March, April, May, June, and July.
In the 1993 surveys, four stations were sampled along the axis of the central basin of the bay (Fig. 1). Water samples were taken with Van Dorn bottles at 0, 1, 3, 5, 7, 10, 15, and 20 m depths, except in October and December, when additional samples at 25 to 30 m depth were taken. During the 1994 surveys, three or four stations were sampled on a transverse axis of the bay (Fig. 1) and water samples were taken at 0, 5, 10, 15, 20, 25, and 30 m.
Temperature was measured with a YSI or HACH probe. Dissolved-oxygen (DO) concentration was estimated by the Winkler titration technique, orthophosphate was estimated according to Murphy and Riley (1962), and nitrate and nitrite according to Parsons et al. (1984). Nutrient samples were filtered on Whatman GF/C filters. Analyses of hydrogen sulfide were made on samples from 0, 20, and 30-m depths using the iodometric procedure described in Clesceri et al. (1989).
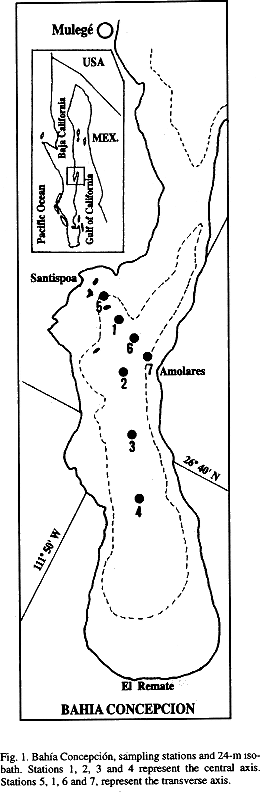
Wind data (provided by the Comisión Nacional del Agua) were obtained from a meteorological station at Mulegé, 3 km north of the entrance of Bahía Concepción (Fig. 1). Monthly wind averages were computed from the 1993 measurements.
Results
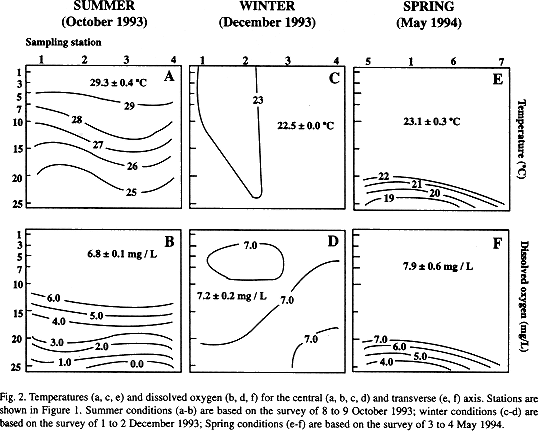
Our data were consistent with the seasonal water structure reported earlier (Lechuga-Devéze and Morquecho-Escamilla 1998). Summer is characterized by a stratified water column, identified by a well-developed thermocline and oxycline (Figs. 2a, 2b). This structure is broken in winter to a totally homogeneous water column. The temperature and oxygen shows a little variation along and across the central basin and depth (Figs. 2c, 2d). The restart to a stratified condition is in spring (Figs. 2e, 2f) when the surface layer is again warmed by the seasonal warming.
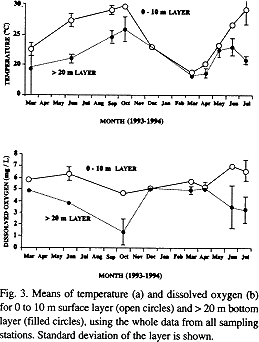
According the expected vertical structure of temperature and dissolved oxygen, the whole data (sampling stations 1 to 7) were monthly pooled in 0 to 10 m layer (surface layer; n = 7 to 28), and >20 layer (bottom layer; n = 2 to 8). Lowest n numbers (7 and 2) were for March 1993 (data from sampling station 1). ANOVA and TUKEY test compared temperature and dissolved oxygen data of the two layers; depth and time were the independent variables. At 95% of confidence level, both layers were proved to be significantly different. The weight of this significance was the summer months. Temperature is consistently higher, up to 8 °C, at the surface than in the bottom layer during March to October (Fig. 3a). In November-December, both surface and bottom layers are mixed by a decrease in temperature (around 22 °C), and in March, the entire water column reaches its lowest temperature (around 18 °C). From April to July, once again, the surface and bottom layers are divided by the thermocline.
During this period of thermal stratification, the bottom layer showed a decrease in dissolved oxygen concentration (Fig. 3b). The lowest values were in October (1.3 ± 1.1 mg O2/L), and this extended over the entire basin (Fig. 2b).
By the period of thermal homogenization (winter), values of dissolved oxygen did not show differences between surface and bottom layers (Figs. 2d, 3b). Once again, the cycle towards the stratification is restarted when the thermocline is developed, and the bottom oxygen is depleted.
Dissolved oxygen in the surface layer did not show a seasonal trend, remaining between 5 to 7 mg O2/L.
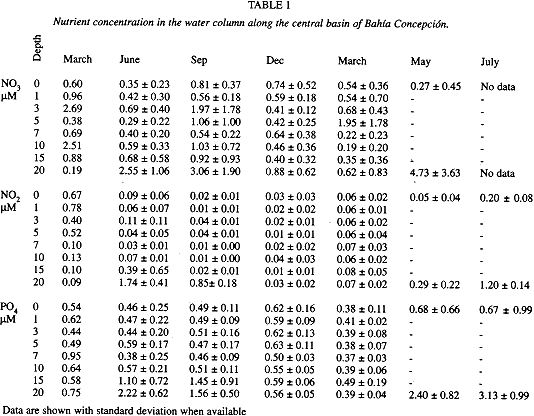
The isolation of the bottom layer by the thermocline, depleted the dissolved oxygen by organic matter oxidation. This process is thought to be responsible for the increase of nitrates, nitrites, and orthophosphates during the summer (table 1). In spite of the high variability of nutrient concentration at surface and bottom layers, the bottom values clearly are well differentiated from those of the surface.
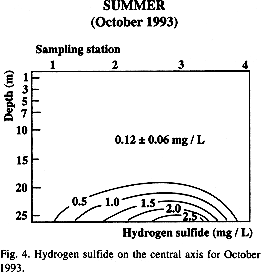
Low oxygen levels or bottom anoxia observed in October 1993 along the central axis of the bay, was also conducive to the production of hydrogen sulfide (Fig. 4, Table 2). The highest concentration (up to 3.1 mg L-1) was found at sampling station 3 when bottom anoxia was recorded (Fig. 2b and 4). The slight increase in dissolved oxygen at 20 m was followed by a decrease in hydrogen sulfide (0.5 mg L-1) because dissolved sulfide oxidizes rapidly in the presence of oxygen. This hydrogen sulfide layer disappeared in the winter when the water column was mixed (Table 2).

Discussion
Origin of the anoxic layer: The temperature structure in Bahía Concepción is reasonably well defined throughout the annual cycle. In winter (December to March), the whole bay is well mixed probably by a) the convective processes forced by a NW-NE wind stress (Dressler 1981) that might drive waters from the Gulf of California into the bay, and/or b) the fast cooling of surface waters.
The beginning of thermal stratification could be a response to a change of hydrographic forces driven by the change of wind direction (Fig. 5). In early spring (April), this lowering of energy in the water column, and warming of air temperature, act together to create and maintain a strong pycnocline isolating the surface (0 to 10-m) from bottom waters (> 15-m).
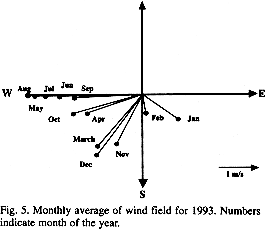
Changes in the oxygen concentration at the bottom layer could be correlated with a) the oxygen consumption from biological oxidation of organic matter (May 1973, Babenerd 1991, Llansó 1992), or b) intrusions of adjacent oxygen-depleted bottom waters advected by wind influence (Officer et al. 1984, Breitburg 1990, Prena 1994). The advection of low oxygen waters coming from the ocean into Bahía Concepción does not seem to occur. The low DO waters of the Gulf of California (1.0 mg O2 L-1) are present at 300 m, and the thermocline is well-developed in summer acting as a barrier to impede the upwelling of this water (Emilsson 1993). Therefore we can explore the in situ inputs of oxidizable material to the bay.
Bahía Concepción is in an arid area, where the annual precipitation averages 113 mm. There is no agricultural or industrial land use. The only inputs of organic matter from human activity originate at Mulegé village (3000 inhabitants). The urban wastewater is treated before being discharged into a small river flowing 3 km north of the mouth of the bay. Some tourist facilities are along the coast of the bay. The nutrient or carbon inputs from this very low population density, even though not measured, seem easily diluted upon reaching the bay. Natural inputs of organic matter seem be supplied by red tides of Noctiluca scintillans occurring every winter (Gárate-Lizárraga 1991, 1995). Lechuga-Devéze and Morquecho-Escamilla (1999) showed, also in winter, a diatom-dominated population.
Martínez-López and Gárate-Lizárraga (1994) reported a widespread spawning of shellfish, again in the winter. It seems the physical and chemical conditions of winter render the whole bay capable of sustaining an environment rich in autotrophs and heterotrophs. This produces a high grazing pressure and therefore an increase in the settling of organic particles. Further, plankton biomass and organic debris could be trapped in the bottom layer by the density gradient formed during spring to summer.
The gradual decrease of dissolved oxygen at the bottom during spring and summer are directly correlated to an increase in nutrients. This process ends in late summer to mid-autumn when hypoxic conditions are reached. When the DO is depleted, and organic nitrogen has been oxidized into NO3 and NO2, the anaerobic metabolism seems to be stimulated, promoting the production of hydrogen sulfide (Theede et al. 1969).
Rates of carbon degradation: We can obtain a rough estimate of the rate of carbon degradation through the rates of oxygen consumption of the deep stagnant water (> 20 m). This assumes no significant oxygen exchange between surface and bottom layers (e.g. diffusion from the photic layer, tidal mixing, wind mixing), and the main input of organic matter is from the winter plankton biomass. The mean rate of oxygen consumption during March to October 1993 and March to July 1994, was 0.028 ± 0.006 mg O2L-1 d-1 (equals to 0.028 ± 0.006 g O2 m-3 d-1). Using a 10-m depth of stagnant bottom water (20 to 30 m depth), we obtained a consumption of 0.28 g O2 m-2 d-1. Because 3.47 g of oxygen is used to oxidize 1 g of organic carbon, we obtained a rate of 0.081 g C m-2 d-1. To extrapolate for the total organic matter load, we again assumed a linear oxygen depletion during 225 days, obtaining 18 g C m-2 y-1 (1.6 mol C m-2y-1) accumulated during the winter in the bottom of Bahía Concepción. Using the standard deviation of data (0.006) the range could extend from 14 to 22 g C m-2 y-1.
Other environments affected by discharges of anthropogenic organic matter, as the Baltic Sea (the Öresund and Western Baltic), had rates and organic loads near the range obtained in this study. For the Öresund, Mattsson and Stigebrandt (1993) reported an oxygen consumption rate of 0.038 mg O2 L-1d-1 and an annual flux into the basin of 20.7 g C m-2 y-1. For the Western Baltic, Balzer(1984) reported 2.5 mol C m-2 y-1 (30 g C m-2 y-1) of organic matter decomposed at the sediment water interface.
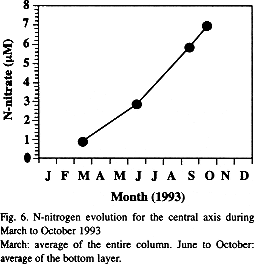
The similarities with our rates may be fortuitous and very speculative. However to validate this first approach, we can also estimate the organic load through the N-nitrate release (Fig. 6) using:
N-nitrate rate (mmol m-2 d-1) = [(Nf – Ni)/ dt] Sw
where Ni is the initial mean of N-nitrate concentration for the entire water column (March: 0.977 mmol m-3), Nf is the final mean of N-nitrate concentration for the deep layer (October: 6.89 mmol m-3), dt is the number of days of this period (196 days), and Sw is the depth of stagnant layer (10 m). Using this formula, we obtain an estimate of 0.3015 mmol N m-2 d-1.
Calculating 225 days (21 March to 31 October) as the total period of water stagnation, we obtain a rate of 68 mmol N m-2 y-1, or 5.4 g C m-2y-1 [calculated by: mmol N x (N: P) x 12,where N: P = 106: 16, and 12 is the molecular weight of carbon], about one-third of the carbon computed from oxygen data in this work. This rate is near to that reported for the Western Baltic Sea (Balzer 1984). There, 160 mmol N m-2 y-1 or 12.7 g C m-2 y-1 represents a little less than one third of the oxidation of 30 g C m-2 y-1 (2.5 mol C m-2 y-1).
We have assumed for this area there may have been no oxygen influences from the surface layers and the organic load comes from winter inputs. However, these assumptions could be inaccurate because of direct vertical episodic mixing or diffusion through the thermocline. Nevertheless, the calculated total organic matter load, certainly underestimated, represents an adequate approach that encourages further examination.
Causal effects of oxygen depletion: Anthropic activities should promote coastal eutrophication resulting in an increased organic load and, consequently, increased oxygen demand leading to episodic or aperiodic anoxic conditions (May 1973, Llansó 1992, Prena 1994). However, the development of anoxia in Bahía Concepción cannot be associated with human impact because the area has a low population density. Furthermore, according to Gilmartin and Revelante (1978) data on anoxic bottom, high nitrate and phosphate values were first observed in 1976. This could indicate that anoxic conditions are more related to natural influences.
Tilting fluctuation of isotherms originating from the tidal and wind motion of surface waters should induce a gentle upwelling into the shallows of the southern plain (El Remate) advecting the anoxic and hydrogen sulfide-rich bottom layer, where shellfish banks are located (Félix-Pico 1987). If the tolerance to oxygen deficiency and hydrogen sulfide by marine bivalves is affected by high temperatures (Theede et al. 1969), and the mean lethal time in these conditions is around 4 days (Barthel 1984), then anoxia and hydrogen sulfide should be responsible for some sudden summer to autumn mollusk mortality. In the light of these data, Bahía Concepción has recently been considered as an endangered area (Arriaga et al. 1998)
Conclusions
The decreased oxygen content in the deep layer of Bahía Concepción is most likely a seasonal increase to oxidize natural inputs of organic matter. Extensive non-periodic shellfish mortality could be linked to wind- or tide-induced advection of the anoxic layer into the shellfish banks. It is evident that the coupling of natural processes of organic matter production, water mass structure, and biogeochemical transformations in the sediments dominates to a great extent the biological environment of Bahía Concepción.
Acknowledgments
The authors are grateful to D. López-Cortés and J. Hernández-Alfonso, for the coordination of several surveys; to F. Hernández and I. Murillo, for skillful chemical analyses. Part of this work was presented by one of the authors (R.S.A.) to fulfill the requirements for a Bachelor Degree in Biology (UNAM). This work was financially supported by the Centro de Investigaciones Biológicas del Noroeste, S.C.: institutional projects E3 (1993), and E2 and E3 (1994). Valuable comments by A. Genin, J. Bustillos and anonymous reviewers are fully appreciated. R. Bowers and E. Glazier edited the English-language manuscript.
Resumen
Se midió seis variables físico-químicas en Bahía Concepción (Golfo de California, 1993 y 1994) para explicar el origen de valores muy bajos de oxígeno disuelto en el fondo. La estructura hidrográfica mostró una fuerte estratificación térmica vertical del inicio de la primavera hasta el otoño, mientras que durante el invierno se observa una columna de agua homogénea. Las concentraciones de oxígeno disuelto siguen el mismo patrón: homogéneas de invierno a primavera (5-7 mg L –1), mientras que del verano al otoño disminuyen en la capa de fondo (> 20-m) hasta alcanzar condiciones de hipoxia (1-2 mg L-1) o anoxia. Las concentraciones de nitrato, nitrito y ortofosfatos también aumentan en la capa de fondo (13, 1.7 y 2.2 *M respectivamente), seguido de un corto período de formación de sulfuro de hidrógeno (hasta 3.1 mg L-1). Por cálculos indirectos se establece que 18 g C m-2 a-1 de materia orgánica son atrapados en el fondo provocando la severa disminución del oxígeno disuelto. Este período se considera crítico para la fauna bentónica de esta bahía.
References
Arriaga Cabrera, L., E. Vázquez-Domínguez, J. González-Cano, R. Jiménez Rosenberg, E. Muñoz López, V. Aguilar Sierra (coords). 1998. Regiones prioritarias marinas de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, 198 p. [ Links ]
Baberned,B. 1991. Increasing oxygen deficiency in Kiel Bay (Western Baltic). A paradigm of progressing coastal eutrophication. Meeresforsch. 33: 121-140. [ Links ]
Balzer,W. 1984. Organic matter degradation and biogenic element cycling in a nearshore sediment (Kiel Bight). Limnol. and Oceanogr. 29: 1231-1246. [ Links ]
Barthel,D. 1984. Adenylate energy charge in three marine bivalve species during anoxia. Ophelia 23: 155-164. [ Links ]
Breitburg,D.L. 1990. Near-shore hypoxia in the Chesapeake Bay: patterns and relationships among physical factors. Est. Coast. and Shelf. Sci. 30: 593-609. [ Links ]
Clesceri. L.S, A.E. Greenberg & R.R. Trussell. 1989. Standard Methods for the examination of water and wastewater 17h edition, American Public Health Association, American Water Works Association, Water Pollution Control Federation, Washington D.C., 4-197 p. [ Links ]
Dressler,R. 1981. Preliminary knowledge of hydro-static summer conditions within Bahía Concepción, Baja California. Technical Report, Centro de Investigaciones Científicas y Estudios Superiores de Ensenada, México,13 p. [ Links ]
Emilsson,I. 1993. Oceanografía físico-química del Golfo de California. Technical Report,Centro Interdisciplinario de Ciencias Marinas, La Paz, B.C.S. 64 pp. [ Links ]
Félix-Pico,E.F. 1987. Scallop fishing in Conception Bay, Baja California Sur, México. In: Book of abstracts of the 6th International Pectinid Workshop, Menai Bridge, 12 pp. [ Links ]
Gárate-Lizárraga,I. 1991. Análisis de una marea roja causada por Noctiluca scintillans (McCartney) Ehr. en Bahía Concepción, Baja California Sur, en febrero de 1989. Rev. Inv. Cient. Universidad Autónoma de Baja California Sur 2: 35-43. [ Links ]
Gárate-LizárragaI. 1995. Mareas rojas en Bahía Concepción, B.C.S., México. Boletín No. 40, Centro Interdisciplinario de Ciencias Marinas, 2 p. [ Links ]
Gilmartin, M. & N. Revelante. 1978. The phytoplankton characteristics of the barrier island lagoons of the Gulf of California. Est. Coast. Mar. Sci. 7: 29-47. [ Links ]
Jørgensen,B.B. 1990. Seasonal oxygen depletion in the bottom waters of a Danish fjord and its effects on the benthic community. Oikos 34: 68-76. [ Links ]
Lechuga-Devéze, C.H. & M.L. Morquecho-Escamilla. 1998.Early spring potentially harmful phytoplankton in Bahía Concepción, Gulf of California. Bull. Mar. Sci.63: 503-512. [ Links ]
Llansó,R. 1992. Effects of hypoxia on estuarine benthos: the lower Rappahannock river (Chesapeake Bay), a case study. Est. Coast. and Shelf. Sci. 35: 491-515. [ Links ]
Martínez-López, A. & I. Gárate-Lizárraga. 1994. Cantidad y calidad de la materia orgánica particulada en Bahía Concepción en la temporada de producción de almeja catarina Argopecten circularis (Sowerby, 1835). Ciencias Marinas 20: 301-320. [ Links ]
Mattsson, J. & A. Stigebrandt. 1993. The vertical flux of organic matter in the Öresund estimated by two different methods using oxygen measurements. Est. Coast. and Shelf. Sci. 37: 329-342. [ Links ]
May,E.B. 1973. Extensive oxygen depletion in Mobile Bay, Alabama. Limnol. and Oceanogr. 18: 353-366. [ Links ]
Murphy, J. & J.P. Riley. 1962. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 27: 31-36. [ Links ]
Officer, C.B., R.B. Biggs, J.L. Taft, L.E. Cronin, M.A. Tyler & W.R.Boynton. 1984. Chesapeake Bay anoxia: origin, development and significance. Science 223: 22-27. [ Links ]
Parsons, T.R., Y. Maita & C.M. Lalli. 1984. A manual of chemical and biological methods for seawater analyses. Pergamon Press, New York, 173 pp. [ Links ]
Prena,J. 1994. Oxygen depletion in Wismar Bay (Western Baltic Sea) 1988. Arch. Fish. Mar. Res. 42: 77-87. [ Links ]
Reyes Salinas, A. 1994. Relación entre estructura hidrográfica y la abundancia, distribución y origen de diferentes expresiones de biomasa del seston orgánico en Bahía Concepción, Golfo de California. Thesis. Escuela Nacional de Estudios Profesionales, Iztacala, UNAM. 53 p.
Santos, S.L. & J.L. Simon. 1980. Response of soft-bottom benthos to annual catastrophic disturbance in a south Florida estuary. Mar. Ecol. Progr. Ser. 3: 347-355. [ Links ]
Sepesca (Secretaría de Pesca). 1993. Sistema Descentralizado de Información Pesquera. Secretaría de Pesca, Delegación Federal de Pesca en La Paz, B.C.S., 117 p. [ Links ]
Theede, H., A. Ponat, K. Hiroki & C. Schlieper. 1969. Studies on the resistance of marine bottom invertebrates to oxygen-deficiency and hydrogen sulfide. Mar. Biol. 2: 325-337. [ Links ]
Vargas, J.A. 1995. The Gulf of Nicoya estuary, Costa Rica: Past, present, and future cooperative research. Helgolander Meeres. 49: 821-828. [ Links ]
Villalejo-Fuente, M. & R.I. Ochoa-Báez. 1993. El ciclo reproductor de almeja catarina Argopecten circularis (Sowerby, 1835), en relación con la temporada y fotoperíodo en Bahía Concepción, B.C.S., México. Ciencias. Marinas 19: 181-202. [ Links ]
1 Centro de Investigaciones Biológicas del Noroeste, S.C., P.O. Box 128, 23000 La Paz, Baja California Sur, México, fax: +52 (112) 547 10; clechuga@cibnor.mx

