











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.48 no.4 San José dic. 2000
Titulo corto: Densidad de cultivo en Lyropecten (Nodipecten) nodosus
Vanessa Acosta Luis Freites y César Lodeiros1
Recibido 28-VI-1999. Corregido 13-VI-2000. Aceptado 10-VI- 2000.
Abstract
The effect of culture density on growth and survival of juveniles was studied in the scallop Lyropecten nodosus with intermediate hanging cultures in Golfo de Cariaco, Sucre state, Venezuela, from December 1993 to July 1994. Six spat-densities (7, 15, 30, 60, 125 and 250 scallops/nets) were evaluated. The initial shell height and dry mass tissue were 9.4 ± 0.18 mm and 0.007 ± 0.0026 g, respectively. The specimens were placed into pearl nets and suspended in a long line at 8 m depth. Growth patterns of somatic tissue were similar, but with different magnitudes and inverse proportionality to culture density. No effect of density was observed on survival. These results suggest that intraspecific competition of cultivated scallops cause stress, which affects growth but not survival. We suggest as strategy for the management of culture density, successive density reductions, to obtain in a six months culture (intermediate culture) specimens of 50 mm in shell height in order to start the final culture period.
Key words
scallop, culture density, hanging culture, Lyropecten (Nodipecten) nodosus, Golfo de Cariaco, Venezuela
La producción total de pectínidos a nivel mundial ha tenido un aumento de su proporción de un 235% desde 1987 a 1996, de igual manera el aporte generado por su cultivo a la producción total también aumentó notablemente, de 26.7 a 73.4% (Anónimo 1998a,b). Este incremento, indudablemente ha sido producto de la tecnificación y adaptación del cultivo de nuevas especies en diversas latitudes, establecidas a causa de la gran demanda y comercialización de pectínidos.
Uno de los factores esenciales para determinar la rentabilidad del cultivo, en función de obtener un crecimiento que conduzca a una producción adecuada, es el de establecer la densidad de organismos mantenidos en recintos de cultivo. Lógicamente, existe una relación inversa entre la densidad de organismos cultivados y el crecimiento. En algunos pectínidos bajo condiciones de cultivo suspendido se ha demostrado esta relación, tales como Argopecten irradians irradians (Rhodes y Widman 1991), Placopecten magellanicus (Côté et al. 1993) y Euvola ziczac (Freites et al. 1995). Estos trabajos muestran que la densidad de organismos para establecer una producción adecuada, difiere entre las especies, siendo lógico pensar que los organismos que en su hábitat natural están dispersos (comportamiento no agregado), como en el caso de Lyropecten nodosus, no pudieran soportar una densidad adecuada para la producción rentable.
L. nodosus, es la especie de mayor tamaño entre los representantes de la familia pectinidae en el Caribe, alcanzando tallas normalmente comprendidas entre 120 y 150 mm, se encuentra frecuentemente en fondos arenosos con substratos duros, entre 5 y 15 m de profundidad. Se distribuye desde las costas de Florida, Estados Unidos, hasta Santa Catarina, Brasil (Lodeiros et al. 1999). En Venezuela, debido a que se distribuye en bajas densidades no sostiene una pesquería organizada; no obstante, es considerada como una especie con un alto potencial para la acuicultura (Vélez y Lodeiros 1990). L. nodosus ha sido estudiada para establecer su ciclo reproductivo en las poblaciones naturales (Vélez et al. 1987), también se ha estudiado la inducción al desove bajo condiciones controladas (Marín 1984); éstos conjuntamente con otras investigaciones sobre el mantenimiento larvario y postlarvario (Vélez y Freites 1993), han permitido establecer la factibilidad de producción de semillas. Otros estudios se han orientado hacia la mejora del cultivo estableciéndose la variabilidad genética de sus poblaciones (Coronado 1987), el crecimiento en cultivo intermedio (Lodeiros et al. 1998) y la posterior factibilidad de cultivo de fondo y suspensión en la etapa de engorde final (Mendoza 1999)
La presente investigación evalúa el efecto de la densidad de cultivo intermedio de L. nodosus sobre su crecimiento y supervivencia en condiciones suspendidas en el Golfo de Cariaco, Venezuela.
Materiales y métodos
Este estudio se llevó a cabo en la localidad de Turpialito, ubicada en el Golfo de Cariaco, Estado Sucre, Venezuela (10°2656" N; 64°0200" W), en el período de diciembre 1993 - julio 1994. El estudio se realizó con juveniles de L. nodosus (4600 ejemplares) de 9.4 ± 0.18 mm de longitud antero-posterior de la concha, obtenidas en condiciones ambientales controladas, siguiendo la metodología establecida por Vélez y Freites (1993). Los juveniles fueron mantenidos en cultivo suspendido a 8 m de profundidad, utilizando para ello un "long line" de 100 m de longitud y cestas tipo "pearl nets" (1225 cm2 de área de base). Se establecieron seis tratamientos, cada uno de los cuales estuvo conformado por densidades de 250, 125, 60, 30, 15 y 7 ejemplares/cesta, correspondientes a 2040, 1020, 490, 245, 122 y 54 ejemplares/m2, respectivamente. Cada tratamiento estuvo conformado por tres repeticiones experimentales y repeticiones de sustitución, en función de utilizar los ejemplares de éstas últimas para mantener la densidad a través del período experimental.
La supervivencia se estimó mediante el recuento mensual del número de ejemplares vivos por repetición experimental de cada tratamiento. El crecimiento fue determinado por la evaluación mensual de 10 ejemplares por repetición, a los cuales se les determinó la longitud antero-posterior de la concha utilizando un calibrador vernier digital de 0.01 mm de apreciación, así como por la biomasa seca (70 °C/72h) del músculo, la gónada y el resto de tejido del somático.
A la misma profundidad de cultivo, la disponibilidad de alimento fue estimada por el seston en sus componentes total y orgánico determinado mediante técnicas gravimétricas y la biomasa fitoplanctónica por la concentración de clorofila a mediante técnicas colorimétricas, según las recomendaciones en Strickland y Parsons (1972). La temperatura fue registrada a intervalos de 3 minutos mediante el uso de un termógrafo electrónico (SEALOG Vemco LTD, Canadá).
Los parámetros de crecimiento (talla de la concha y masas de la gónada, músculo y resto de tejido somático) fueron analizados cada mes, por una análisis de varianza de una vía (ANOVA I), considerando las densidades de ejemplares por cesta como tratamientos. En aquellos análisis donde se encontraron diferencias significativas (P²0.05), se les aplicó un análisis de comparaciones múltiples de Duncan para diferenciar significativamente (P=0.05) las medias alcanzadas en las diferentes densidades. Para la evaluación de la sobrevivencia se utilizó la prueba no paramétrica de Kruskal-Wallis relativa a la ANOVA I, según las recomendaciones en Zar (1984).
Resultados
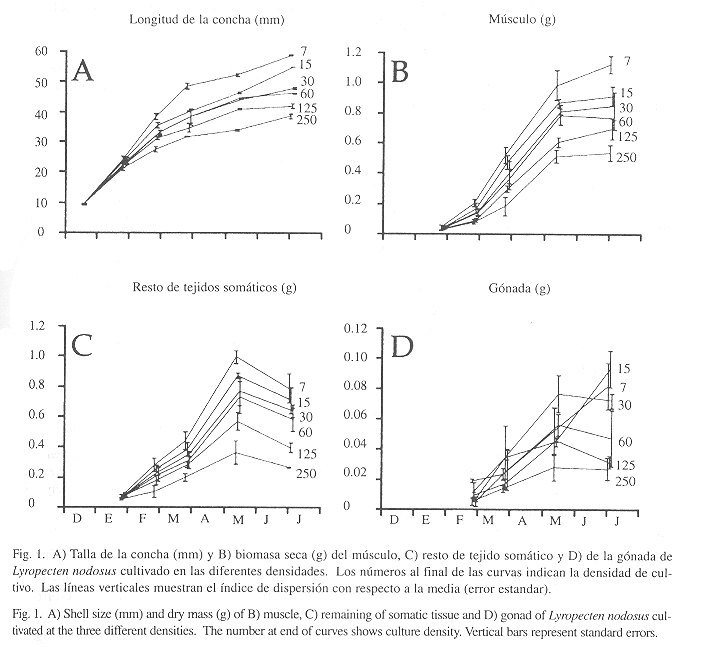
Crecimiento de la concha: En general, las curvas de las diferentes densidades mostraron tendencias similares, con altas tasas de crecimiento durante los dos y tres primeros meses de estudio, posteriormente la tasa disminuyó (Fig. 1A). Durante el período experimental la diferencia en las tallas alcanzadas entre las densidades aumentaron proporcionalmente. Para el primer mes de cultivo se establecieron diferencias significativas entre algunas densidades (ANOVA, P<0.001) y al final del estudio entre todas (ANOVA, P<0.001, Duncan P<0.05). La mayor longitud alcanzada fue de 55.80 ± 0.22 mm para la densidad de 7 ejemplares/cesta y la menor para los ejemplares mantenidos a 250 ejemplares/cesta (38.59± 0.72 mm).
Crecimiento de los tejidos somáticos: La tendencia del crecimiento en los tejidos somáticos (músculo y resto) fue similar en todas las densidades; siendo este patrón diferente al de la concha (Fig. 1B,C). El crecimiento fue exponencial hasta mediados del mes de mayo, posteriormente las tasas disminuyeron hasta el final del período experimental. En este último período las masas del resto de tejido somático decrecen y las del músculo se mantienen sin variación significativa, con excepción de la densidad de 7 ejemplares/cesta (ANOVA, Duncan P <0.05). Al final del estudio, la densidad de 250 ejemplares/cesta fue significativamente menor al de las demás densidades (ANOVA, Duncan P<0.05). En general, las densidades intermedias conformaron un grupo de igualdad significativa (Duncan P >0.05). Las masas alcanzadas en los tejidos somáticos de los individuos mantenidos a la densidad de 7 ejemplares/cesta duplicó a la obtenida en la densidad de 250 ejemplares/cesta.
El tejido gonadal comenzó a evidenciarse en febrero (Fig. 1D). En general, las densidades superiores a 15 ejemplares/cesta presentaron un crecimiento continuo hasta el mes de mayo, posteriormente las masas se mantienen o disminuyen al final del período. En contraste, los ejemplares mantenidos a las densidades de 7 y 15 ejemplares/cesta, los cuales a pesar de haber obtenido masas inferiores con respecto a los tratamientos de 30 y 60 ejemplares/cesta en mayo, presentaron siempre un incremento del tejido gonadal, que conllevó a alcanzar la mayor masa al final del período experimental (ANOVA, Duncan P <0.05).
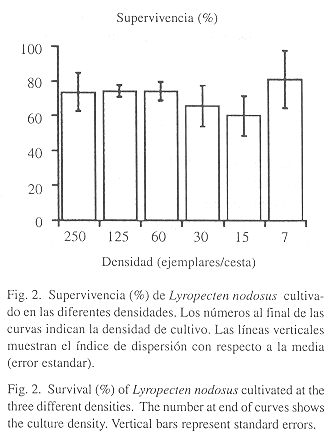
Supervivencia: La supervivencia acumulativa, al final del estudio fue mayor para los individuos mantenidos en el tratamiento de 7 ejemplares/cesta (Fig. 2); sin embargo, no se encontraron diferencias significativas entre los tratamientos (Kruskal-Wallis P>0.1).
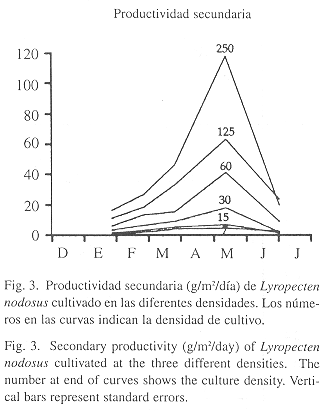
Productividad secundaria: La tendencia de la productividad secundaria fue directamente proporcional con la densidad (Fig. 3), mostrando un incremento progresivo en todos los tratamientos desde enero hasta mayo. La máxima productividad se obtuvo en este mes, en el tratamiento de 250 ejemplares/cesta (118 g/m2/día), la menor productividad se presentó en la densidad de 7 ejemplares/cesta (2.40 g/m2/día).
Parámetros ambientales: En líneas generales, la tendencia de la temperatura fue inversa a la biomasa fitoplanctónica, estimada por clorofila a (Fig. 4A,D), lo cual concuerda con los patrones establecidos para el Golfo de Cariaco, originado por la dirección y magnitud de los vientos prevalecientes en la región que causan surgencia (Okuda et al. 1978, Ferraz-Reyes 1987, Lodeiros y Himmelman 1994). No obstante, el seston total y orgánico presentaron tendencias no definidas en el transcurso del período experimental (Fig. 4 B,C). El rango de temperatura estuvo entre 21.5 y 25 °C y la clorofila a generalmente sobre 0.5 µg/L, alcanzando picos máximos entre 7 y 10 µg/L. El seston total presentó valores entre 5 y 20 mg/L, con máximos entre 50-70 mg/L y el orgánico valores sobre los 3 mg/L con máximos entre los 8 y 13 mg/L.
Discusión
Los resultados muestran que el crecimiento de los diferentes compartimientos del soma estudiado fue dependiente de la densidad de organismos de cultivo, ya que a partir del primer mes de cultivo se determinaron diferencias en crecimiento entre las densidades estudiadas. Ello sugiere que, el aumento de la densidad causó a un aumento de competencia intraespecífica por el alimento y espacio, el cual es un factor limitante en el cultivo de organismos filtradores (Okamora 1986, Vicent et al. 1989). No obstante, la densidad de organismos no afectó significativamente a la supervivencia, lo cual evidencia que, dentro del rango de densidades establecidas, la competencia bajo cultivo conllevó a un estrés que afectó al crecimiento pero no a la supervivencia.
Los patrones de crecimiento de las diferentes densidades en cada uno de los compartimientos del soma fueron similares, pero con magnitudes diferentes e inversamente proporcionales a la densidades de cultivo, ello sugiere que la influencia de factores (exógenos y endógenos) fueron periódicamente establecidos, afectando de manera negativa a los organismos mantenidos en las mayores densidades. Los factores ambientales que más influyen en el crecimiento son la disponibilidad de alimento y la temperatura (Griffiths y Griffiths 1987, Lodeiros y Himmelman 2000). En nuestro estudio el crecimiento de los tejidos somáticos fue exponencial hasta mediados del mes de mayo, coincidiendo con el período de mayor abundancia fitoplanctónica, posteriormente el crecimiento disminuye, principalmente en el resto de tejido somático coincidiendo con el período de menor abundancia fitoplanctónica. En este último período, el tejido gonadal muestra decrecimiento, probablemente por efectos de actividad reproductiva, exceptuando los organismos mantenidos a densidades de 7 y 15 ejemplares/cesta. De esta manera la reproducción, en períodos de baja disponibilidad de alimento, pudo inferir en el crecimiento somático, conllevando a una transferencia de energía del soma a la gónada. Estudios previos muestran un efecto negativo del desarrollo gonadal con respecto al crecimiento somático en períodos de baja abundancia fitoplantónica en L. nodosus (Lodeiros et al. 1998), Euvola ziczac (Boadas et al. 1997, Lodeiros 1996) y Pinna carnea (Narváez 1999) en el Golfo de Cariaco, así como en otros bivalvos en aguas templadas (Sundet y Vahl 1981, Epp et al. 1988).
El comportamiento de la masa gonadal de los pectínidos mantenidos a 7 y 15 ejemplares/cesta muestra que no existió actividades de desove, contradiciendo la posible actividad reproductiva señalada para los organismos de las demás densidades. El desove de los organismos mantenidos con mayores densidades pudo ser efectivo debido a condiciones evolutivas de los bivalvos, tal como lo establece Browne y Russel-Hunter (1978) y Toumi et al. (1983), quienes manifiestan que, cuando las fuentes de alimento se encuentran limitadas, el desarrollo gonadal es prioritario al crecimiento somático como una respuesta adaptativa a la conservación de las especies. El continuo crecimiento de la concha de los ejemplares mantenidos a densidades de 7 y 15 ejemplares/cesta y el menor decrecimiento del músculo en el período final del estudio, soportando una menor limitación de alimento para estos ejemplares, sostiene la hipótesis antes señalada. Por otra parte, estos resultados muestran que los efectos negativos provocados por la densidad de organismos mantenidos en cultivo, no fue notable, lo cual sugiere la posible utilización de 7 ejemplares/cesta para la continuación del cultivo en el engorde final.
La productividad secundaria fue directamente proporcional a las densidades de cultivo, ello podría conducir a establecer cultivos con las máximas densidades. Sin embargo, debido a que para esta especie la mejor estrategia para una mayor rentabilidad en la producción, es obtener organismos de talla grande que alcancen músculos de gran peso (>20 g) con los mejores precios unitarios en el mercado (Dore 1991), el objetivo del cultivo intermedio para L. nodosus es obtener organismos de tallas adecuadas (50 mm) para el engorde final. Lo anterior permite sugerir el cultivo de L. nodosus en tratamientos de baja densidad. No obstante, en el cultivo intermedio, esta estrategia conllevaría a altos costos de producción que disminuyen la rentabilidad económica del cultivo. En este sentido, se recomienda una estrategia de combinación de desdoble de la densidad de cultivo, siendo lo más recomendable iniciar con 250 ejemplares/cesta, una vez transcurrido el primer mes, se realizaría un desdoble a 125 ejemplares/cesta, el cual se mantendría durante dos meses, al cuarto mes se realizaría otro desdoble a la densidad 60 ejemplares/cesta y finalmente en el siguiente mes se reduciría a 30 ejemplares/cesta. Con esta estrategia se obtendría una talla aproximada de 50 mm adecuada para comenzar el cultivo de engorde final. En esta etapa, la densidad de 15 o 7 ejemplares/cesta sería la recomendada. Es importante indicar que la estrategia de cultivo se ha establecido en un período de surgencia y la misma puede ser dependiente de las condiciones ambientales relativas al período de cultivo.
Agradecimientos
La investigación fue financiada por el Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICIT) Proyecto S1-2456 y al Consejo de Investigación de la Universidad de Oriente. Se le agradece la valiosa colaboración técnica a M. Nuñez, A. Sotillet, A. Rojas, E. González y al personal obrero de la Estación Hidrobiológica de Turpialito del Instituto Oceanográfico de Venezuela de la Universidad de Oriente.
Resumen
El efecto de la densidad sobre el crecimiento y la supervivencia de juveniles del pectínido Lyropecten nodosus se estudió en condiciones de cultivo intermedio en el Golfo de Cariaco, Estado Sucre, Venezuela (diciembre 93 - julio 94). Se evaluaron seis densidades de siembra (7, 15, 30, 60, 125 y 250 ejemplares/cesta) con una longitud y una masa seca inicial de 9.4 ± 0.18 mm y 0.007 ± 0.0026 g, respectivamente. Los juveniles fueron colocados en cestas japonesas "pearl nets" y posteriormente suspendidas en un "long line" a 8 m de profundidad. Los patrones de crecimiento en cada uno de los compartimientos del soma fueron similares, pero con magnitudes diferentes e inversamente proporcionales a la densidades de cultivo; sin embargo, la supervivencia no fue afectada significativamente, sugiriendo que la competencia intraespecífica de los organismos en cultivo provocó un estrés notable, aunque insuficiente para causar mortalidad. Los resultados y estrategias proyectadas para el cultivo intermedio de L. nodosus sugieren un procedimiento de disminución de la densidad de organismos a través de desdobles sucesivos en un período de cultivo de seis meses obteniendo tallas cercanas a los 50 mm para comenzar el cultivo de engorde.
Referencias
Altuve, C. A. 1986. Variación genética en una población del pectínido Lyropecten nodosus (Linné, 1758) de la Isla de Cubagua. Trabajo de grado, Licenciatura en Biología, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Anónimo. 1998a. Aquaculture production statistics. 1987-1996. FAO Fisheries circular No. 815, Rev. 10 FAO, Roma, Italia. 197 p. [ Links ]
Anónimo. 1998b. Fisheries statistics. Capture production. Vol 82, FAO Fisheries series. No. 50. FAO, Roma, Italia. 678 p. [ Links ]
Boadas, M. A.; O. Nusetti, F. Mundarain, C. Lodeiros & HE Guderley. 1997. Seasonal variation in the properties of muscle mitochondria from the tropical scallop, Euvola (Pecten) ziczac. Mar Biol. 128: 247-255. [ Links ]
Browne, R. A. & W.D. Russel-Hunter. 1978. Reproduction effort in molluscs. Oecologia (Berlin) 37: 23-27. [ Links ]
Coronado, M.C. 1987. Variación genética y estudios de la meiosis en la vieira Lyropecten nodosus (L. 1758). Tesis de M. Sc. Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Côté, J., J.H. Himmelman, M.R. Claereboudt & J. Bonardelli. 1993. Influence of density and depth on the growth of juvenile giant scallops (Placopecten magellanicus, Gmelin, 1791) in suspended culture in the Baie des Chaleurs. Can. J. Fish. Aquat. Sci. 50: 1857-1869. [ Links ]
Dore, I. 1991. Shellfish. Van Nostrand Reinhold, Nueva York. 239p. [ Links ]
Epp, J., M. Bricelj & R. Malouf. 1988. Seasonal partitioning and utilization of energy reserves in two ages clases of the bay scallop Argopecten irradians (Lamarck). J. Exp. Mar. Biol. Ecol. 121: 112-136. [ Links ]
Ferraz-Reyes, E. 1987. Productividad primaria del Golfo de Cariaco, Venezuela. Bol. Inst. Oceanogr. Univ. Oriente 26 (1-2): 91-110. [ Links ]
Freites, L., B. Vera, C. Lodeiros & A. Vélez. 1995. Efecto de la densidad sobre el crecimiento y la producción secundaria de Euvola (Pecten) ziczac bajo condiciones de cultivo suspendido. Ciencias Marinas 21 (4): 1-12. [ Links ]
Griffiths, C.L. & J.S. Griffiths. 1987. Bivalvia, p. 1-88. In Pandian, J.H., Vernberg, F.J. (eds.). Animals Energetics. Vol. 2. Academy, Nueva York. [ Links ]
Lodeiros, C. J. 1996. Influence des facteurs environnementaux sur la croissance du pétoncle tropical Euvola (Pecten) ziczac (L. 1758) cultivé en suspension au Golfo de Cariaco, Vénézuela. Tesis de Ph.D., Departamento de Biología, Universidad Laval. Québec, Canada. 83 p. [ Links ]
Lodeiros, C. J. & J. H. Himmelman. 2000. Identification of factors affecting growth and survival of the tropical scallop Euvola (Pecten) ziczac in the Golfo de Cariaco, Venezuela, Aquaculture 182: 91-114. [ Links ]
Lodeiros, C.J.& J. H. Himmelman. 1994. Relations among environmental conditions and growth in the tropical scallop Euvola (Pecten) ziczac (L.) in suspended culture in the Golfo de Cariaco, Venezuela. Aquaculture 119: 345-358. [ Links ]
Lodeiros, C., B. Marín & A. Prieto. 1999. Catálogo de moluscos de las costas nororientales de Venezuela. Clase Bivalvia. Ediciones APUDONS, 103 p., 9 Láminas. [ Links ]
Lodeiros, C., J. Rengel, L Freites, F. Morales & J. H. Himmelman. 1998. Growth and survival of the tropical scallop Lyropecten (Nodipecten) nodosus, maintained in suspended culture at three depths. Aquaculture 165: 41-50. [ Links ]
Marín, B. 1984. Desove y desarrollo larval de la vieira Lyropecten nodosus Trabajo de grado, Licenciatura en Biología, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Mendoza, I. 1999. Alternativas para el cultivo final del pectínidos Lyropecten (Nodipecten) nodosus (Linné, 1758), en el Golfo de Cariaco, Estado Sucre, Venezuela. Trabajo de grado, Licenciatura en Biología, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Narváez, N. 1999. Crecimiento de Pinna carnea L., 1758 (Mollusca: Bivalvia) en cultivo suspendido en el Golfo de Cariaco, Estado Sucre, Venezuela. Trabajo de grado, Licenciatura en Biología, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Okuda, T., J. Benitez-Alvarez, J. Bonilla & G. Cedeño. 1978. Caracteristicas hidrográficas del Golfo de Cariaco, Venezuela. Bol. Inst. Oceanogr. Univ. Oriente 17: 69-88 [ Links ]
Okamora, Y. 1986. Review of scallop culture technique study. Aquacult. Cent. Amori Pref. 19: 65-68. [ Links ]
Rhodes, E. & J. Widman 1991. Some aspects on the controlled production of the bay scallop Argopecten irradians. World Maricult. Soc. (11): 235-246. [ Links ]
Strickland, J. & T. Parsons 1972. A practical handbook of water analysis. Bull. Fish. Rev. Gd. Canadá. 2da. Edition, 16: 167- 315. [ Links ]
Sundet, J.H. & Vahl, O. 1981. Seasonal changes in dry weight and biochemical composition of sexually mature and immature Iceland scallops, Chamys islandica.. J. Mar, Biol. Ass.U.K. 61: 1001-1010. [ Links ]
Toumi, J., Hakala T. & Haukioja E. 1983. Alternative concepts of reproductive effort, costs of reproduction and selection in life history evolution. Am. Zool. 23: 25-34. [ Links ]
Vélez, A. & C. Lodeiros 1990. El cultivo de moluscos en Venezuela, pp. 345-369. In Hernandez A. (ed.). Cultivo de moluscos en América Latina. Red Regional de Entidades y Centros de Acuicultura de América Latina. CIID-Canadá. [ Links ]
Vélez, A. & L. Freites. 1993. Cultivo de semillas de la vieira Pecten ziczac bajo condiciones ambientales controladas (Hatchery). IV COLACMAR. Serie ocasional 2: 311-317. [ Links ]
Vélez, A., F. Sotillo & J. Pérez 1987. Variación mensual de la composición química de las vieiras Pecten ziczac, Lyropecten nodosus y su relación con los períodos de desove. Bol. Inst. Oceanogr. Univ. Oriente 25 (1-2): 155-162. [ Links ]
Vicent, B., D. Joly & C. Brassard. 1989. Effects de la densité sour la crossance du bivalve Macoma balthica (L) en zone intertidale. J. Exp. Mar. Biol. Ecol. 126: 145-162. [ Links ]
Zar, J.H. 1984. Biostatistical Analysis. 2nd Edition. Pentice-Hall, Nueva Jersey. [ Links ]
1Lab. Acuicultura, Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná 6101, Venezuela. Fax: ø93-315902/512276 Correo electrónico: clodeiro@ci.udo.edu.ve , clodeiro@sucre.udo.edu.ve

