











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.48 no.4 San José dic. 2000
diversidad y similitud fungística
María de la Luz Fierros, José Luis Navarrete-Heredia y Laura Guzmán-Dávalos1
Recibido: 4-VIII-99 Corregido: 24-IV-00 Aceptado: 24-V-00
Abstract
Diversity and similarity of macrofungi of Sierra de Quila, Jalisco, México were analized in three different kinds of vegetation. Fungal diversity in the area is high. The pine-oak and cloud forests, were more diverse in their community structure than the oak forest. Similarity among the three kinds of vegetation was low, there are few species share among them; pine-oak and cloud forests show higher affinity than oak forests. This pattern of similarity is a general condition for others regions with environmental conditions similar to Sierra de Quila.
Key Word Index
diversity, similarity, macrofungi, ecology, Mexico.
La Sierra de Quila es considerada una área silvestre protegida, establecida bajo la categoría de "Zona de Protección Forestal y Fáunica" (Diario Oficial de la Federación, 1982). A pesardeque fue decretada desde hace más de 15 años, no cuenta con un plan de manejo y aunque lo tuviera, por lo general, los hongos pocas veces son tomados en cuenta dentro de los planes de manejo de las áreas protegidas. De ahí que resulte importante la necesidad de resaltar el papel de los hongos dentro del ecosistema. Lamentablemente, en muchas ocasiones la diversidad debe ser medida en términos económicos para poder otorgarle una protección real (Fox, 1993), aunque una justificación suficiente para proteger de igual manera a los hongos es la de mantener la existencia de toda variedad de formas de vida desarrolladas a lo largo de la evolución (Arnolds, 1990).
Por otro lado, en México, son relativamente pocos los trabajos micológicos realizados que incluyan análisis ecológicos. Como antecedente, se tiene a Heredia (1989) en la Reserva de la Biosfera El Cielo, en donde compara la similitud fungística entre los diferentes tipos de vegetación presentes; y el de Cifuentes, Villegas y Pérez-Ramírez (1993), en el Parque Omiltemi en donde analizaron la similitud fungística entre el parque y varias comunidades del país. En la Sierra de Quila, Fierros y Guzmán-Dávalos (1997), realizaron el inventario de los hongos macroscópicos presentes en la zona. Con base en este trabajo, se analizaron algunos aspectos referentes a su distribución de acuerdo a sustrato y tipo de vegetación, así como la diversidad y similitud de hongos entre diferentes asociaciones vegetales.
La Sierra de Quila es una región boscosa situada a unos 100 Km al SO de la ciudad de Guadalajara, Jalisco, México, con una extensión de 15,192 ha. de las 32000 ha. de superficie boscosa. Geográficamente se localiza entre los paralelos 20º12´10" y 20º23´23" de latitud norte y 103º53´25" y 104º11´35" de longitud oeste (Anónimo, 1982). Esta zona montañosa está dentro de la región fisiográfica denominada Neovolcánica. La sierra presenta una altitud que varía de los 1300 msnm en su zona más baja y los 2560 msnm en su punto más alto. Su clima es templado subhumedo con lluvias en verano, en sus partes más altas, mientras que en las partes más bajas es templado semicálido.
De acuerdo a Guerrero (1994) y Guerrero-Nuño y López-Coronado (1997), la Sierra de Quila presenta seis tipos de vegetación de acuerdo al criterio de Rzedowski (1981), de los cuales, los tres siguientes fueron objeto del presente estudio:
Bosque de encino. Se distribuye de manera general entre los 1500 y 1900 msnm. En sus límites más bajos llega a traslaparse con el bosque tropical caducifolio. Se puede encontrar tanto en suelos planos como laderas. Las principales especies son Quercus magnoliifolia Née., Q. resinosa Liebm., Q. gentryi C.H. Müeller, Q. laeta Liebm., Q. castanea Née., entre otros, junto con Agave spp., Calliandra spp., Senna spp., Arctostaphylos pungens H.B.K. y Verbesia spp.
Bosque de pino y encino. Este tipo de vegetación se distribuye en la mayor parte de la sierra, desde los 1500 hasta los 2560 msnm. Los elementos arbóreos que se encuentran son Arbutus xalapensis H.B.K., A. glandulosa Mart. & Gal., Quercus castanea, Q. resinosa, Q. magnoliifolia,Pinus douglasiana Martínez, P. lumholtzii Fern., P. oocarpa Schiede., Alnus spp., entre otros.
Bosque mesófilo de montaña. Se localiza de forma fragmentaria y su composición florística varía de una comunidad a otra. Se le puede encontrar principalmente en cañadas y sobre todo asociado a condiciones de gran humedad y cierta influencia subtropical. Se presenta de los 1950 a los 2560 msnm. Los géneros de árboles más comunes son: Clethra, Prunus, Phoebe, Quercus, Salix, Alnus y Fraxinus.
Materiales y métodos
Se realizaron diversos análisis ecológicos con base en los ejemplares recolectados de julio de 1993 a octubre de 1994, en 16 sitios de muestreo a diferentes niveles altitudinales, desde los 1750 a los 2100 msnm, y se obtuvo un total de 452 ejemplares (Fierros y Guzmán-Dávalos, 1997). Se tomaron en cuenta tanto especies identificadas como aquéllas que no se logró determinar, con el fin de no sesgar información. El grado de significancia para los diferentes tipos de vegetación se evaluó mediante una prueba de t.
El análisis de similitud fungística se realizó por medio del índice de Jaccard, como lo sugiere Kohlmann (1994) cuando se tienen datos de presencia-ausencia ya que excluye coincidencias negativas. Los resultados se presentan a través de fenogramas con el método de ligamiento simple (Crisci y López-Armegol, 1983). Este análisis se realizó tanto para los diferentes tipos de vegetación de la sierra, como para otras localidades del estado y del país.
Resultados
Se tomaron en cuenta un total de 340 especies, pertenecientes a los Myxomycotina (2 especies determinadas), a los Ascomycotina (10 especies: 4 determinadas y 6 no determinadas), a los Basidiomycotina (329 especies: 158 determinadas y 171 no determinadas) (Fierros y Guzmán-Dávalos, 1997). La diversidad ecológica se analizó en los tipos de vegetación mejor representados en la sierra; de ellos, el bosque de pino-encino (BPE) y el bosque mesófilo de montaña (BMM) fueron los que presentaron valores más altos de diversidad, mientras que el bosque de encino (BE) tuvo el valor más bajo (Cuadro 1). Se calculó además la uniformidad (U) en cada tipo de vegetación. En los tres casos el valor de la uniformidad fue alta, pero el que presentó mayor uniformidad fue el bosque de encino (U= 0.972), comparado con el bosque de pino-encino y el bosque mesófilo de montaña (Cuadro 1).
Al comparar la estructura de la comunidad fúngica en los tres tipos de vegetación, el grupo formado por el BPE-BMM (t = 0.138/ 669 gl) no presenta diferencias significativas; sin embargo, si existen diferencias significativas en cuanto a su composición específica. Los grupos formados por BE-BPE (t = 4.92 / 91 gl) y BE-BMM (t = 4.91/ 90 gl), son ambos significativamente distintos (t = 99.5%; 60-120, gl = 2.91-2.86; ![]() , gl = 2.80).
, gl = 2.80).
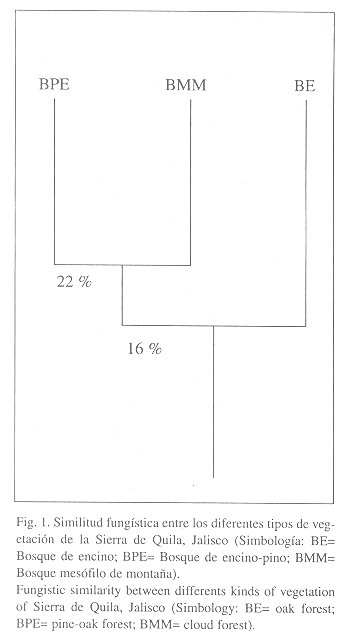
Al analizar la similitud fungística entre los diferentes tipos de vegetación, se observa que en general presentan una similitud muy baja, pero dentro de éstos, los más afines fueron el bosque de pino-encino y el bosque mesófilo de montaña, con un valor de 22% y el bosque de encino en relación conl grupo formado por pino-encino/mesófilo de montaña, obtuvo un valor del 16% (Figura 1).
En cada tipo de vegetación se encontraron especies típicas; dentro del bosque de pino-encino, prosperan de forma abundante las especies micorrízicas, aunque también existen en menor abundancia, humícolas y lignícolas. Algunos hongos comunes en el bosque de pino-encino dentro de la zona de estudio fueron: Amanita muscaria (Sing.) Jenking, A. rubescens (Pers. : Fr.) Gray, Boletus frostii A.W. Russell Collybia spp. Cystoderma spp., Macrolepiota procera (Scop. : Fr.) Singer, Phyllotopsis nidulans (Pers. : Fr.) Singer, Ramaria flava (Fr.) Quél., R. formosa (Fr.) Quél., Suillus brevipes (Peck.) Kuntze, S. granulatus (L. : Fr.) Kuntze, por mencionar algunos. Dentro del bosque mesófilo de montaña, aunque también existen especies micorrízicas, es posible encontrar de forma abundante, debido a la influencia subtropical de este tipo de vegetación, numerosas especies lignícolas, y además por acumulación excesiva de materia orgánica y al alto grado de humedad que presenta esta zona, muchas especies humícolas. En el bosque mesófilo de montaña fueron comunes Amanita cokeri (Gilb. & Künh.) Gilb., Boletellus coccineus (Fr.) Singer, Boletus flammans Dick. & Snell., Polyporus spp., Panus crinitus (L. : Fr.) Sing., Mycena leaina (Berk.) Sacc., Lactarius chrysorreus Fr., Leccinum scabrum (Bull.: Fr.) Gray, Ganoderma curtisii (Berk.) Murr., Paxillus panuoides (Fr. : Fr.) Singer y Xeromphalina tenuipes (Schwein.) A.H. Sm. Lo anterior concuerda de una manera general con lo señalado por Guzmán-Dávalos y Guzmán (1979).
Se detectó también que algunas especies típicas de cierto tipo de bosque, están también presentes en otro, por ejemplo Amanita caesarea, A. muscaria y Laccaria laccata (Scop. : Fr.) Berk. & Br., típicas de pinos, se encontraron en el bosque mesófilo de montaña. Esto puede deberse a que el lugar donde se realizaron los muestreos, presenta características muy peculiares; por un lado su composición florística es muy variable y por otro y el más importante es que en esta zona se encuentra asociada al bosque de pino-encino, hay lugares en que los elementos de ambos llegan a traslaparse, lo cual ocasiona la existencia de especies compartidas
Entre las especies que fueron más abundantes en el bosque de encino se encuentran Amanita fulva Schaeff. : Pers., Amanita gemmata (Fr.) Gill., A. vaginata (Bull. : Fr.) Pat., Lactarius piperatus (L. : Fr.) Gray y L. vellereus (Fr.) Fr., y algunas especies de Collybia. Algo interesante de esta zona es la presencia de especies como Dacryopinax spathularia (Schwein.) G.W. Martin, Panus crinitus, Ganoderma curtisii (Berk.) Murr., que son típicas de zonas tropicales y subtropicales, lo que denota la presencia de una influencia tropical en el bosque de encino de la sierra.
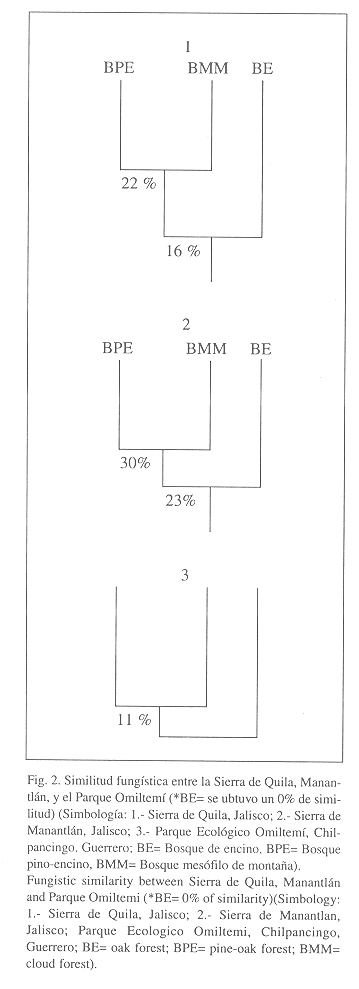
Con el fin de analizar si el patrón de baja similitud observado en la Sierra de Quila, estaba influenciado por factores externos, lo que daría un patrón exclusivo para la zona o si era una condición general, se analizaron otras regiones con tipos de vegetación similares, ya que se esperaba que el bosque de encino tuviera una afinidad mayor con el bosque pino-encino y el bosque mesófilo de montaña, debido a la presencia de varias especies de encinos en ambos tipos de vegetación. Se examinaron, del estado de Jalisco, la Sierra de Manantlán, de acuerdo a los datos tomados de Téllez, Guzmán-Dávalos y Guzmán (1988) y del estado de Guerrero, el Parque Ecológico Estatal Omiltemi, que pertenece a la vertiente del Pacífico, con datos de Cifuentes, Villegas y Pérez-Ramírez (1993). Los resultados obtenidos del análisis de similitud fungística, siguen un patrón semejante al encontrado en la Sierra de Quila (Fig. 3).
En el caso de Manantlán, el bosque mesófilo de montaña y el de pino-encino presentaron una similitud del 30% y el bosque de encino en relación conl grupo de pino-encino y mesófilo, obtuvo un valor del 23%. Este patrón se repite en Omiltemi, que presentó una similitud del 11 % para los bosques de pino-encino/mesófilo, pero para el bosque de encino se tenían muy pocas recolectas en relación con los otros tipos de vegetación, por lo que no fue suficientemente representativo, obteniendo un valor de cero (Fig. 3).
Discusión y conclusiones
La diversidad fúngica en la Sierra de Quila, es elevada, el grupo de hongos que se desarrollan en el bosque de pino-encino y en el bosque mesófilo de montaña, significativamente más diverso en cuanto a su estructura de comunidad, en relación conl otro tipo de vegetación analizado. Cada tipo de vegetación, dentro de los bosques templados y subtropicales, presenta una micobiota característica y en realidad son muy pocas las especies compartidas con otro tipo de vegetación. En cuanto a la menor diversidad observada en BE puede deberse a que la localidad presenta un grado de perturbación mayor que los otros tipos de vegetación, lo que provoca un menor crecimiento de los hongos (41 especímenes recolectados), o bien a la intensidad de muestreo, ya que en este bosque se recolectó un menor número de veces (Fierros y Guzmán-Dávalos, 1997). Sería conveniente buscar en zonas de encino que se encuentre mejor conservadas, para poder realizar un análisis más confiable y comparar si el patrón presentado en cuanto a la diversidad de especies se mantiene en este caso y para este tipo de vegetación en general, o si el resultado fue influenciado por factores externos.
Para el bosque de pino-encino y el bosque mesófilo de montaña, el número de especies recolectadas es casi tres veces mayor que la del bosque de encino, y por lo tanto hace que estas comunidades sean más complejas en cuanto a su estructura. Por otro lado, hay especies que llegan a predominar en determinado tipo de vegetación, como es el caso de Panus crinitus (L. : Fr.) Singer, Favolus brasiliensis Fr. o Laccaria laccata (Scop. : Fr.) Berk. & Br. para el bosque mesófilo de montaña, y Suillus granulatus (L. : Fr.) Kuntze, Amanita muscaria (Singer) Jenking o Russula foetens Pers. : Fr. para el bosque de pino-encino.
De acuerdo a Magurran (1989), una alta uniformidad puede reflejar una alta diversidad, ya que la especies son iguales o virtualmente iguales en abundancia. Pero en este caso, el hecho de que el bosque de encino presentara el valor más alto puede explicarse en el sentido de que se recolectaron pocas especies en relación con los otros tipos de vegetación y además que la mayor parte de las especies solamente están representadas por un ejemplar, lo que provoca que su uniformidad sea elevada.
En lo referente a la similitud, los bajos valores presentados pueden interpretarse si se toma en cuenta las características físicas y su composición fungística. En el caso del bosque mesófilo de montaña, cuya característica principal es la heterogeneidad en su composición florística y distribución fragmentaria de manera general para el país (Rzedowski, 1981), y para el caso de la propia sierra (Guerrero-Nuño y López-Coronado, 1997), da como consecuencia que esta condición heterogénea se vea reflejada en su composición fungística, lo que ocasiona un bajo número de especies compartidas entre este tipo de vegetación y el bosque de pino-encino.
Sin embargo, también se detectó que algunas especies típicas de cierto tipo de bosque, están presentes en otro, por ejemplo Amanita caesarea, A. muscaria y Laccaria laccata, típicas de pinos, encontraron también en el bosque mesófilo de montaña. Esto puede deberse a que el lugar donde se realizaron los muestreos, presenta características muy peculiares; por un lado su composición florística es muy variable y por otro y el más importante es que esta zona se encuentra asociada al bosque de pino-encino, hay lugares en que los elementos de ambos llegan a traslaparse, lo cual ocasiona la existencia de especies compartidas.
En lo referente al análisis realizado con las diferentes localidades del estado y el país, de manera general, todos los lugares analizados presentaron una similitud muy baja entre estos tres tipos de vegetación, aunque el grupo formado por el bosque de pino-encino/bosque mesófilo de montaña, presentó valores de similitud más elevados. En Manantlán, Jalisco, se obtuvo el valor más alto, seguido de la Sierra de Quila, Jalisco y por último Omiltemi, Guerrero. En conclusión, el patrón de similitud observado es general para las áreas con tipos de vegetación similares a los estudiados en otras partes del país.
Este patrón de distribución en diferentes tipos de vegetación, se ha comunicado con anterioridad por Mueller y Halling (1995) en su estudio sobre la evidencia de la alta diversidad de Agaricales en bosques neotropicales; estos autores señalan la existencia de una distribución definida en la composición de especies entre diferentes regiones geográficas de América y en realidad son pocos los táxones que resultan ser cosmopolitas.
Agradecimientos
Se agradece a las autoridades de la Dirección General Académica de la Universidad de Guadalajara las facilidades otorgadas para realizar este trabajo a través del Programa de Motivación a la Investigación de Estudiantes Sobresalientes. A Julieta Carranza por la revisión crítica de éste manuscrito.
Resumen
Se analizó a diversidad y similitud de los hongos macroscópicos de la Sierra de Quila, Jalisco, México en tres tipos de vegetación. En general la diversidad fúngica en la zona fue elevada. Los bosques de pino-encino y mesófilo de montaña resultaron ser significativamente más diversos en cuanto a su estructura de comunidad en relación conl bosque de encino. La similitud fungística entre los tres tipos de vegetación fue baja y son pocas las especies compartidas entre sí, resultando más afines el bosque de pino-encino y el mesófilo de montaña. Este patrón de similitud es una condición general para otras regiones que presentan características similares a las de la zona estudiada.
Referencias
Anónimo, 1982. "Secretaria de Agricultura y Recursos Hidráulico". Diario Oficial 373 (25): 67-68. [ Links ]
Arnolds, E., 1990. Mycologist and nature conservation, p. 243-264. In D.L. Hawksworth, (ed.). Frontiers in Mycology. CAB International, Wallingford [ Links ]
Cifuentes, J., M. Villegas y L. Pérez-Ramírez, 1993. Hongos macroscópicos, p. 59-126. In I. Luna-Vega & J. Llorente (eds.) Historia natural del Parque Ecológico Estatal Omiltemi, Chilpancingo Guerrero, México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad-Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Crisci, J. V. y M. F. López-Armegol, 1988. Introducción a la teoría y práctica de la taxonomía numérica. OEA, Washington, D.C., 132 p. [ Links ]
Fierros, M. L. y L. Guzmán-Dávalos, 1997. Inventario preliminar de los hongos macroscópicos de la Sierra de Quila, Jalisco, México. Bol. Inst. Bot. (Universidad de Guadalajara) Época 3, 3(1-3): 129-142.
Fox, F. M. 1993. Tropical fungi: their commercial potencial, p. 253-263. In S. J. C. Isacc, Franklandd, R. Watling and A. J. S. Whalley (eds.). Aspects of tropical mycology. Cambridge University Press, Cambridge [ Links ]
Guerrero, J. J., 1994. Contribución al conocimiento de la vegetación y flora de la Sierra de Quila, Jalisco, México. Tesis de Licenciatura. División de Ciencias Biológicas y Ambientales, Universidad de Guadalajara, Guadalajara, México. [ Links ]
Guerrero-Núño, J. J. y G. A. López-Coronado, 1997. La vegetación y la flora de la Sierra de Quila. Universidad de Guadalajara, Guadalajara, 134 p. [ Links ]
Guzmán-Dávalos L. y G: Guzmán, 1979. Estudio ecológico comparativo entre los hongos (macromicetos) de los bosques tropicales y los de coníferas del sureste de México. Bol. Soc. Mex. Mic. 13: 89-125. [ Links ]
Heredia, G., 1989. Estudio de los hongos de la Reserva de la Biosfera El Cielo, Tamaulipas. Consideraciones sobre su distribución y ecología de algunas especies. Act. Bot. Mex. 7: 1-18 [ Links ]
Kohlmann, B., 1994. Algunos aspectos de la taxonomía numérica y sus usos en México 423-438 pp. In J. Llorente. y I. Luna (compiladores). Taxonomía biológica. UNAM-Fondo de Cultura Económica, México, D.F. [ Links ]
Morales, R: y C. M. MacFarland, 1980. Compendio sobre la metodología para la planificación de áreas silvestres. CATIE, Turrialba, 120 p. [ Links ]
Mueller, G. M. y R. E. Halling. 1995. Evidence for High Biodiversity of Agaricales (Fungi) in Neotropical Montane Quercus Forest. 303-312 pp. In Churchill, S. P., H. Balsev, E. Forero & J. L. Luteyn (eds.). Biodiversity y Conservation of Neotropical Montane Forests. The New York Botanical Garden, Nueva York. [ Links ]
Rzedowski, J., 1981. La Vegetación de México. Limusa, México, D. F., 342 p. [ Links ]
Téllez, C., L. Guzmán-Dávalos y G. Guzmán, 1988. Contribución al conocimiento de los hongos de la Reserva de la Biosfera de la Sierra de Manantlán, Jalisco. Rev. Mex. Mic. 4: 20-24. [ Links ]
CUADRO 1.
Diversidad (H´), uniformidad (U) y número de especies en los diferentes tipos de vegetación. Diversity (H´), uniformity (U) and species number in different kinds of vegetation.
| Tipos de vegetación | | | |
| Bosque de encino | | | |
| Bosque pino-encino | | | |
| Bosque mesófilo de montaña | | | |
1 Departamento de Botánica y Zoología, Universidad de Guadalajara, Apartado Postal 139, C.P. 45110, Zapopan, Jalisco, México. Fax (523)6820003. mfierros@maiz.cucba.udg.mx

