










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.1 San José Mar. 2001
J.B. Escamilla1, E. Suárez-Morales 2 y R. Gasca 2
Recibido 9 II-2000. Corregido 31-VII-2000. Aceptado 19-IX-2000
Abstract
Zooplankton was surveyed in a tropical lagoon system of the northern coast of the Yucatan Peninsula in high tide, December (1998) and low tide, March 1999 (northerlies season). Zooplankton biomass was measured, zooplankters were counted, and copepods were identified and quantified. Despite the fact that both months were influenced by winds from the North, they showed a different salinity gradient which developed a particular structure of the zooplankton community. Biomass tended to be accumulated in certain areas apparently because of the high residence time of water in Chelem, the forcing effect of the northerlies, and of the tidal current. Biomass values suggest a relatively high secondary production when compared with other systems of the Yucatan Peninsula. The distribution of the copepods Acartia lilljeborgii and A. tonsa is related to saline conditions and tidal flow. The overall faunistic and hydrologic data suggest that even during a single climatic season, the zooplankton community shows strong changes due to mesoscale hydrological processes.
Key words: zooplankton, tides, copepods, tropical.
El zooplancton de extensos sistemas costeros de la península de Yucatán permanece desconocido. Uno de ellos es Chelem, Yucalpetén, en la costa norte de la península de Yucatán, México. Herrera-Silveira et al. (1999) han estudiado aspectos ecológicos-hidrológicos de este sistema. En este estudio se aportan los primeros datos acerca del zooplancton de Chelem y se hacen consideraciones acerca de su relación con variables hidrológicas. Esto se hace a escala reducida al comparar datos de dos meses de una misma época climática, pero con condiciones de marea distintas.
Este complejo lagunar se localiza entre los 21° 17' N y los 89° 40' W, litoral norte de la Península de Yucatán. Tiene fuerte influencia marina (Herrera-Silveira y Ramírez-Ramírez 1998) y se divide en varias secciones fisiográficas (ver Fig. 1). El flujo transversal de agua está influenciado por las mareas (Flores et al. 1995).
Se recolectó el zooplancton durante dos meses del periodo de nortes (invierno) en seis estaciones en diciembre 8, 1998 y siete el 9 de marzo, 1999 (Fig. 1). Las muestras de diciembre corresponden a marea baja franca (salida de agua hacia el mar), las de marzo son de marea alta (entrada de agua marina). El zooplancton se obtuvo mediante arrastres superficiales de 7 mins. con una red estándar con malla filtrante de 0.33 mm y equipada con un flujómetro. Se midió la biomasa (peso húmedo) del zooplancton (mg/m3). Se determinó la composición general del zooplancton y la específica de los copépodos y su densidad (org./100m3). Se midieron temperatura, salinidad y otros factores (Cuadro 1).
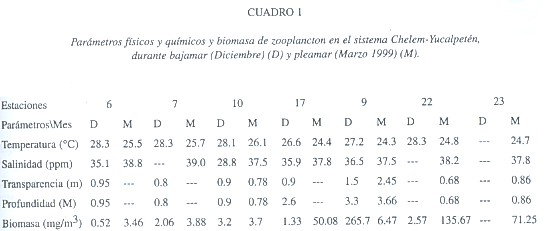
En bajamar el agua salía de la laguna hacia el mar, y en pleamar el agua de mar entraba a la laguna y ello se reflejó en la distribución de la salinidad. La salinidad más baja fue registrada, en ambos casos, en la zona más interna de la laguna, que recibe aportes freáticos de agua dulce (Herrera-Silveira 1996). En diciembre (bajamar) se observó un gradiente salino, de menor (28 PSU) a mayor (ca. 36) y hacia la zona marina; este patrón se modifica en marzo, con valores más altos y homogéneos, evidenciando una mayor influencia marina relacionada con la marea entrante.
Los valores promedio de biomasa fueron similares en ambos meses (45.9 mg/m3 en diciembre vs. 39.21 mg en marzo); su distribución muestra diferencias entre la ciénega y la laguna, la primera (est. 17, 22, 23) consistentemente con mayores biomasas (Cuadro 1). Las diferencias generales en biomasa no fueron significativas (ANOVA: F=0.0009, p=0.9977). Durante bajamar la mayor biomasa se encontró cerca de la boca, consecuencia de la salida de agua y plancton hacia el frente marino. En marzo tres factores favorecen la mayor biomasa en la zona oriental: 1) el efecto de acumulación del zooplancton debido a la influencia de los vientos del norte (Suárez -Morales y Gasca 1996); 2) alto tiempo de residencia del agua en Chelem (Herrera-Silveira et al. 1999); 3) el flujo de la marea hacia la zona costera. Las mareas son uno de los principales mecanismos transportadores de plancton en las zonas costeras y arrecifales (Suárez-Morales y Rivera-Arriaga 1998). En general, los valores locales de biomasa son muy superiores (50-500%) a los encontrados en sistemas costeros del litoral oriental de la península (Gasca et al. 1994), lo que sugiere una productividad secundaria relativamente alta.
La densidad promedio global del zooplancton para bajamar fue de 30 913 org./100 m3; aumentó en pleamar (84 131 org./100 m3). La cifra dentro del cuerpo lagunar fue de 6 103 org./100 m3 y de 94 415 org./100 m3, repectivamente. En ambos casos predominaron los copépodos, formaron el 89.8 % del número total de zoopláncteres en bajamar y el 72.2 % en pleamar. Las larvas de crustáceos decápodos formaron el 3.12 % y el 10.21 %, respectivamente. Los quetognatos, representados por Sagitta hispida Conant, 1895 constituyeron el 1.26% en diciembre y el 2.29% en marzo. La mayor variabilidad de la densidad del zooplancton en las lagunas costeras de la región parece darse entre las épocas climáticas; las variaciones mensuales suelen ser relativamente débiles en sistemas similares (Gasca & Suárez-Morales, 1994). La estructura de la comunidad es estable en términos de biomasa y composición general, pero está sujeta a variaciones a una escala menor y éstas pueden identificarse mediante el análisis de grupos clave como los copépodos. Por ello, las diferencias en abundancia observadas en este estudio pueden asociarse con la acción de procesos locales de mesoescala, como lo son las mareas.
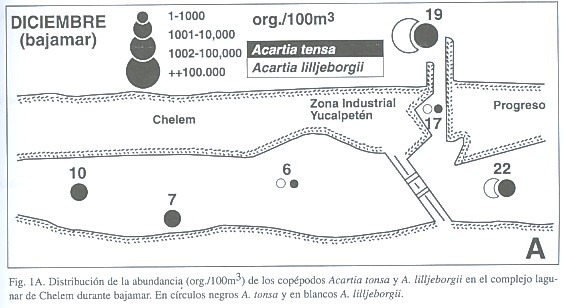
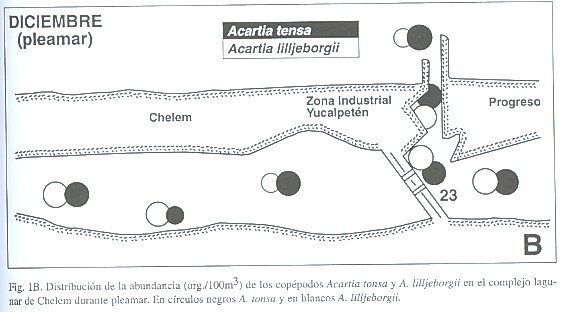
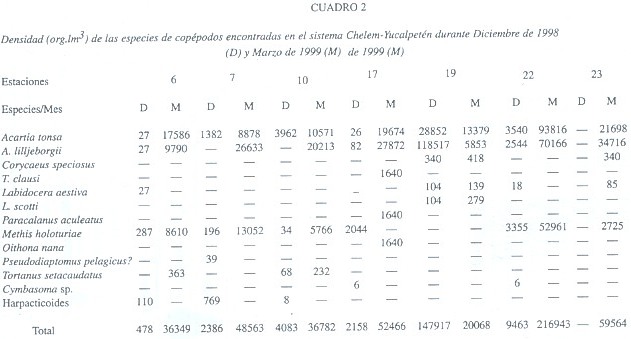
En diciembre el copépodo más abundante fue Acartia lilljeborgii Giesbrecht, 1889, una forma costera (72.78% del total de copépodos), seguido por A. tonsa Dana, 1849 (22.69%), una forma estuarina, y por Metis holothuriae (Edwards, 1891) (3.55%). En marzo las proporciones cambiaron (A. lilljeborgii: 41.65%, A. tonsa: 39.35%, M. holothuriae: 17.52%) (Cuadro 2, Fig. 1). La distribución de estas dos especies en Chelem indica las distintas condiciones salinas que se presentaron en ambos meses estudiados. El gradiente salino de diciembre se caracteriza por la distribución de A. lilljeborgii (en zonas con influencia marina) y de A. tonsa (en zonas más internas de menor salinidad). Al salir el agua por efecto de la bajamar, se producen densidades altas de A. tonsa en la zona nerítica y en la zona interna cercana al canal de comunicación. Al interior de Chelem esta especie tiene densidades moderadas. Por el contrario, A. lilljeborgii es abundante en la zona nerítica, pero su densidad es similar a la de A. tonsa al interior y desaparece de las zonas más internas (ver Fig. 2A), lo que indica condiciones de baja salinidad incompatibles con A. lilljeborgii. En marzo, con la entrada de agua marina en pleamar, A. lilljeborgii llega a ser más abundante que A. tonsa en las partes más internas de la laguna de Chelem (Fig. 2B). Las diferencias de densidad entre baja y pleamar de estas especies en las zonas más internas son estadísticamente significativas (ANOVA de 1 vía, F=13.3, p=0.022 para A. tonsa y F=14.78, p=0.018 para A. lilljeborgii). Desde luego, en las áreas donde la salinidad permite su coexistencia las diferencias no son significativas (p=0.1046 para A. tonsa y p=0.583 para A. lilljeborgii). Esto indica una influencia marina que es más intensa hacia las zonas internas donde transforma la estructura de la comunidad de copépodos. El comportamiento de estas dos especies es similar al encontrado por Suárez-Morales (1994) en otro sistema de la región. El ingreso de A. lilljeborgii a las zonas más internas de los esteros durante la pleamar ha sido reportado por Alvarez-Cadena y Cortés-Altamirano (1990). En Chelem es claro que estas dos especies determinan el perfil del zooplancton e indican cercanamente el comportamiento de la salinidad en el sistema. El seguimiento de su distribución y abundancia permite hacer inferencias acerca de la dinámica de la comunidad del zooplancton.
El primer autor agradece a Luis J. Morales Arjona su apoyo para realizar las recolectas de zooplancton en Chelem. El Instituto Tecnológico de Mérida ofreció su respaldo para que este estudio se desarrollara.
Referencias
Álvarez-Cadena, J. N. & R. Cortés-Altamirano. 1990. Algunos factores físicos y biológicos que afectan las poblaciones naturales de Acartia tonsa y A. lilljeborgii (Copepoda: Acartiidae) en el estero de Urías, Sinaloa, México. Inv. Mar. CICIMAR 5: 69-77. [ Links ]
Flores, S. J., E. Batllori-Sampedro, M. Villasuso & A. Mendoza-Millán. 1995. Marco de referencia para el Manejo de la Zona Costera del Estado de Yucatán. Consejo Estatal de Consultoría Ecológica. Doc. Téc. Núm. 1. 89 p. [ Links ]
Gasca, R. & E. Suárez-Morales. 1994. Zooplankton biomass fluctuations in a Mexican Caribbean bay during a year cycle. Carib. J. Sci. 30: 116-123. [ Links ]
Gasca, R., E. Suárez-Morales & L. Vásquez-Yeomans. 1994. Estudio comparativo del zooplancton (biomasa y composición) en dos bahías del Mar Caribe Mexicano. Rev. Biol. Trop. 42: 595-604. [ Links ]
Herrera-Silveira, J. A. 1996. Salinity and nutrients in a tropical coastal lagoon influenced by groundwater discharges. Hydrobiologia 321: 165-176. [ Links ]
Herrera-Silveira, J. A., M. Martín & V. Díaz-Arce. 1999. Variaciones del fitoplancton en cuatro lagunas costeras del Estado de Yucatán, México. Rev. Biol. Trop. 47(supl. 1): 47-56. [ Links ]
Herrera-Silveira, J. A. & J. Ramírez-Ramírez. 1998. Salinity and nutrients in the coastal lagoons of Yucatan, Mexico. Verh. Internat. Verein. Limnol. 26: 1473-1478. [ Links ]
Suárez-Morales, E. 1994. Copépodos pláncticos de la Bahía de Chetumal, México (1990-1991). Carib. J. Sci. 30: 181-188. [ Links ]
Suárez-Morales, E. & R. Gasca. 1996. Planktonic copepods of Bahía de la Ascensión, Caribbean coast of Mexico: a seasonal survey. Crustaceana 69: 162-174. [ Links ]
Suárez-Morales, E. & E. Rivera-Arriaga. 1998. Zooplancton e hidrodinámica en zonas litorales y arrecifales de Quintana Roo, México. Hidrobiológica 8: 19-32. [ Links ]
1 Instituto Tecnológico de Mérida (ITM), A.P. 9-11, Mérida, Yucatán 97118, México.
2 El Colegio de la Frontera Sur (ECOSUR). Chetumal, Quintana Roo. 77000. México; fax: (983)21666x240; esuarez@ecosur-qroo.mx

