










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.1 San José Mar. 2001
Alberto de Jesús Navarrete 1
Recibido 15-III-2000. Corregido 13-VII-2000. Aceptado 31-VII-2000
Abstract
The growth rate of queen conch cultured in pens was studied from October 1993 to March 1994. Sixteen pens (50 m2 each, four pens per environment), were set in four environments: Thalassia, Thalassia-sand, Sand and Coral within a reef lagoon on Punta Gavilan and Banco Chinchorro. Twenty conchs were introduced in each pen (sizes: 100-120, 120-140, 140-160 and 160-180 mm shell length) and measured monthly to the nearest mm. Growth rate was assessed by two methods: a) shell marginal mean increase and b) the Gulland-Holt method considering all conch within pens. In the first method, the environment Sand had the highest growth (3.21 ± 0.26 mm/month) at Punta Gavilan, whereas at Banco Chinchorro, highest growth was recorded in Coral (2.31 ± 0.44 mm/month). Considering the second method, highest asymptotic length conch in Punta Gavilan occurred in Thalassia-sand (287.5 mm), whereas in Banco Chinchorro the highest asymptotic length was measured in Sand (318.1 mm). There were significant differences in growth between sites; juvenile growth is related with habitat quality mainly food availability.
Key words: Caribbean, semi-culture, Gastropoda, Strombus gigas, Quintana Roo.
En el sur de Quintana Roo, el caracol rosado (Strombus gigas Linnaeus 1758) representa la segunda pesquería económicamente importante, después de la langosta espinosa (Panulirus argus Latreille 1804) (Sosa-Cordero et al. 1993). Debido a la presión de pesca en los últimos años, las poblaciones de caracol han disminuido de tal forma que el recurso se considera sobrexplotado (Chávez y Arreguín 1994, de Jesús-Navarrete y Oliva-Rivera 1997).
Este panorama es compartido por otras naciones del Caribe y ha dado lugar a diversas prácticas de manejo, como vedas, talla mínima, cuotas de captura y en muchos casos el cierre total de la pesquería. Sin embargo, las poblaciones no muestran una recuperación sustancial (Stoner y Ray 1996).
El cultivo se ha planteado como una posibilidad para repoblar áreas y como una medida complementaria en la explotación del caracol. Actualmente puede decirse que las técnicas de cultivo, desde larvas a juveniles de 5 cm de longitud, están dominadas (Brownell 1977, Creswell 1984, Cruz 1984, Stoner y Sandt 1988, Rodríguez y Ogawa 1990, Davis, 1994, de Jesús-Navarrete 1997). El cultivo de ciclo incompleto o semicultivo es una alternativa de manejo del recurso con dos opciones: la generación de semillas para repoblación de bancos naturales, y la engorda de organismos en encierros bajo condiciones similares a las de su hábitat hasta que alcancen la talla comercial. En Quintana Roo, los pescadores han planteado el traslado de caracoles de zonas con densidades altas a áreas despobladas, como una medida de repoblación, pero en realidad los caracoles son colocados en encierros hasta que se autoriza la captura, sin que exista una evaluación del efecto de este traslado sobre los organismos. El objetivo de este trabajo fue estimar el crecimiento en ambientes y sitios diferentes, para tener elementos que permitan decidir si establecer encierros es adecuado y se pretende además contribuir a un mejor entendimiento en el manejo del recurso pesquero.
Resultados
El incremento marginal mensual de la concha de los caracoles se muestra en el Cuadro 1. En PG, el incremento promedio más alto ocurrió en el ambiente Arena, con 3.21 ± 0.26 mm/mes, seguido por Thalassia-arena con 2.15 ± 0.93 mm/mes y finalmente Coral y Thalassia con 2.04 ± 0.73 mm/mes y 1.49 ± 0.58 mm/mes respectivamente. Para BCH, los incrementos en la concha de los caracoles fueron más altos en el ambiente Coral, con 2.31 ± 0.44 mm. Otro ambiente en donde se observó un buen incremento fue Arena con 2.01 ± 0.69 mm/mes. El menor crecimiento se presentó en Thalassia con 1.51 ± 0.21 mm/mes.
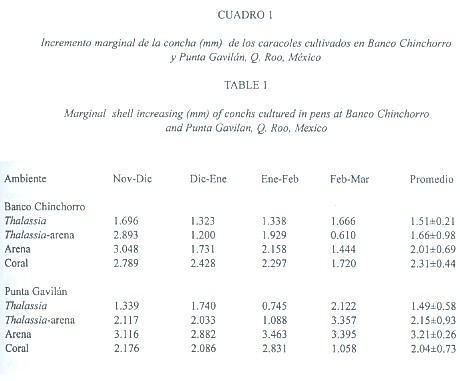
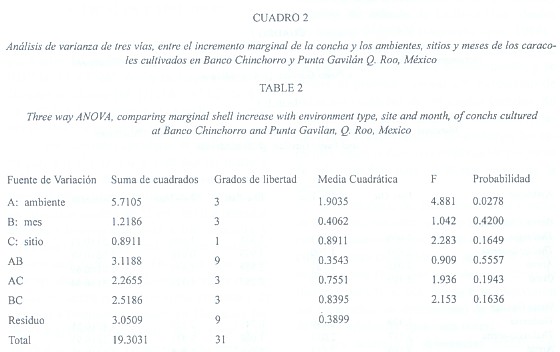
El análisis de varianza (ANOVA, tres vías, p< 0.05)) mostró diferencias significativas en el crecimiento entre ambientes, lo que explica las diferencias en los crecimientos en Thalassia-arena y Coral. No se encontraron diferencias significativas en el crecimiento entre los sitios, ni entre los meses, ni entre las interacciones entre las fuentes de variación (Cuadro 2).
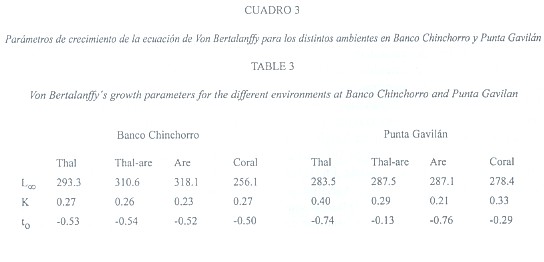
Los parámetros de crecimiento de la ecuación de Von Bertalanffy obtenidos con el método de Gulland-Holt, se muestran en el Cuadro 3. Los valores de la L![]() fueron muy similares entre los ambientes de Punta Gavilán y variaron entre 278.4 a 287.5 mm de longitud de concha, mientras que K, varió de 0.21 en el ambiente Arena a 0.40 en Thalassia. En Banco Chinchorro, los valores de L
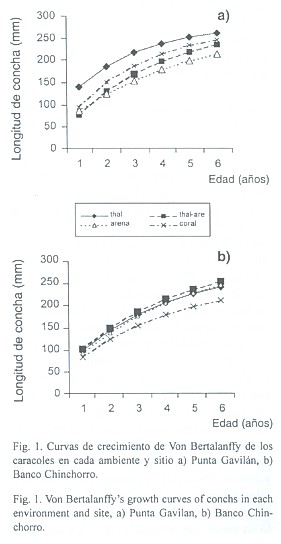
fueron muy similares entre los ambientes de Punta Gavilán y variaron entre 278.4 a 287.5 mm de longitud de concha, mientras que K, varió de 0.21 en el ambiente Arena a 0.40 en Thalassia. En Banco Chinchorro, los valores de L![]() variaron de 256.1 en Coral a 318.1 en Arena, la tasa de crecimiento K, fue ligeramente más baja que en Punta Gavilán y varió de 0.23 a 0.27, respectivamente. Al integrar esos valores a la ecuación de Von Bertalanffy, se obtienen las curvas de crecimiento para cada ambiente y se puede observar que en PG, la curva correspondiente al ambiente Thalassia, se encuentra por encima de las demás e indica que los caracoles crecieron mejor que en el resto de los otros ambientes. El análisis de covarianza indicó que no hubo diferencias significativas en la pendiente de las curvas (F 0.05, 2, 19), pero si en la posición de las mismas (F 0.05, 2, 22), lo que refuerza la afirmación que en su conjunto ocurre un mejor crecimiento en Thalassia. El siguiente sitio fue Coral y los ambientes con menor crecimiento fueron Thalassia-arena y Arena.(Fig.1a).
variaron de 256.1 en Coral a 318.1 en Arena, la tasa de crecimiento K, fue ligeramente más baja que en Punta Gavilán y varió de 0.23 a 0.27, respectivamente. Al integrar esos valores a la ecuación de Von Bertalanffy, se obtienen las curvas de crecimiento para cada ambiente y se puede observar que en PG, la curva correspondiente al ambiente Thalassia, se encuentra por encima de las demás e indica que los caracoles crecieron mejor que en el resto de los otros ambientes. El análisis de covarianza indicó que no hubo diferencias significativas en la pendiente de las curvas (F 0.05, 2, 19), pero si en la posición de las mismas (F 0.05, 2, 22), lo que refuerza la afirmación que en su conjunto ocurre un mejor crecimiento en Thalassia. El siguiente sitio fue Coral y los ambientes con menor crecimiento fueron Thalassia-arena y Arena.(Fig.1a).
En BCH, las curvas tampoco mostraron diferencias entre las pendientes (ANCOVA, F 0.05, 2, 19), y aunque se encontraron mas juntas si hubo diferencias en su posición (ANCOVA; F 0.05, 2, 22). Así, el mejor crecimiento correspondió al ambiente Thalassia-arena, seguido por Arena y Thalassia, mientras que Coral tuvo el menor crecimiento (Fig.1 b).
Cabe señalar que las curvas son el resultado de la combinación de las parámetros de la ecuación de Von Bertalanffy y por lo tanto muestran el crecimiento de los caracoles colocados en los corrales, desde las tallas menores (100-120 mm) hasta la mayor (160-180 mm), observándose una mayor tasa crecimiento en las tallas pequeñas que en los corrales con caracoles más grandes.
Discusión
El incremento en longitud es un parámetro que generalmente se usa para determinar el crecimiento de los organismos. Un ambiente propicio con la cantidad de alimento suficiente, producirá en el caracol un incremento en la longitud de la concha.
Se ha señalado que S. gigas es microherbívoro, que se alimenta principalmente de epífitos de Thalassia testudinum, detrito de pastos y macroalgas y en los arenales, de microfitobentos (Alcolado 1976). Sin embargo, al considerar los incrementos directos de la concha, no está claro cómo es el crecimiento en cada una de esas áreas, ya que si el alimento principal son los epífitos de Thalassia, entonces se esperaría que los corrales ubicados en ese ambiente o en Thalassia-arena tuvieran un crecimiento mayor en longitud, que los caracoles de áreas sin vegetación; sin embargo, este aspecto no fue evidente ni en PG ni en BCH. No obstante, al considerar las curvas de crecimiento de todas las tallas, se observó que los caracoles crecieron mejor en Thalassia en Punta Gavilán y en Thalassia-arena en Banco Chinchorro. Las curvas marcan la tendencia de la población confinada en los corrales, considerando la tasa de crecimiento de las poblaciones de cada corral, incluyendo a los caracoles pequeños y los grandes, lo que brinda una mejor idea del crecimiento poblacional.
Los caracoles juveniles tienen un crecimiento acelerado en los primeros meses de vida, (Berg 1976, de Jesús-Navarrete 1997) y esto fue evidente en el experimento, ya que el incremento marginal de la concha fue mayor en las tallas más pequeñas: 100-120 y 121-140 mm de longitud de concha.
Los incrementos promedio mensuales de 3.21 mm/mes en Punta Gavilán y 2.30 mm/mes en Banco Chinchorro, son similares a los encontrados en otros sitios del Caribe, como incrementos de 3.3 mm/mes en las aguas cubanas (Alcolado 1976), de 4.16 mm por mes en Islas Vírgenes (Randall 1964), de 4.5 mm/mes en Florida (Brownell 1977), o de 0.139 mm/día (4.17 mm/mes) y 0.058 mm/día (1.74 mm/mes) en caracoles juveniles silvestres y criados en cautiverio, en Bahamas, encontrándose una diferencia estacional en el crecimiento (Ray y Stoner 1994). Los crecimientos registrados en este trabajo fueron menores a los encontrados por Gibson et al. (1983) de 7.2 mm/mes en Belice, o de 15 mm/mes en condiciones óptimas en Venezuela (Weil y Laughlin 1984). En Punta Gavilán, utilizando técnicas de marcaje y recaptura, se midió un crecimiento promedio de juveniles de 10 mm/mes (n=10, ds=0.36 mm) (de Jesús-Navarrete y Oliva Rivera 1997).
El análisis de varianza mostró diferencias significativas en el crecimiento de los caracoles entre los ambientes y esto refuerza la idea de que el crecimiento de los caracoles depende mucho de la cantidad de alimento disponible en cada sitio, alimento que es encontrado en ambientes con vegetación y que marca un período en el desarrollo ontogenético del caracol, ya que se ha señalado que los juveniles >60 mm de longitud de concha, e incluso los de 180 mm dependen de las praderas de Thalassia para su alimentación y protección (Ray y Stoner 1994).
Es posible que el traslado de caracoles de áreas con alta densidad a zonas despobladas, tal y como lo han propuesto las cooperativas pesqueras de Quintana Roo, tenga un efecto similar y entonces habría que seleccionar aquellos sitios con biomasas de pastos similares a las del hábitat natural de tal manera que no se afecte el crecimiento. Sin embargo, es necesario recabar mayor información sobre la cantidad de alimento disponible en los sedimentos de los arenales y áreas de parches coralinos, principalmente del microfitobentos.
Las densidades utilizadas en este experimento (0.40 ind/m2) no afectaron el crecimiento, debido a que no se presentó competencia por espacio ni alimento, dando como resultado un crecimiento adecuado, ya que los caracoles exhiben un crecimiento lento a densidades elevadas, 4-8 caracoles/m2, pero a densidades de un caracol/m2 no se afecta el crecimiento (Ray y Stoner 1994). Así, el crecimiento de los juveniles depende de la cantidad de alimento disponible y por lo tanto se obtiene mayor crecimiento en zonas con pastizales como los ambientes Thalassia y Thalassia-arena, tanto en Punta Gavilán como en Banco Chinchorro. Las curvas de crecimiento integran las tendencias del crecimiento de cada ambiente y son los que se debieran considerar para decidir si el crecimiento fue adecuado o no. Aparentemente, incremento marginal de la concha indica que el crecimiento fue mejor en ambientes sin vegetación, como Arena en Punta Gavilán y Coral en Banco Chinchorro, pero las curvas de crecimiento muestran que a nivel poblacional hubo un mayor crecimiento en los ambientes con vegetación, por lo que en caso de hablar de repoblación esos deben ser los ambientes que se deban considerar para ubicar a los caracoles.
Materiales y métodos
Los estudios se realizaron dentro de la laguna arrecifal en Punta Gavilán (PG) (18° 21' 011" N, 87°48' 26" W) y en Cayo Centro en Banco Chinchorro (BCH) (18° 36' 12" N, 87° 18' 28" W). El experimento se realizó de octubre de 1993 a marzo de 1994.
Cuatro corrales circulares de malla de acero galvanizado de 1.0 cm de luz de malla y 0.30 m de altura fueron instalados en cada ambiente. Cada corral tuvo un área de 50 m², con una división interna de 25 m², lo cual permitió distribuir homogéneamente a los caracoles y tener una réplica por corral. Los ambientes seleccionados fueron: a) Thalassia, con pasto marino abundante (Thalassia testudinum), b) Thalassia-arena pastos con arena c) Arena y d) Coral. La biomasa de pastos en Thalassia fue de 23.4 gramos de peso seco (g. p. s./m2, y de 16.26 g. p. s. /m2 en Thalassia-arena para Punta Gavilán. En Banco Chinchorro el ambiente Thalassia estuvo dominado por las algas verdes Halimeda incrasata y H. monile, con biomasas de 83.8 g. p. s. /m2 y de 54.3 g. p. s. /m2 de las mismas algas en Thalassia-arena. Los corrales se anclaron uniformemente al fondo con grapas de acero de 60 cm de longitud, para evitar la pérdida de organismos. En Punta Gavilán, los corrales se colocaron entre 0.60 m y 3.0 m de profundidad, mientras que en Banco Chinchorro la profundidad varió de 1.5 a 6.5 m.
En cada corral se introdujeron 20 caracoles, 10 en cada sección del corral, de las siguientes tallas: 1) 100-120 mm 2) 120-140 mm 3) 140-160 mm y 4) 160-180 mm de longitud de concha.
Los caracoles se capturaron en zonas adyacentes y se midieron con un vernier con una precisión de un mm. Antes de introducirlos a los encierros se marcaron con una etiqueta de plástico numerada, que se ató a la espira con una cinta del mismo material.
El crecimiento de los caracoles se evaluó de dos maneras: primero, por el incremento directo de la longitud de la concha y segundo, con la curva de crecimiento de Von Bertalanffy a través del método de Gulland-Holt, dentro del paquete ELEFAN (Gayanilo, et al. 1988).
Para el incremento directo de la concha, los caracoles se midieron mensualmente, y se obtuvo el promedio global del incremento de todos los caracoles de los cuatro corrales. Se consideró sólo a los caracoles que sobrevivieron a todo el experimento. Sin embargo, los caracoles muertos fueron reemplazados por otros de talla similar, para mantener constante la densidad.
Con los datos del incremento de la concha de todos los caracoles, incluso los que repusieron a los muertos, se realizó un análisis del crecimiento para cada ambiente, aplicando el método Gulland-Holt, que considera los datos de marcaje recaptura, con los que se obtuvieron los parámetros de crecimiento de la ecuación de von Bertalanffy para cada corral.
Para determinar diferencias estadísticas significativas entre los incrementos la longitud de la concha promedio, entre el tipo de ambiente, sitio y mes, se aplicó un ANOVA de tres vías, con efectos fijos. Cuando las diferencias fueron significativas (p< 0.05), se realizó un análisis a posteriori, con el método de Tukey.
Para determinar si había diferencias estadísticamente significativas entre la pendiente y altura de cada una de las curvas de crecimiento, de cada ambiente y cada sitio, se transformaron las curvas de crecimiento de Von Bertalanffy, a ecuaciones lineales y se aplicó un análisis de covarianza.
Agradecimientos
A José Oliva, Martín Domínguez, Alejandro Medina y Mauro Góngora por su colaboración en la recolecta de información, a Eloy Sosa por su ayuda en el análisis estadístico. El Centro de Investigaciones de Quintana Roo financió este trabajo. A Minerva Arce y Jacobo Schmitter por sus comentarios al manuscrito.
Resumen
Se estudió la tasa de crecimiento de caracoles cultivados en corrales en cuatro ambientes diferentes. El cultivo se realizó de octubre de 1993 a marzo de 1994. 16 corrales de 50 m² de área cada uno, cuatro por ambiente, fueron colocados en los siguientes ambientes: Thalassia, Thalassia-arena, Arena y Coral, todos dentro de la laguna arrecifal en Punta Gavilán y Banco Chinchorro. En cada ambiente se introdujeron 20 caracoles de las siguientes tallas: 1) 100-120, 2) 120-140, 3) 140-160 y 4) 160-180 mm de longitud de concha. Los caracoles se midieron mensualmente con un vernier con una precisión de un mm. El crecimiento se evaluó por dos métodos: a) el incremento marginal promedio de la concha y por el método de Gulland-Holt. En el primer método, el ambiente Arena mostró el mayor crecimiento (3.21 ± 0.26 mm/mes) en Punta Gavilán, mientras que en Banco Chinchorro, el mayor crecimiento ocurrió en coral (2.31 ± 0.44 mm/mes). Con el segúndo método la mayor longitud asintótica en Punta Gavilán se midió en Thalassia-arena (287.5 mm), mientras que en Banco Chinchorro, la mayor longitud asintótica se midió en Arena (318.1 mm). Existieron diferencias significativas en el crecimiento entre los sitios y eso muestra que el crecimiento de los juveniles se relaciona con la cantidad de alimento disponible el cual fue mayor en Thalassia y Thalassia-arena. Sin embargo se requiere evaluar la cantidad de alimento, principalmente microfitobentos en los ambientes sin vegetación como Arena y Coral.
Referencias
Alcolado, P. M. 1976. Crecimiento, variaciones morfológicas de la concha y algunos datos biológicos del "Cobo" Strombus gigas L. (Mollusca, Mesogastropoda). Acad. Cienc. Cuba. Ser. Oceanol. 34: 1-36. [ Links ]
Berg, C. J. Jr. 1976. Growth of the queen conch Strombus gigas, with a discussion of the practicality of its mariculture. Mar. Biol. 34: 191-199. [ Links ]
Brownell, W.N. 1977. Reproduction, laboratory culture and growth of Strombus gigas, S. costatus and S. pugilis in Los Roques Venezuela. Bull. Mar. Sci. 27: 668-680. [ Links ]
Creswell, L. 1984. Ingestion, Assimilation and Growth of juveniles of the queen conch Strombus gigas Linné Fed experimental diets. J. Shellfish Res. 4: 23-30. [ Links ]
Cruz, R. 1984. Avances en la experimentación de producción masiva de caracol en Quintana Roo, México. Proc. Gulf and Caribb.Fish Inst. 37: 12-20. [ Links ]
Chávez, E. A. & F. Arreguín-Sánchez. 1994. Simulation modelling for conch fishery management, p. 125-136. In R. S. Appeldoorn y Q. Rodríguez (eds). Queen Conch Biology, Fisheries and Mariculture. Fundación Científica Los Roques, Caracas Venezuela. 169-189. [ Links ]
Davis, M. 1994. Mariculture techniques for queen conch (Strombus gigas L.): Egg mass to juvenile stage, p. 231-252. In Appeldoorn, R.S. y B. Rodriguez (eds). Queen Conch Biology, Fisheries and Mariculture. Fundación Científica los Roques, Caracas, Venezuela 356 p. [ Links ]
de Jesús-Navarrete, A. 1997. El crecimiento del caracol rosado Strombus gigas y la posibilidad de cultivo en Quintana Roo, p. 73-80 In Caro, C. C. & E. Sosa (eds). La acuicultura en el sureste de México. El Colegio de la Frontera Sur, 192 p. [ Links ]
de Jesús-Navarrete, A. & J. Oliva-Rivera. 1997. Densidad, crecimiento y reclutamiento del caracol rosado Strombus gigas en Punta Gavilán, Quintana Roo, México. Rev. Biol. Trop. 42: 797-801. [ Links ]
Gayanilo, F. C. Jr., M. Soriano & D. Pauly. 1998. A draft guide to the complete ELEFAN. ICLARM Contribution 435, Manila, Philippines 65 p. [ Links ]
Gibson, J., S. Strasdine & K. González. 1983. The status of conch industry of Belize. Proc. Gulf and Caribb. Fish Inst. 35: 99-107. [ Links ]
Randall. J. E. 1964. Contribution to the Biology of the Queen Conch Strombus gigas. Bull. Mar. Sci. 14: 246-295. [ Links ]
Ray, M. & A. W. Stoner. 1994. Experimental analysis of growth and survivorship in a marine gastropod aggregation: balancing growth with safety in numbers. Mar. Ecol. Prog. Ser. 105: 47-59. [ Links ]
Rodríguez, L. A. & J. Ogawa. 1990. Cría de larvas de caracol rosado Strombus gigas L. en dos sistemas diferentes. Proc. Gulf and Caribb. Fish Inst. 43th: Abstract. [ Links ]
Sosa-Cordero, E., A. Medina Quej, A. Ramírez-González, M. Domínguez-Viveros & W. Aguilar. 1993. Invertebrados marinos explotados en Quintana Roo, p. 709-734. In Salazar-Vallejo, S. & N. E. González (eds). Biodiversidad Marina y Costera de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad-Centro de Investigaciones de Quintana Roo, Chetumal, Quintana Roo, 865 p. [ Links ]
Stoner, A. W. & J. V. Sandt. 1988. Transplanting as a test procedure before large-scale out planting of juvenile queen conch. Proc. Gulf and Caribb. Fish. Inst.40: Abstract. [ Links ]
Stoner, A. W. & J. M. Waite. 1990 Distribution and behavior of Queen Conch Strombus gigas relative to seagrass Standing Crop. Fish. Bull. 88: 573-585. [ Links ]
Stoner, A.W. & M. Ray. 1996. Queen conch, Strombus gigas, in fished an unfished locations of the Bahamas: Effects of a marine fishery reserve on adults, juveniles, and larval production. Fish. Bull. 94: 551-565. [ Links ]
Weil, E. & R. Laughlin. 1984. Biology, population dynamics and reproduction of the queen conch Strombus gigas in the Archipelago Los Roques National Park. J. Shellfish Res. 4: 45-62. [ Links ]
1 Departamento de Pesquerías Artesanales. El Colegio de la Frontera Sur. Unidad Chetumal. A. P. 424 Chetumal Q. Roo México. C.P. 77000, Fax 52+983 20447; alberto@ecosur-qroo.mx

