










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.4 San José Dec. 2000
José Rolando Bastida-Zavala y Sergio I. Salazar-Vallejo1
Recibido: 9-IX-99 Corregido: 12-V-00 Aceptado: 16-VI-00
Abstract
From the study of more than 450 specimens of Hydroides and Serpula, 12 species from the Grand Caribbean Region were identified and characterized. Eight species were collected along the shores of the Yucatan Peninsula and five were found in other localities in the Gulf of Mexico, seven others are from Cuba and comments on type specimens of five species are also included. One morphometric analysis made on Hydroides mucronatus Rioja and Hydroides cf. mucronatus, indicated several differences among them. Comments on all species are also included
Key words
Caribbean Sea, Morphometric Variation, Polychaetes, Serpulids, Taxonomy
El género Hydroides Gunnerus es el que presenta más especies en la familia Serpulidae (más de 100). Para la diferenciación de las especies del género, resulta muy importante la forma y ornamentación del opérculo y ya existe una propuesta de estandarización en su terminología (ten Hove 1990).
El desarrollo del opérculo de Hydroides y su duplicidad ha llamado la atención desde hace mucho tiempo. Ichikawa & Takagaki (1942) analizaron casi 10 000 ejemplares de H. ezoensis y mostraron que menos del 10% tenían opérculos funcionales duplicados; el resto tenía sólo un opérculo mayor del lado derecho o del lado izquierdo. Otras observaciones similares, aunque con menos ejemplares fueron hechas por Rioja (1919, 1944) y por de León-González (1990) con especies del mismo género del Atlántico de España y del Pacífico mexicano, respectivamente. En sus intensas investigaciones científicas en México, el Dr. Rioja describió siete especies del género Hydroides (Rioja 1941, 1942, 1958); sus colecciones parecían haberse perdido (Salazar-Vallejo 1989), aunque Vivianne Solís (ICML-UNAM) recuperó sus materiales y pronto sabremos su estado (F. Solís com. pers. 1999).
Las relaciones filogenéticas entre especies del género no han sido analizadas a profundidad; sin embargo, contamos con un análisis gráfico realizado por Pillai (1972). Aunque el estudio no abarcaba todas las especies descritas hasta ese entonces, el esfuerzo permitió obtener una idea de las relaciones entre las especies. Es menester realizar una evaluación objetiva de la propuesta de Pillai (1972) y formar a su vez una nueva propuesta de las relaciones entre las especies del género.
El género Serpula Linnaeus 1758 fue el primero en ser descrito y luego fue usado para el nombre de la familia. A Risso (1826) se le debe atribuir la autoría de la familia por ser el primero en reconocer como agrupación natural a los serpúlidos (ver argumentación en Bastida-Zavala & Salazar-Vallejo en prep.).
En este trabajo se analizaron más de 450 ejemplares, de 41 localidades del litoral oriental de México y de siete localidades de Cuba (Apéndice 1). Resultaron 12 especies de serpúlidos de los géneros Hydroides y Serpula. En la Fig. 1 y 2 de un trabajo previo se pueden ubicar los sitios citados en el texto (Bastida-Zavala & Salazar-Vallejo en prep.). Se anexan breves comentarios del material tipo de cinco especies más descritas de Estados Unidos, Jamaica y Puerto Rico, los cuales están depositados en el American Museum of Natural History (AMNH) y en el National Museum of Natural History, Smithsonian Institution (USNM).
Materiales y métodos
El catálogo de Hartman (1959) permitió ubicar más fácilmente los nombres. Por su parte, los listados de especies de la región del Gran Caribe de Perkins & Savage (1975), Salazar-Vallejo (1996) y Perkins (1998) facilitaron la selección de referencias. La metodología y las consideraciones morfométricas usadas aquí se detallaron en un trabajo previo (Bastida-Zavala & Salazar-Vallejo en prep). El material recolectado se depositó en la colección de referencia de ECOSUR (Clave INE: QNR.IN.021.0497). Los ejemplares de Cuba están depositados en el Instituto Oceanológico de Cuba (IOC); otros materiales están en la colección de poliquetos del Instituto de Ciencias del Mar y Limnología (ICML-UNAM), de la Universidad Autónoma de Nuevo León (UANL) y de la Universidad Veracruzana (UV).
Resultados y discusión
Serpulidae Risso, 1826
Serpulinae Risso, 1826
Hydroides Gunnerus, 1768
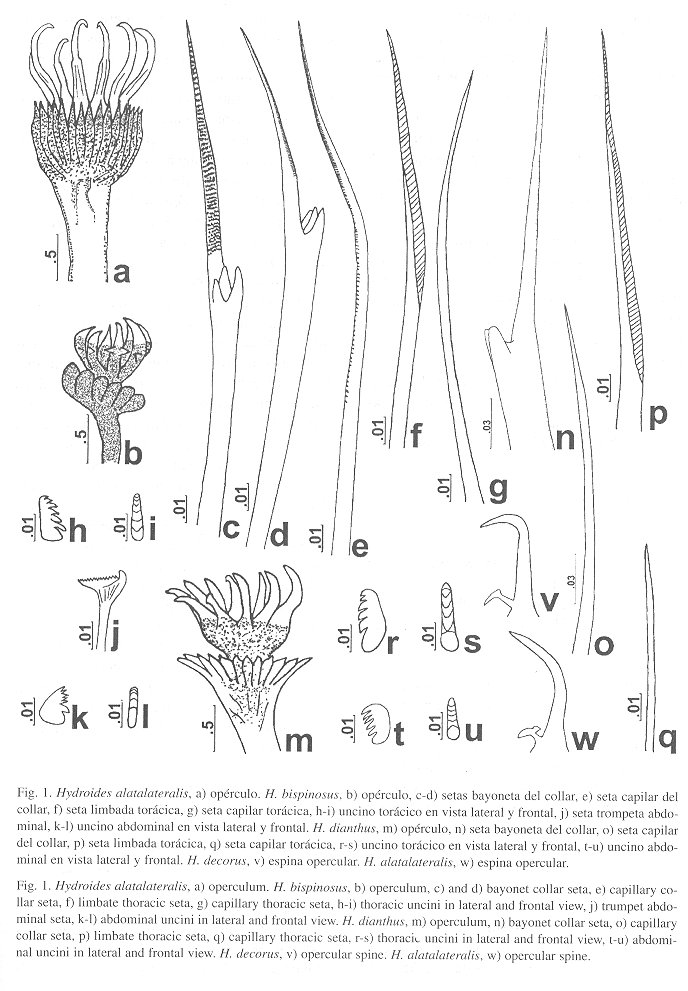
Hydroides alatalateralis(Jones, 1962)
(Fig. 1a, w)
Referencia: Jones 1962: 205-207, Fig. 139-146.
Material examinado: 3 ejemplares. Holotipo (AMNH 3617), 1 paratipo (AMNH 3618), CU6(1).
Caracterización: Tubo blanco con una serie de anillos transversales, pequeños y delgados en toda su longitud. Cuerpo amarillo pálido, con 28 mm de largo. Pedúnculo opercular de 7 mm de longitud. Embudo opercular con radios delgados y aguzados; verticilo con 14 espinas quitinosas, hialinas, ligeramente incurvadas a partir de la porción media distal. A ambos lados de las espinas se presentan alas o limbos delgados, que van desde la base hasta la parte media de la espina (Fig. 1a). Cada espina presenta en su parte basal una espínula interna aguzada, la cual lleva un pequeño tubérculo dorsal. Tórax de 4 mm de largo y 1.5 mm de ancho. Setas del collar bayoneta, con dos dientes en el hombro y limbo liso, así como setas capilares con una fina denticulación medio-distal. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con seis dientes aguzados en una hilera, similares entre sí. Abdomen con setas trompeta, distalmente dentadas y con un pequeño lóbulo en el lado externo. Uncinos similares a los torácicos pero más pequeños, con seis dientes en una hilera, el primero es ligeramente más grueso que los restantes.
Distribución y hábitat: Anfiamericana? Florida, Jamaica, norte de Cuba y el Pacífico de Colombia (?). El ejemplar se encontraba con fango dentro del tubo.
Observaciones: Ejemplares de H. alatalateralis han sido referidos como H. uncinatus, pero el nombre fue reconocido como nomen dubium por Zibrowius (1971) por lo que no se reconoce como especie real. Como otras especies del grupo Eupomatus han sido confundidas con esta especie, Zibrowius (1971) propuso que no se siguiera utilizando dicho nombre; de esta forma, el registro de Fauvel (1953) correspondería a H. alatalateralis (véase Zibrowius 1971: 694). Asimismo, H. floridanus (Bush, 1910), nombre propuesto para el H. uncinatus de Ehlers (1887) puede estar presente en Campeche o el canal de Yucatán (ten Hove, com. pers. 1999, también ver nota en la clave de Bastida-Zavala & Salazar-Vallejo en prep.). Laverde-Castillo (1988: 88) registró la especie para el Pacífico colombiano; sin embargo, no mencionó ni ilustró el característico limbo lateral de las espinas del verticilo. Por ello debe tomarse este registro con precaución hasta confirmar la presencia de este carácter. Con las características que se aprecian en la figura y comentarios, su ejemplar puede incluirse en H. alatalateralis o en H. spongicola, sin embargo esta última no se ha registrado en la costa Pacífico de América.
Características del holotipo: Tiene 24 mm de largo y 2 mm de ancho. Comparando las espinas operculares del paratipo (Fig. 1w) con la de H. decorus, este último es menor [incongruente comparando con el pie de la figura] y tiene el opérculo con espinas mejor desarrollados, pero también presenta la espínula accesoria (Fig. 1v). Esta diferencia pudiera atribuirse al modo de vida en relación con la zona de mareas o con el eventual aporte de agua de baja salinidad que obstruya la consolidación (o favorezca) de cada una de las espinas operculares.
Hydroides bispinosusBush, 1910
(Fig. 1b-l)
Referencia: ten Hove & Wolf 1984: 55.19-21, Fig. 55.14.
Material examinado: 108 ejemplares. CHA1-3(88), CHA9-10(1), CHA13-14(80), CON1(3), CON3(1), COZ1(5), CU1(1), LAG1(3), NC3(1), SF1(1).
Caracterización: Tubo blanco y sinuoso, con costillas longitudinales interrumpidas. Cuerpo amarillo. Embudo opercular con radios anchos y redondeados en su ápice. Verticilo compuesto por siete espinas quitinosas, amarillas, hialinas, e incurvadas suavemente, las dorsales son ligeramente más grandes que las ventrales (Fig. 1b). Las espinas presentan un par de espínulas laterales en su porción media, así como un diente basal interno. Setas del collar bayoneta, con tres dientes en el hombro y limbo piloso (Fig. 1c-d), así como setas capilares con una fina serración medio-distal (Fig. 1e). Los demás setígeros torácicos con setas limbadas (Fig. 1f) y capilares (Fig. 1g). Uncinos torácicos subtriangulares, con siete dientes en una hilera, todos aguzados excepto el primero que es obtuso (Fig. 1h-i). Abdomen con setas trompeta, distalmente dentadas y con el diente externo más ancho y fuertemente incurvado en forma de gancho (Fig. 1j). Uncinos similares a los torácicos pero más pequeños, con seis dientes en una hilera, los dos primeros más gruesos que los restantes (Fig. 1k-l).
Distribución y hábitat: Gran Caribe. Florida, Bermuda, Cuba. Sobre rocas calcáreas, en el talo de algas (Halimeda, Udotea), en roca coralina, en pilotes de madera con los tubos cubiertos por briozoos.
Observaciones: Las espinas del verticilo de algunos ejemplares son similares en tamaño; mientras que en otros casos, las espinas dorsales son mucho más grandes y anchas que las ventrales; o bien, ejemplares con espinas cortas y muy anchas. Algunos ejemplares presentan el verticilo incoloro, o amarillo, pero en otros ejemplares se presentan las espínulas y los bordes de las espinas oscuras, e incluso existen ejemplares con el verticilo totalmente negro. Asimismo, se encontraron algunos ejemplares con dos opérculos, de tamaños diferentes. Por la variabilidad en la forma de las espinas del verticilo de esta especie, es confundida frecuentemente con H. parvus. Sin embargo, los radios del embudo opercular son anchos y romos en H. bispinosus, mientras que en H. parvus son más delgados y aguzados. Además, esta última especie presenta espinas en ángulo recto al contrario de H. bispinosus, cuyo doblez es más suave. Por último, las espínulas laterales de H. parvus son más grandes y agudas que las de H. bispinosus.
Hydroides decorus(Treadwell, 1931)
(Fig. 1v)
Descripción: Treadwell 1931: 4-5, Fig. 3a-d.
Material examinado: Tres paratipos (AMNH 2033), Grand Isle, Lousiana, sin fecha.
Características: Tres ejemplares, uno completo y al resto les faltan algunos segmentos abdominales. El organismo completo mide 15 mm de largo y 2 mm de ancho, con radiolos de 3.5 mm y pedúnculo opercular de 6 mm. El opérculo presenta un embudo opercular y un verticilo con 11 espinas recurvadas simples, con nódulos poco notorios en vista dorsal. Tres espinas dorsales son un poco mayores que las demás.
Observaciones: Para Hartman (1951: 118, 1956: 250) es E. dianthus (Verrill, 1873), pero son diferentes. El frasco tiene una nota colocada por Zibrowius en 1970, donde se le renombra como Hydroides decorus (Treadwell, 1931).
Hydroides dianthus(Verrill, 1873)
(Fig. 1m-u)
Referencia: Zibrowius 1971: 697-705, Fig. 1-5.
Material examinado: 87 ejemplares. CEL1-2(30), CEL4(2), CHA1-3(3), CHA9-10(3), CON1(18), CON3(4), LAG1(2), NC2b(1), NC3(1), SF1(20), SJU1(1), YA12(1).
Caracterización: Tubo blanco y sinuoso, con rugosidades longitudinales que dan la apariencia de crestas. Cuerpo amarillo. Embudo opercular ligeramente asimétrico con radios enteros y aguzados. Verticilo compuesto por 11 espinas quitinosas, hialinas; las dorsales son mayores y están incurvadas, mientras que algunas espinas ventrales son más cortas, delgadas y recurvadas (Fig. 1m). Setas del collar bayoneta, con dos dientes en el hombro y limbo liso (Fig. 1n), así como setas capilares (Fig. 1o). Los demás setígeros torácicos con setas limbadas (Fig. 1p) y capilares muy delgadas (Fig. 1q). Uncinos torácicos subrectangulares, con seis dientes anchos en una hilera, el primero mucho mayor que los demás (Fig. 1r-s). Abdomen con setas trompeta, distalmente dentadas, diente externo fuertemente incurvado. Uncinos similares a los torácicos pero más pequeños; también presentan seis dientes en una hilera, aunque más delgados (Fig. 1t-u).
Distribución y hábitat: Anfiatlántica. Gran Caribe, Golfo de Viscaya (introducida), Mediterráneo (introducida), Senegal. En rocas calcáreas de playas mixtas de arena, talo de algas (Halimeda, Udotea), pilotes de madera con los tubos cubiertos por briozoos, epibionte de ascidias en una pared de vermétidos y ostras. La especie fue introducida al Mediterráneo desde el siglo XIX y también ha llegado a las costas atlánticas de Francia (Zibrowius 1978). Esta dispersión ha sido facilitada por la transportación marítima.
Observaciones: Varían la forma y tamaño de las espinas del verticilo, tanto en el tamaño relativo entre dorsales y ventrales, como su incurvación. Asimismo, algunos opérculos presentan 1-2 espinas dobladas hacia fuera y una o dos espinas rectas, mientras que en otros son 3-6. Algunos ejemplares presentan dos opérculos de tamaño diferente, pero con el mismo arreglo del verticilo. Ten Hove (com. pers. 1999) considera que puede haber confusión entre H. dianthus e H. sanctaecrucis (Mörch, 1863), sin embargo, la primera no tiene espínulas interna o externa, mientras que la segunda tiene un par de espínulas internas (sensu Rioja 1958) o una espínula externa (sensu ten Hove com. pers. 1999). Además, H. dianthus presenta espinas geniculadas y espinas dirigidas hacia fuera, mientras que H. sanctaecrucis presenta sólo espinas geniculadas.
Hydroides diramphusMörch, 1863
(Fig. 2a-d)
Referencia: Zibrowius 1971: 705-707, Fig. 6-9.
Material examinado: 7 ejemplares. SJU1(7).
Caracterización: Tubo blanco y sinuoso, con rugosidades longitudinales que dan la apariencia de crestas. Cuerpo amarillo. Embudo opercular ligeramente asimétrico y oscuro, presenta radios triangulares. Verticilo con 14 espinas quitinosas transparentes, con la punta expandida lateralmente en forma de T y la parte distal redondeada, similares en tamaño y con un pequeño diente basal (Fig. 2a-b). Setas del collar bayoneta, con dos dientes en el hombro y limbo liso (Fig. 2c), así como setas capilares (Fig. 2d). Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con seis dientes anchos en una hilera, el primero mucho mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con el diente externo fuertemente incurvado. Uncinos similares a los torácicos pero más pequeños, con seis dientes en hilera.
Distribución y hábitat: Gran Caribe. Desde Bahamas a Brasil. Epibionte de ascidias en una pared de vermétidos y ostras. Zibrowius (1978) consideró que la especie llegó al Mediterráneo en tiempos recientes, tal vez el siglo XIX.
Observaciones: Los registros de esta especie en el Mediterráneo y el Indopacífico podrían incluir, por lo menos, una especie hermana, H. lunulifer (Claparède, 1870), registrada también en el Caribe (e.g.Liñero-Arana 1999). Sin embargo, Zibrowius (1971), que revisó material del Mediterráneo, del Caribe y del Indopacífico, consideró que H. diramphus es circumtropical y de aguas templadas. Mantuvo en H. diramphuss.l. a varias especies como H. cumingii (Mörch, 1863), H. lunulifer (Claparède, 1870) e H. malleophorus Rioja, 1942. Ten Hove y Zibrowius (com. pers. 1999) revisaron cientos de ejemplares de todo el mundo y no encontraron diferencias para mantener los nombres anteriores independientes de H. diramphus, ni siquiera para mantener este nombre como sensu lato. Sin embargo, tenemos la sospecha que pueda tratarse de un complejo de especies, en vista de que no existen pruebas suficientes que aclaren cómo las poblaciones en cada una de las franjas tropicales marinas puedan mantener el flujo génico, sobre todo entre ambos lados del océano Pacífico.
Hydroides elegans(Haswell, 1883)
(Fig. 2e-f)
Referencia: Zibrowius 1971: 721-727, Fig. 56-64.
Material examinado: 27 ejemplares. CEL1-2(2), CHA1-3(9), CHA9-10(1), CON1(10), SJU1(5).
Caracterización: Tubo blanco y sinuoso, con rugosidades longitudinales que dan la apariencia de crestas. Cuerpo amarillo Embudo opercular con radios anchos y redondeados. Verticilo con 13 espinas quitinosas, hialinas, rectas, de tamaño similar; cada una con 2-3 pares de espínulas laterales así como un diente interno basal (Fig. 2e). Algunos ejemplares presentan dos dientes, uno basal y uno medio; otros presentan un proceso espinoso en el centro del verticilo. Setas del collar bayoneta, con muchos dientes en el hombro que le dan apariencia pilosa y un limbo finamente denticulado (Fig. 2f), así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con seis dientes anchos en una hilera, el primero mucho mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con un diente externo más ancho y obtuso. Uncinos similares a los torácicos pero más pequeños, con seis dientes en una hilera.
Distribución y hábitat: Circumtropical. Gran Caribe, Mediterráneo, Mar del Norte (introducida), sur de Africa, golfo Pérsico, India, Australia e islas del Pacífico. En rocas calcáreas sobre playas de arena, talo de algas (Halimeda, Udotea), pilotes de madera con los tubos cubiertos por briozoos, epibionte de ascidias en una pared de vermétidos y ostras.
Observaciones: Esta especie parece ser de amplia distribución y ha sido confundida en muchas ocasiones con H. norvegicus, debido a que las espinas operculares son parecidas y muy variables. Sin embargo, Zibrowius (1971) clarificó el estatus de ambas especies, cuya diferencia principal se encuentra en la denticulación basal de las setas del collar; H. norvegicus presenta dos grandes dientes en la parte media de la seta, mientras que H. elegans tiene muchos dentículos que le dan un aspecto piloso (Fig. 2f). También se han realizado experimentos de fertilización cruzada entre ambas especies y, a pesar de que el porcentaje de fertilización interespecífica efectiva (división celular observada) fue similar al de la intraespecífica, la liberación de trocóforas obtenidas del entrecruzamiento de las especies fue muy reducida (Gee 1963).
H. elegans fue descrita para Australia pero se ha registrado en todo el Indopacífico y en el Caribe (e.g. Liñero-Arana 1999). Aparentemente la especie llegó a Europa en el siglo XIX (Zibrowius 1978). Su amplia dispersión se debe a los movimientos transoceánicos de las navieras comerciales; su éxito para ocupar nuevas zonas está asociada a la descarga de aguas termales de centrales de energía (ten Hove 1974, Zibrowius 1978), y a su tolerancia a la contaminación industrial (Moran & Grant 1984).
Hydroides gairacensisAugener, 1934
(Fig. 2g)
Referencia: Zibrowius 1969: 366-376, Fig. 2.
Material examinado: 3 ejemplares. CU2(1), IXC1(1), PAC1(1).
Caracterización: Sin tubo. Cuerpo amarillo. Embudo opercular, ligeramente asimétrico, con radios negros, con la punta expandida en forma de T. Verticilo con siete espinas quitinosas, transparentes y con el tercio distal negro. Las espinas son similares en tamaño (Fig. 2g). Setas del collar bayoneta, con dos dientes en el hombro y limbo liso, así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con ocho dientes anchos en una hilera. Abdomen con setas trompeta, distalmente dentadas y con el diente externo incurvado. Uncinos similares a los torácicos pero más pequeños. Presentan cinco dientes en una hilera.
Distribución y hábitat: Anfiamericana. Mar Caribe, golfo de Panamá y sur de Brasil. Sobre coral y en pastos marinos (ceibadal).
Observaciones: Es una especie muy rara pues se han encontrado pocos ejemplares de la misma en pocos sitios, muy distantes entre sí (Zibrowius 1969, Ibarzábal 1986, ten Hove & San Martín 1995).
Hydroides mongeslopeziRioja, 1958
(Fig. 2h-j)
Referencia: Rioja 1958: 257-259, Fig. 4.
Material examinado: 3 ejemplares. CU3(2), PO75003(1).
Caracterización: El fragmento de un tubo es blanco y presenta tres protuberancias dorsales (Fig. 2h). Cuerpo amarillo. Embudo opercular ligeramente asimétrico con radios anchos y distalmente redondeados. Verticilo con 13 espinas quitinosas, separables en dos secciones (Fig. 2i). Parte basal negra y con 3-4 espínulas laterales, una pequeña espínula dorsal en su borde distal y una espínula interna aguzada e incurvada (Fig. 2j). Parte distal de la espina hialina y aguzada, incurvándose ligeramente. Presenta 2-3 pequeños dientes basales arreglados en triángulo y dos dientes laterales, todos negros (Fig. 2j). Setas del collar bayoneta, con dos dientes en el hombro y limbo finamente serrado, así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con ocho dientes aguzados en una hilera, el primero mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con el diente externo fuertemente incurvado. Uncinos similares a los torácicos pero más pequeños, con cinco dientes agudos en una hilera.
Distribución y hábitat: Golfo de México y Cuba. En roca coralina, asociado a arrecifes.
Observaciones: Es una especie muy rara. Después de la descripción de Rioja (1958) para Playa Norte e isla de Santiaguillo, Veracruz, sólo se ha vuelto a encontrar en Cuba y en el arrecife de Triángulos Oeste.
Hydroides mucronatusRioja, 1958
(Fig. 2k, 3a-d)
Referencia: Rioja 1958: 254-257, Fig. 3.
Material examinado: 1 ejemplar. CHA1-3(1).
Caracterización: Sin tubo. Cuerpo amarillo pálido, con 2 mm de largo. Pedúnculo opercular con 1 mm de largo. Embudo opercular con 14 radios. Verticilo hialino con seis espinas quitinosas rectas, aguzadas y dirigidas hacia arriba; cada una con dos dientes laterales diminutos (Fig. 2k). Tórax con 0.7 mm de largo y 0.2 mm de ancho. Setas del collar bayoneta, con dos dientes en el hombro y limbo liso, así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con ocho dientes agudos en una hilera, el primero ligeramente mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con el diente externo más ancho e incurvado. Uncinos similares a los torácicos pero más pequeños, con seis dientes agudos en una hilera.
Distribución y hábitat: Golfo de México (Veracruz). Sobre el talo de algas (Halimeda, Udotea).
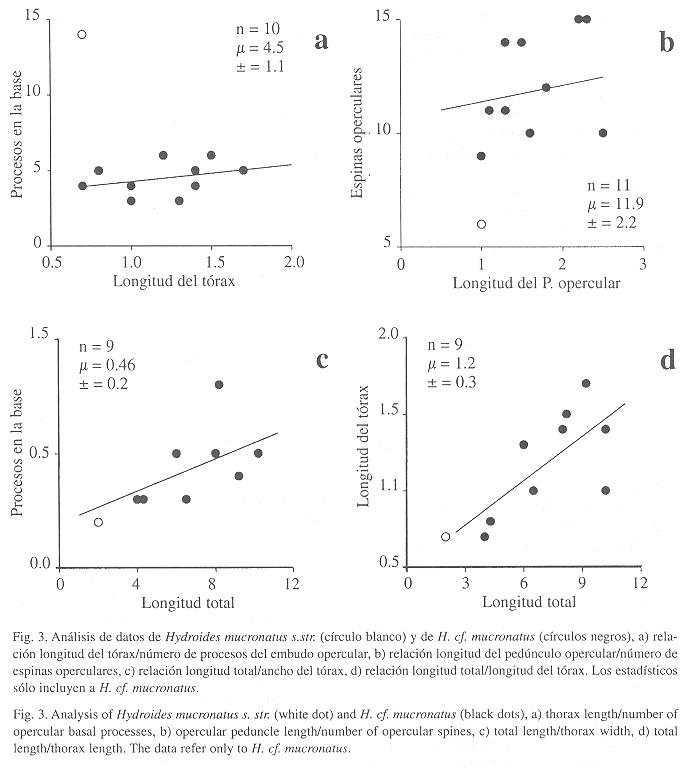
Observaciones: Al parecer, se trata del primer registro de H. mucronatus s.str. después de la descripción de Rioja (1958). Aunque las características del opérculo coinciden, el ejemplar es minúsculo (Fig. 3a-d). Véase observaciones de H. cf. mucronatus.
Hydroides cf. mucronatusRioja, 1958
(Fig. 2l-m, 3a-d)
Referencias: ten Hove 1984: 191, Fig. 6a-c; ten Hove & San Martín 1995: 15.
Material examinado: 11 ejemplares. AVE4(2), BUE1(1), CON1(1), COZ1(1), CU4(2), ECH2(1), ECH3(1), NC3(1), XCA3(1).
Caracterización: Tubo blanco, con rugosidades discontinuas. Cuerpo amarillo pálido, con 10.2 mm de largo (n=9, r: 4-10.2, µ=7.4 ±2.3). Pedúnculo opercular con 1.6 mm de largo (n=11, r: 1-2.5, µ=1.7 ±0.6). Embudo opercular atípico, presenta cinco procesos carnosos amarillos (n=11, r: 3-6, µ=4.4 ±1.1). Verticilo pardo oscuro con diez espinas quitinosas (n=11, r: 9-15, µ=11.9 ±2.2), con dos dientes laterales diminutos (Fig. 2l). Todas las espinas son rectas, aguzadas y dirigidas hacia arriba (Fig. 2m). Tórax con 1.4 mm de largo (n=10, r: 0.7-1.7, µ=1.2 ±0.3) y 0.5 mm de ancho (n=10, r: 0.3-0.8, µ=0.5 ±0.2). Setas del collar bayoneta, con dos dientes en el hombro y limbo liso, así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con ocho dientes agudos en una hilera, el primero ligeramente mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con el diente externo más ancho e incurvado. Uncinos similares a los torácicos pero más pequeños, con seis dientes agudos en una hilera.
Distribución y hábitat: Gran Caribe. Florida, Golfo de México, Antillas, Atlántico de Panamá. Sobre pilotes de madera con los tubos cubiertos por briozoos, adheridos a rocas coralinas y calcáreas.
Observaciones: El verticilo opercular de los ejemplares estudiados es muy semejante al descrito por Rioja (1958); sin embargo, todos presentan una serie de procesos carnosos (3-6) en lugar de un embudo con radios. Es relevante insistir en que nuestros ejemplares son pequeños (hasta 10 mm), mientras que los ejemplares de Rioja presentan una longitud mínima de 10 mm y una máxima de 15 mm. La primera impresión supondría que se trata de ejemplares con un opérculo en desarrollo o en regeneración. Sin embargo, el ejemplar identificado como H. mucronatus s.str. es más pequeño y tiene un embudo opercular con 16 radios. Por otra parte, el verticilo de los demás ejemplares está completamente quitinizado y se ha señalado (e.g. H. dianthus) que el desarrollo opercular inicia con el embudo y continúa después con el verticilo (ten Hove 1984: 186), lo cual contradice la suposición inicial de que sean ejemplares con un opérculo en regeneración.
La serie de transformación del opérculo propuesta por ten Hove (1984), para Serpula, Crucigera e Hydroides, muestra pasos sucesivos en la transformación del opérculo, desde un embudo sencillo (Serpula), pasando por un embudo con procesos basales (Crucigera) hasta llegar a Hydroides, con dos estructuras bien diferenciadas: embudo y verticilo. Así, H. cf. mucronatus parece encajar en este esquema a manera de eslabón entre Crucigera y el Hydroides típico; H. priscus Pillai, 1971, presenta un verticilo muy simple, con un embudo parcialmente desarrollado, aunque se sospecha que esta pueda ser inválida si se comprueba que los dos pequeños ejemplares, usados en la descripción original, eran inmaduros como aseguran Ben-Eliahu & ten Hove (1989). Por su parte, ten Hove (1984) y ten Hove & San Martín (1995), consideran que estos H. cf. mucronatus, con proto-embudo, pueden ser una especie diferente. Ellos revisaron cerca de 500 ejemplares del Caribe y no se encontró ningún ejemplar con el opérculo que dibujó Rioja (1958). Otra evidencia más, que refuerza la idea de una especie indescrita, es que las espinas de nuestros ejemplares presentan una espínula basal redondeada, misma que falta en las espinas dibujadas por Rioja e incluso mencionó dicha carencia (p: 256).
Se realizó un análisis morfológico con los ocho ejemplares y el ejemplar de H. mucronatus s.str. Se aclara que un ejemplar presentó doble pedúnculo opercular y las medidas de ambos opérculos fueron utilizadas, por lo que el tamaño de muestra será igual a nueve en los casos que procedió. Al comparar la longitud del tórax con la longitud del pedúnculo opercular (ver Bastida-Zavala & Salazar-Vallejo en prep), el número de procesos basales (Fig. 3a), y el número de espinas operculares (Fig. 3b), se observa una clara estabilización del número de procesos; es decir, es independiente de la longitud o del número de espinas. En los últimos dos casos el dato disparado corresponde al único ejemplar con embudo estelado. Por otro lado, la longitud total en relación con el ancho del tórax (Fig. 3c) muestran una tendencia isométrica poco definida; mientras que al relacionar la longitud total con el ancho del tórax (Fig. 3d) se observa una tendencia claramente isométrica. Al relacionar la longitud del pedúnculo opercular con el número de espinas operculares, se observa que, por igual, se encuentran pedúnculos cortos con 10-14 espinas operculares como pedúnculos largos con sólo diez espinas. Esto indicaría que, independientemente de una posible regeneración del opérculo, el número de espinas siempre resulta elevado y se estabiliza por arriba de las nueve espinas. Por el contrario, H. mucronatus (sensu Rioja 1958) presenta como máximo nueve espinas (r: 7-9) y el ejemplar de H. mucronatus s.str. que encontramos sólo tiene seis espinas. Los dibujos de ten Hove (1984: 192, Fig. 6a-c) también confirman esta idea.
Por las relaciones morfométricas que se mencionaron resulta evidente que se trata de un grupo homogéneo de organismos, a pesar de que provienen de diversas localidades y por ende, diferentes a H. mucronatus s.str. Como mencionan ten Hove & San Martín (1995), faltaría revisar ejemplares topotípicos (Isla Sacrificios, Veracruz).
Hydroides parvus (Treadwell, 1902)
(Fig. 2n-p)
Referencia: Rioja 1958: 251-254, Fig. 1-2.
Material examinado: 131 ejemplares. Un tipo (USNM 16173), CCE1(2), CEL1-2(1), CHA1-3(18), CHA9-10(1), CHA13-14(+100), CON1(1), ECH4(2), EL2779(1), ESP8(1), ESP10(1), F5(1), PO75001(1).
Caracterización: Tubo blanco y sinuoso, con rugosidades longitudinales discontinuas. Cuerpo amarillo. Embudo opercular ligeramente asimétrico, con radios enteros y aguzados. Verticilo con ocho espinas quitinosas de color amarillo. Cada espina incurvada en ángulo recto, con dos espínulas laterales en el tercio basal y un capítulo dorsal, presentando la apariencia de un buitre en vista lateral (Fig. 2n). Setas del collar bayoneta, con dos dientes en el hombro y limbo liso, así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con seis dientes anchos en una hilera, el primero mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con el diente externo fuertemente incurvado. Uncinos similares a los torácicos pero más pequeños, con seis dientes más delgados.
Distribución y hábitat: Gran Caribe. Carolina del Norte hasta Brasil. Sobre raíces de mangle, en rocas calcáreas sobre playas de arena, sobre el talo de algas (Halimeda, Udotea), en pilotes de madera con los tubos cubiertos por briozoos, como epibionte del erizo Eucidaris tribuloides, en roca coralina.
Observaciones: Esta especie se ha confundido muchas veces con H. bispinosus por tener opérculos similares (ver comentarios de esa especie).
Características del ejemplar tipo: Ejemplar destrozado. Corona radiolar y opérculo desprendidos del cuerpo. Cuerpo de 6 mm de largo y 0.7 mm de ancho en su porción anterior; tiene una incisión a lo largo del cuerpo. Radiolos de 3 mm de largo, verticilo opercular de 1 mm de largo. Opérculo asimétrico, con ocho espinas con un desarrollo variable de la espínula interna distal, siendo pequeño y triangular en las espinas cortas, delgada y recurvada en espinas largas. El capítulo de las espinas tiende a desaparecer en las espinas mayores. Todas las espinas tienen un par de pequeñas espínulas triangulares. Dientes basales verticilados.
Hydroides protulicola Benedict, 1887
Referencia: Benedict 1887: 550, Lám. 20, Fig. 17, Lám. 21, Fig. 18-23.
Material examinado: Sintipos (USNM 972), frente a Cabo Hatteras, NC, Albatross, estación 2307, 43 brazas, 1884.
Características: 25 sintipos y porciones anteriores sin opérculo (separados), los sintipos tienen opérculo pero no todos están completos. Opérculo simétrico, los radios del embudo cubren la base de las espinas centrales. Todas son amarillentas y transparentes, con la punta dirigida hacia fuera y sin espínulas accesorias.
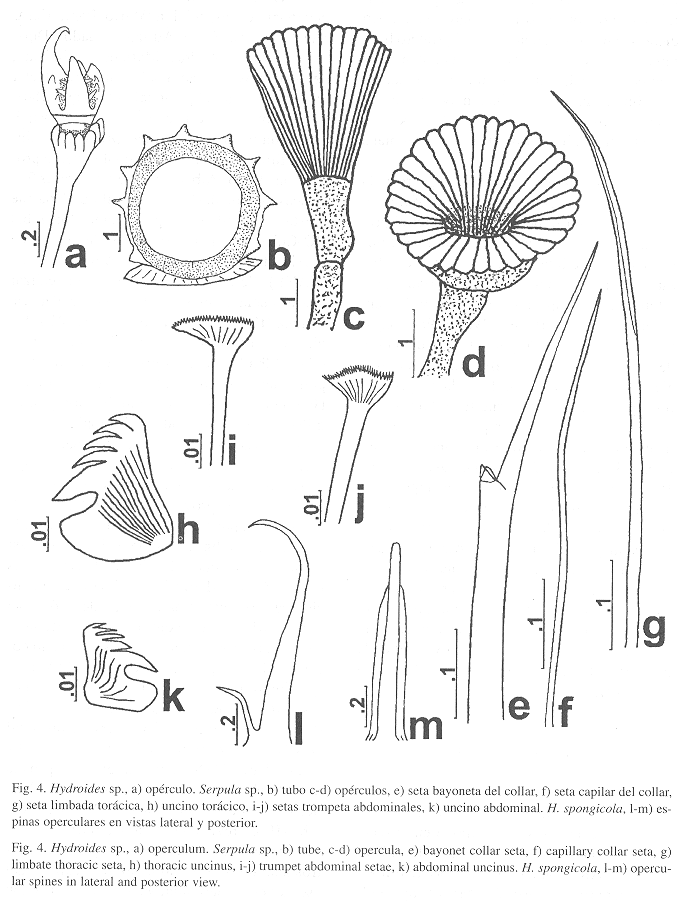
Hydroides spongicolaBenedict, 1887
(Fig. 4l-m)
Referencia: Benedict 1887: 549-550, Lám. 20, Fig. 11-12, Lám. 21, Fig. 13-16.
Material examinado: Sintipos (USNM 975), Tampa Bay y Dry Tortugas, Florida, Albatross, estación 2409, 26 brazas, 18 III 1885 (laminillas 147-153 no vistas).
Características: Varios ejemplares incompletos de 1-2 cm de largo. Opérculo simétrico con verticilo basal de espinas muy largas, amarillentas y aguzadas, que llegan hasta la mitad de las espinas centrales. Espinas del verticilo distal o centrales muy delgadas, incurvadas, con una espínula interna muy desarrollada (Fig. 4l); las espinas también presentan limbos laterales que alcanzan tres cuartas partes de su longitud (Fig. 4m).
Hydroides sp.
(Fig. 4a)
Material examinado: 1 ejemplar. CU5(1).
Caracterización: Sin tubo. Cuerpo incompleto, amarillo. Pedúnculo opercular con 2.5 mm de largo. Embudo opercular asimétrico, con radios redondeados. Verticilo con cinco espinas quitinosas, con forma de botella, pardo rojizas, uno de ellas mucho más grande que las demás (Fig. 4a). Las espinas son rectas pero en su parte distal están ligeramente incurvadas, excepto la espina mayor que está muy recurvada a modo de gancho. Tienen cuatro espínulas laterales basales, asimétricas, así como una espínula interna basal. Tórax con 1 mm de largo y 0.4 mm de ancho. Setas del collar bayoneta, con dos dientes en el hombro y limbo liso, así como setas capilares. Los demás setígeros torácicos con setas limbadas y capilares. Uncinos torácicos subrectangulares, con seis dientes anchos en una hilera, el primero mayor que los demás. Abdomen con setas trompeta, distalmente dentadas y con el diente externo fuertemente incurvado. Uncinos similares a los torácicos pero más pequeños, con seis dientes más delgados.
Distribución y hábitat: Cuba. En el área de los cayos, a 15 m de profundidad.
Observaciones: Esta especie es similar a H. rostratus Pillai, 1971, que fue descrita de Sri Lanka (océano Indico); sin embargo, las espinas no están tan incurvadas ni presentan una espínula externa como la especie de Pillai, sino que son dos pares de espínulas laterales. Otra especie similar es H. exaltatus Marenzeller, descrita para Japón. Se diferencía en que los radios del embudo opercular son aguzados en H. exaltatus mientras que en la forma de Cuba son redondeados. Además, las espinas de H. exaltatus, excepto la más grande, están dirigidas hacia fuera mientras que el ejemplar de Cuba las tiene dirigidas ligeramente hacia adentro. Asimismo, ten Hove & Wolf (1984) registra a Hydroides sp. A para el norte del Golfo de México, la cual es diferente a la nuestra porque tiene 6-7 espinas, todas incurvadas, con dos pares de espínulas laterales. Por su parte, Liñero-Arana (1999) registra a Hydroides sp. para Venezuela, el cual presenta 7-8 espinas fuertemente incurvadas y sin espínulas laterales, por lo cual tampoco se trata de la misma especie.
Serpula Linnaeus, 1758
Serpula sp.
(Fig. 4b-k)
Material examinado: 2 ejemplares. CU5(1), EL2792(1).
Caracterización: Tubo pardo con nueve costillas longitudinales con crestas suaves (Fig. 4b). Cuerpo amarillo. Opérculo blanco, formado por un embudo blando, con 33 radios, sin tubérculos internos y 3 mm de largo (Fig. 4c-d); el ejemplar de Cuba tiene un embudo con 26 radios, sin tubérculos internos y una longitud de 2.2 mm. Membrana torácica fusionada ventralmente formando un pequeño aprón (mandil). Setas del collar bayoneta, con dos dientes en el hombro (Fig. 4e), así como setas capilares (Fig. 4f). Los demás setígeros torácicos con setas limbadas (Fig. 4g) y capilares. Uncinos torácicos triangulares, con cinco dientes aguzados en una hilera, el primero ligeramente mayor (Fig. 4h). Abdomen con setas trompeta, distalmente con una denticulación muy fina (Fig. 4i-j). Uncinos muy similares a los torácicos pero más pequeños (Fig. 4k).
Distribución y hábitat: Cuba y Quintana Roo.
Observaciones: Day (1973) registró en la región a S. vermicularis granulosa Marenzeller, 1884, descrita de Japón; su material presenta 20-40 radios operculares con tubérculos en la superficie interna del embudo opercular, estas últimas son un carácter importante para diferenciar las especies (ten Hove & Jansen-Jacobs 1984). Ten Hove & Wolf (1984) registraron a Serpula sp. A, para el norte del Golfo de México, la cual es diferente de nuestros ejemplares porque tiene 18 radios operculares, aparentemente no tiene tubérculos internos y las setas del collar presentan una fina denticulación proximal. Kupriyanova (1999) revisó el estado de S. cf. columbiana Johnson, 1901 y mostró que la supuesta cosmopolita, S. vermicularis incluye tres especies: S. vermicularis, para el Atlántico norte y Artico, S. columbiana para el Pacífico de América y describió S. uschakovi para las formas del Pacífico boreal occidental. Para separar las especies utiliza el color del tubo, peristomas, promedio de radios operculares, constricción entre el pedúnculo y el opérculo y fusión ventral de la membrana torácica. Nuestros ejemplares son una especie indescrita.
Agradecimientos
La recolecta fue autorizada por SEMARNAP (permiso DOO750-1578). Leopoldo Cosgalla (Quelonios A.C.) apoyó en la recolecta en Champotón y junto con Wilbert Borges (ITMAR-Champotón) nos brindaron espacio para trabajar. Adrián Marfil apoyó en la recolecta en Ría Lagartos. Se agradece la hospitalidad y apoyo para la realización de las recolectas en Isla Contoy al personal del Parque Nacional, en especial a Mario Lara, César Sánchez, Omar Ortiz, Adriana Amador, Dorina Basurto y Andrés Mariano. Asimismo, agradecemos las facilidades para recolectar en Isla Cozumel al personal del Parque Marino Nacional, en especial a Elvira Carvajal, Juan Domínguez y Fernando Orozco; el vehículo utilizado en la isla fue amablemente proporcionado por Alberto Sleme (Empresas Turísticas Nacionales). El material de Cuba fue prestado gentilmente por Diana Ibarzábal (IO-Cuba) y otros ejemplares del golfo de México fueron prestados por Angel de León (UANL). Se agradece el apoyo para realizar una estancia y revisar colecciones a Patricia Gómez (ICML), Francisco Solís (ICML), Vivianne Solís (ICML) y Juan Manuel Vargas (UV). Elba Escobar y Luis Soto (ICML) proporcionaron los materiales del "Edwin Link". Humberto Bahena imprimió las fotografías utilizadas en este estudio. El trabajo de campo y las visitas a las colecciones nacionales fueron financiados por el Programa de Apoyo a Tesis de Maestría de ECOSUR, recursos fiscales y por el proyecto CONACYT 4120P-N9607. El apoyo de Scott Monks y Griselda Pulido para la digitalización de las figuras, así como la asistencia en la edición de las mismas de Luis Carrera-Parra fue vital para la conclusión del presente trabajo. Angel de León hizo comentarios que mejoraron este trabajo. JRBZ agradece el tiempo que por derecho correspondía a su pareja, Socorro García-Madrigal e hijas Karla y Marcela. El segundo autor visitó dos museos y agradece el apoyo recibido a Judith Winston y Jerry Thurmond (AMNH, Nueva York) y a Kristian Fauchald y Linda Ward (USNM, Washington). La visita al AMNH fue financiada en parte por la Lerner-Gray Fund for Marine Research del AMNH. La revisión crítica por Harry ten Hove (Universidad de Amsterdam) y de dos árbitros anónimos permitió mejorar la calidad del escrito; el primero también envió información valiosa y observaciones.
Resumen
A partir de la revisión de más de 450 ejemplares de Hydroides y Serpula, 12 especies de la región del Gran Caribe fueron identificadas y caracterizadas. Ocho especies fueron recolectadas en las costas de la Península de Yucatán y cinco fueron encontradas en otras localidades en el Golfo de México, otras siete son de Cuba y se incluyeron comentarios sobre el material tipo de cinco especies más. Se realizó un análisis morfométrico sobre Hydroides mucronatus Rioja e Hydroides cf. mucronatus indicando varias diferencias entre ellas. Se incluyen comentarios a todas las especies
Referencias
Bastida-Zavala, J.R. & S.I. Salazar-Vallejo. En prep. Serpúlidos (Polychaeta: Serpulidae) del Caribe noroccidental con claves para la región del Gran Caribe: Salmacina, Ficopomatus, Pomatoceros, Pomatostegus, Protula, Pseudovermilia, Spirobranchus y Vermiliopsis. Rev. Biol. Trop. [ Links ]
Benedict, J.E. 1887. Descriptions of ten species and one new genus of Annelids from the dredgings of the U.S. Fish Commission steamer "Albatross". Proc. U.S. Natl. Mus. 9: 547-553. [ Links ]
Ben-Eliahu, M.N. & ten Hove, H.A. 1989. Redescription of Rhodopsis pusilla Bush, a little known but widely distributed species of Serpulidae (Polychaeta). Zool. Scripta 18: 381-395. [ Links ]
Day, J.H. 1973. New Polychaeta from Beaufort, with a key to all species recorded from North Carolina. NOAA Tech. Rep. NMFS 375: 1-148. [ Links ]
de León-González, J.A. 1990. Dos serpúlidos nuevos para el Pacífico mexicano y duplicidad opercular en Hydroides crucigerus (Polychaeta: Serpulidae). Rev. Biol. Trop. 38: 335-338. [ Links ]
Ehlers, E. 1887. Report on the annelids, p: 1-355. In Reports on the results of dredging, under the direction of L. F. Pourtalès, during the years 1868-1870, and of Alexander Agassiz, in the Gulf of Mexico (1877-78), and in the Caribbean Sea (1878-79) in the U.S. Coast Survey Steamer "Blake,". Mem. Mus. Comp. Zoöl. Harv. Coll., Cambridge, 15. [ Links ]
Fauvel, P. 1953. Annélides Polychètes de la croisière du "President Théodore Tissier" aux Antilles (1951). Bull. Inst. Oceanogr. Monaco 1033: 1-23. [ Links ]
Gee, J.M. 1963. On the taxonomy and distribution in South Wales of Filograna, Hydroides and Mercierella (Polychaeta: Serpulidae). Ann. Mag. Nat. Hist., ser. 13, 6: 705-715. [ Links ]
Hartman, O. 1951. The littoral marine annelids of the Gulf of Mexico. Publ. Inst. Mar. Sci., Univ. Tex. 2: 7-124. [ Links ]
Hartman, O. 1956. Polychaetous annelids erected by Treadwell, 1891 to 1948, together with a brief chronology. Bull. Amer. Mus. Nat. Hist. 109: 243-310. [ Links ]
Hartman, O. 1959. Catalogue of the polychaetous annelids of the World. Allan Hancock Found. Publ. Occ. Pap. 23: 1-628. [ Links ]
Ibarzábal, D.R. 1986. Lista de especies de poliquetos bentónicos cubanos. Rep. Invest. Inst. Oceanol. Cuba 45: 1-17. [ Links ]
Ichikawa, A. & N. Takagaki. 1942. The reversible asymmetry in the opercula of Hydroides ezoensis, 1. Observations on the intact opercula. J. Fac. Sci. Hokkaido Imp. Univ., ser. 6, Zool. 8: 1-8. [ Links ]
Jones, M.L. 1962. On some polychaetous annelids from Jamaica, the West Indies. Bull. Am. Mus. Nat. Hist. 124: 169-212. [ Links ]
Kupriyanova, E.K. 1999. The taxonomic status of Serpula cf. columbiana Johnson, 1901 from the American and Asian coasts of the North Pacific Ocean. Ophelia 50: 21-34. [ Links ]
Laverde-Castillo, J.J.A. 1988. Notas sobre algunos serpúlidos (Annelida, Polychaeta) de Bahía Málaga, Pacífico Colombiano. An. Inst. Invest. Mar. Punta Betín 18: 83-93. [ Links ]
Liñero-Arana, I. 1999. Serpulidae (Annelida: Polychaeta) de la costa nororiental de Venezuela. Bol. Inst. Oceanogr. Venezuela 38: 00-00. [ Links ]
Moran, P.J. & T.R. Grant. 1984. The effect of industrial pollution on the growth rate of the serpulid polychaete Hydroides elegans (Haswell). Proc. 1st Polychaete Conf., Sydney, p: 361-369. [ Links ]
Perkins, T.H. 1998. Checklist of shallow-water marine polychaetous Annelida of Florida, p. 79-122. In Camp, D.K., W.G. Lyons & T.H. Perkins (eds.). Checklist of selected shallow-water marine invertebrates of Florida. Florida Mar. Res. Inst. Tech. Rep. 3, St. Petersburg. [ Links ]
Perkins, T.H. & T. Savage. 1979. A bibliography and checklist of polychaetous annelids of Florida, the Gulf of Mexico, and the Caribbean Region. Florida Mar. Res. Publ. 14: 1-62. [ Links ]
Pillai, T.G. 1972. A review and revision of the systematics of the genera Hydroides and Eupomatus together with an account of their phylogeny and zoogeography. Ceylon J. Sci. (Biol. Sci.) 10: 7-31. [ Links ]
Rioja, E. 1919. Una curiosa anomalía del Hydroides norvegica Gunn. y algunas consideraciones acerca de la filogenia de los serpúlidos. Bol. R. Soc. Esp. Hist. Nat. 19: 445-449. [ Links ]
Rioja, E. 1941. Estudios anelidológicos, 3. Datos para el conocimiento de la fauna de poliquetos de las costas mexicanas del Pacífico. An. Inst. Biol. UNAM 12: 669-746. [ Links ]
Rioja, E. 1942. Estudios anelidológicos, 4. Observaciones sobre especies de serpúlidos de las costas del Pacífico de México. An. Inst. Biol. UNAM 13: 125-162. [ Links ]
Rioja, E. 1944. Estudios anelidológicos, 12. Observaciones acerca del opérculo de Hydroides crucigera Mörch y descripción de un caso de duplicidad de este órgano. An. Inst. Biol. UNAM 15: 409-414. [ Links ]
Rioja, E. 1958. Estudios anelidológicos, 21. Observaciones acerca de algunas especies de Serpúlidos de los géneros Hydroides y Eupomatus de las costas mexicanas del Golfo de México. An. Inst. Biol. UNAM 28: 247-266. [ Links ]
Risso, A. 1826. Histoire naturelle de principales productions de lEurope meridionale et particulièrement de celles des environs de Nice et des Alpes Maritimes. Paris, F.G. Levrault Libr., 4: 1-439. [ Links ]
Salazar-Vallejo, S.I. 1989. Enrique Rioja y su contribución al estudio de los poliquetos (Annelida: Polychaeta) en México. Brenesia 30: 39-65. [ Links ]
Salazar-Vallejo, S.I. 1996. Lista de especies y bibliografía de los poliquetos (Polychaeta) del Gran Caribe. An. Inst. Biol., ser. Zool. UNAM 67: 11-50. [ Links ]
ten Hove, H.A. 1974. Notes on Hydroides elegans (Haswell, 1883) and Mercierella enigmatica Fauvel, 1923, alien serpulid polychaetes introduced into the Netherlands. Bull. Zool. Mus. Amsterdam 4: 45-51. [ Links ]
ten Hove, H.A. 1984. Towards a phylogeny in serpulids (Annelida: Polychaeta), p. 181-196. In Proc. 1st Polychaete Conf., Sydney. [ Links ]
ten Hove, H.A. 1990. Description of Hydroides bulbosus sp. nov. (Polychaeta, Serpulidae), from the Iranian Gulf, with a terminology for opercula of Hydroides. Beaufortia 41: 115-120. [ Links ]
ten Hove, H.A. & M.J. Jansen-Jacobs. 1984. A revision of the genus Crucigera (Polychaeta; Serpulidae); a proposed methodical approach of serpulids, with special reference to variation in Serpula and Hydroides, p. 143-180. In Proc. 1st Polychaete Conf., Sydney. [ Links ]
ten Hove, H.A. & G. San Martín. 1995. Serpulidae (Polychaeta) procedentes de la I Expedición Cubano-Española a la Isla de la Juventud y Archipiélago de los Canarreos (Cuba). Stud. Nat. Hist. Carib. Region 72: 13-24. [ Links ]
ten Hove, H.A. & P.S. Wolf. 1984. Family Serpulidae Johnston, 1865, p. 55.1-55.34. In J.M. Uebelacker & P.G. Johnson (eds). Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico, Barry A. Vittor, Mobile, 7 vols. [ Links ]
Treadwell, A.L. 1931. Three new species of polychaetous annelids in the collections of the United States National Museum. Proc. U.S. Natl. Mus. 80: 1-5. [ Links ]
Zibrowius, H.W. 1969. Hydroides gairacensis Augener, 1934, a little known serpulid polychaete from Central and South America. Bull. Mar. Sci. 19: 366-376. [ Links ]
Zibrowius, H. 1971. Les espèces Méditerranéennes du genre Hydroides (Polychaeta Serpulidae): Remarques sur le prétendu polymorphisme de Hydroides uncinata. Tethys 2: 691-746. [ Links ]
Zibrowius, H. 1978(1976). Introduction du polychète serpulidae japonais Hydroides ezoensis sur la côte atlantique française et remarques sur la propagation d'autres espèces de Serpulidae. Tethys 8: 141-150. [ Links ]
Datos de los materiales recolectados. En corchetes, los sitios indicados en la Fig. 1 y 2 de un trabajo previo (Bastida-Zavala & Salazar-Vallejo en prep).
AMNH 2033: Grand Isle, Lousiana, sin fecha, W.W. Anderson. [49]
AMNH 3617-18: Puerto Royal, Jamaica, V 1960. [55]
AVE4: Playa Aventuras, DIF, fondo arenoso y coralino, los corales (Acropora spp.) estaban cubiertos de sedimento y algas filamentosas, 1.5 m, 28 II 1999, SISV y JRBZ. [13]
CCE1: Cayo Cedros, Bahía Ascensión, raíz de mangle, 6 III 1987. [15]
CEL1-2: Playa expuesta frente a Celestún, ca. puerto de abrigo, manchón de rocas calcáreas, 0.6 m, 17 II 1999, SISV y JRBZ. [39]
CEL4: Playa expuesta frente a Celestún, ca. puerto de abrigo, manchón de rocas calcáreas, 0.6 m, 17 II 1999, SISV y JRBZ. [39]
CHA1-3: Playa a 20 km al SW de Champotón, arribazones de pastos (Syringodium) y algas (Udotea flabellum, Penicillus dumetosus y Halimeda incrassata), 14 II 1999, SISV y JRBZ. [36]
CHA9-10: Playa a 20 km de Champotón, en algas (Gracilaria sp.) y piedras sueltas, 3.5 m, 15 II 1999, SISV y JRBZ. [36]
CHA13-14: La Manteca, 10 km frente a Champotón, fondo arenoso con pastos (Syringodium, Thalassia) y parches rocosos, 6 m, 16 II 1999, SISV y JRBZ. [36]
CON1: Isla Contoy, raspado en los pilotes del muelle de madera, incrustantes, 0-3 m, 21 II 1999, SISV y JRBZ. [5]
CON3: Isla Contoy, playa Camping, fondo mixto, con grandes moluscos (Strombus sp.) y esponjas, 0.5-1 m, 22 II 1999, SISV y JRBZ. [5]
COZ1: El Cedral, Cozumel, fondo mixto, con rocas, algas y pastos, 1.5-2 m, 24 II 1999, SISV y JRBZ. [29]
CU1: Instituto de Oceanología, costa N de la Habana, rada IDO, sobre madero, 2 m, II 1978 (IOC). [42]
CU2: NW de punta Los Barcos, Golfo de Batabanó, Cuba, E-13, seibadal, 5 m, 29 VIII 1981 (IOC). [43]
CU3: Laguna arrecifal del Cayo Juan García, golfo de Batabanó, Cuba, P1, 10 m, 2 III 1988 (IOC). [44]
CU4: Cayo Juan García, Golfo de Batabanó, Cuba, 5 m, III 1988 (IOC). [44]
CU5: Cayo Diego Pérez, Golfo de Batabanó, Cuba, 15 m, 20 VII 1988 (IOC). [44]
CU6: Bahía Vita, Provincia de Holguín, NE de Cuba, est. 5, 21 IV 1988 (IOC). [47]
ECH2: La Ceiba, Puerto Morelos, epifauna de las espinas de E. tribuloides, 2.9 m, 26 X 1995, F. Laguarda, F. Solís, M. Torres. [10]
ECH3: Frente al ojo de agua, Puerto Morelos, epifauna de las espinas de E. tribuloides, 2 m, 17 II 1995, F. Laguarda, F. Solís, M. Torres, R. Ramírez. [10]
ECH4: Frente al astillero Rodman, fuera del arrecife de Puerto Morelos, col. 51, epifauna de las espinas de E. tribuloides, 19 m, 28 VI 1995, F. Solís, R. Ramírez, M. Torres. [10]
EL2779: S de Banco Chinchorro, sobre concha de ermitaño diogénido, 4 m, 22 VIII 1990, E. Escobar. [21]
EL2792: E de Isla Mujeres, ca. 13 km de la isla (21º14N, 86º36W), 130 m, 28 VIII 1990. [6]
ESP8: Canal de Yucatán, est. 4-5, (22º02N, 87º02W), en esponja (Aplysina lacunosa), 41 m, IV 1985. [57]
ESP10: Puerto Morelos, KII-35B, en piedra coralina, 20 m. [10]
IXC1: Ixchen, Cozumel, 4 VI 1995, SISV y L.F. Carrera-Parra. [25]
LAG1: Trinchera de Nictilic, 1.5-2 km frente a la bocana de Ría Lagartos, fondo mixto con pastos (Syringodium, Thalassia), algas (Caulerpa prolifera.), esponjas y algunos corales aislados (Solenastrea bournoni), 2-4 m, 18 II 1999, SISV y JRBZ. [2]
NC3: Laguna Nichupté, canal de entrada norte, 200 m de SEMARNAP (Cancún), fondo rocoso con algas (Acetabularia, Halimeda opuntia y Laurencia sp.) y esponjas, 0.1-2 m, 20 II 1999, SISV y JRBZ. [8]
PAC1: Paraíso, Isla Cozumel, 5 VI 1995, SISV y L.F. Carrera-Parra. [25]
PO75001: Laguna de Términos, La Bayoneta (ca. de la boca de Puerto Real), est. 1, en Thalassia testudinum, 0.8 m, 9 VI 1982, B. Reveles y A.L. Ibáñez (ICML-UNAM). [35]
PO75003: Triángulos Oeste, B/O Justo Sierra, DIN II, manual, 1 m, X 1990 (ICML-UNAM). [40]
SF1: San Felipe, cerca de una bocana artificial de Ría Lagartos, fondo mixto, con algunos corales pastos (Syringodium) y algas (Caulerpa prolifera y C. pinnata), 2 m, 19 II 1999, SISV y JRBZ. [1]
SJU1: San Juan de Ulúa, UV55, pared con incrustantes (vermétidos, ostras y ascidias), 1-1.5 m, 11 VIII 1996, M.A. Muro (UV). [32]
USNM 972: Frente a Cabo Hatteras, Carolina del Norte, Albatross est. 2307 (35º42N, 74º5430"W), 80 m, 1884. [50]
USNM 975: Tampa Bay-Dry Tortugas, Florida, Albatross est. 2409 (27º04N, 83º2115"W), 50 m, 18 III 1885. [51]
USNM 16173: Bahía Boquerón, Puerto Rico, Str. Fish Hawk, 1898-1899. [53]
XCA3: Xcacel, QR8, rocas, 3 IV 1992, SISV. [12]
YA12: Laguna Yalahau, Punta Chuchú, YH12, 8 IV 1993, JJUR. [3]

1 El Colegio de la Frontera Sur, Unidad Chetumal. Dep. de Ecología Acuática. Apdo. Postal 424, Chetumal, C.P. 77000, QROO, México, Fax (983)204-47, rolando@ecosur-qroo.mx

